











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Blueberries (Vaccinium spp.) have stood out among the small fruits cultivated in Brazil due to their flavor, economic value, and nutraceutical properties (Peña et al., 2012). Cantuarias-Avilés et al. (2014) and Radünz et al. (2016) have shown their great nutraceutical and nutritional values, besides the plant's easy adaptation to small areas of cultivation. These characteristics make blueberries an attractive species for diversifying the productive capacity of families (Radünz et al., 2014).
Rabbiteye (Vaccinium ashei Reade) is among the world's most commercially cultivated species (Strik, 2007; Cantuarias-Aviles et al., 2014), mainly because this group has plants with high yields, vigor, longevity, resistance to heat and drought, tolerance to fungus-related diseases, and low chilling requirements (Ehlenfeldt, 2007). In southern Brazil, a low number of chill hours is very common, a fact that increases the importance of this group for fruit farming in temperate zones.
Genetic and climatic factors may influence the productive characteristics of blueberry plants (Antunes et al., 2008) as well as the quality of the harvested fruits (Rodrigues et al., 2011; Gündüz et al., 2015). For temperate regions of Brazil, there is a need for studies regarding blueberry species and management practices to better adapt these plants to the local edaphoclimatic conditions and, therefore, to obtain higher productivity and fruit quality (Fachinello et al., 2011).
Originally, all the buds in blueberry plants are vegetative (Pescie & López, 2007), and in order for the flower buds to become differentiated, the presence of low temperatures is necessary to meet the required chilling hours of the particular cultivar (Coletti et al., 2011). The number of vegetative buds differing from flower buds might vary depending on the cultivar, day length, temperature (Spann et al., 2003; Pescie & López, 2007; Williamson et al., 2015), and the phytosanitary status of the plants (Williamson et al., 2015). Therefore, there might be a variation in the number of flower buds, flowers, and fruits for the same cultivar, depending on the climatic conditions of the place where the plants are grown.
Thus, in order to obtain increased knowledge on the bioclimatology of blueberry plants cultivated under the climatic conditions of southern Brazil, especially in relation to their fruiting habit and their production linked to environmental conditions, it is important to characterize the crops in this region. The characterization will allow the proper use of management practices and obtain greater yields and better fruit quality. Since studies regarding the characterization of blueberries grown under climatic conditions of Brazil are scarce, this research focused on characterizing the blueberry (Vaccinium spp.) cv. Bluegem group Rabbiteye (Vaccinium ashei Reade) grown under southern Brazilian climatic conditions.
Materials and methods
The study was carried out during two harvest seasons, 2012/2013 and 2013/2014, in a commercial orchard located in Morro Redondo, RS, in Brazil (31°32' S, 52°34' W, 150 m a.s.l.). The average temperatures during the experiment were 17.1°C and 18.2°C for the 2012/13 and 2013/14 harvests, respectively. Climatological normal data for the period 1971 to 2000 demonstrate that the region has a mean annual rainfall of 1366.9 mm, mean annual temperature of 17.8°C and a mean temperature of the hottest month of 23.2°C in January (Embrapa, 2016).
Eight-year-old plants of the cv. Bluegem belonging to the Rabbiteye (Vaccinium ashei Reade) group and in full production were chosen. For this region of the country under the conditions of the experiment, pruning was carried out in mid-July. This cultivar showed vegetative growth for approximately 40 d during the months of July and August. The period of floral initiation and formation of flower buds occurred for approximately 15 d during the months of August and September. The period that comprises the formation and maturation of fruits took place for approximately 140 d during the months of September, October, November, December, and January. The fruit harvest started in January.
A totally randomized experimental design was used and conducted under two factorial arrangements. A 2x2 factorial arrangement (shoot length x harvest season) was used to check the distribution of vegetative and flower buds on the shoot. A 2x3x2 factorial arrangement (shoot length x bud position on the shoot x harvest season) was used to check the number of floral primordia, open flowers, and developed fruits in the plants.
Four groups of plants were randomized for each variable. Each group consisted of four plants. Two central plants in each group were selected and a total of ten long shoots (31 to 50 cm) and ten short shoots (15 to 30 cm) were randomly selected for each harvest season. The shoots evaluated corresponded to the secondary shoots (secondary branches originated on the main stem). Shoots were grouped into long and short to obtain more accurate results, since shoots of different sizes in the plant may have a different number of buds. For the selected shoots, counts of flower and vegetative buds, the position of these buds on the shoots, the base and top diameter of the shoots, and their lengths were assessed. Weekly observations in marked and defined buds were carried out to determine the number of open flowers during the flowering period and the number of developed fruits until the end of the harvest season (Tab. 1).
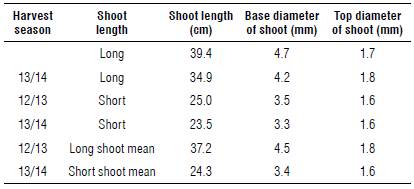
TABLE 1 Morphological characterization of the shoots of blueberry, cv. Bluegem, harvest seasons 2012/2013 and 2013/2014.
In order to evaluate the number of primordia, shoots were collected from plants on five different dates between April and August for each harvest season, covering the period of leaf senescence until the opening of flowers. In each of the assessment dates, a total of ten short and ten long shoots were collected from each group of bushes and were taken to the Fruit Laboratory of the Federal University of Pelotas for the dissection of apical, medial and basal buds, and the counting of floral primordia. At the end of each counting, a mean number of floral primordia was obtained for the apical, medial and basal portions of the shoots.
The shoot diameter was measured with a digital caliper. Measurements were performed twice at the base and at the top of each shoot and the values were averaged. The shoot length was measured with a measuring tape, from the base to the top of the shoot.
The total number of flower and vegetative buds in long and short shoots were counted and weighted in relation to the number of shoots in which the buds were present, for the different levels of shoots. The levels were characterized by the number of times that they showed flower and vegetative buds intercalated, as shown in Figure 1. This method of weighting was performed to avoid overrating the total number of buds. Thus, the flower and vegetative buds that occupied the first level, i.e., the apical part of the shoot, were multiplied by 1, since they could be found in all evaluated shoots. The flower and vegetative buds from the second level were multiplied by the respective number of shoots in which they were found; for example, when they were found in six shoots, they were multiplied by 0.6. The same was done for the other levels (Radünz et al, 2018).
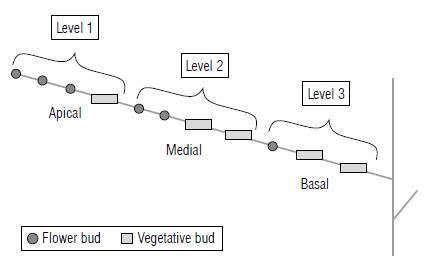
FIGURE 1 Representation of the levels used to characterize the fruiting habit on the secondary shoots of blueberry cv. Bluegem.
Data on the total number of flower and vegetative buds as a function of the shoot length and the harvest season, and the number of floral primordia, flowers, and fruits as a function of the shoot length, bud position, and harvest season were subjected to an ANOVA and, when statistically different, the Tukey's HSD test (P≤0.05) was used to compare the means of the treatments.
Results
The results of the ANOVA (P≤0.05) for the number of flower and vegetative buds, floral primordia, number of flowers, and number of fruits are shown in Table 2.
TABLE 2 Results of the analysis of variance (ANOVA) for the number of flower (FB) and vegetative (VB) buds, floral primordia (FP), number of flowers (NFW) and number of fruits (NFT) of the blueberry cv. Bluegem.
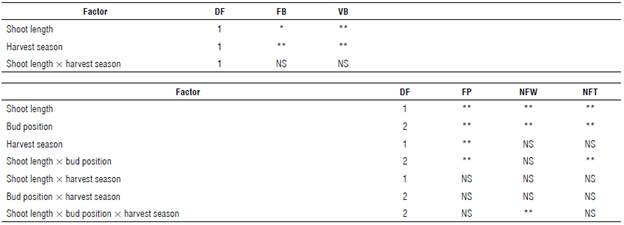
*, **, *** significant at P≤0.05, P≤0.01 and P≤0.001, respectively; NS - not significant. DF - Degrees of freedom.
The interaction between the factors harvest season and shoot length did not significantly affect the total number of flower and vegetative buds. However, the single effect of these variables influenced the total amount of flower and vegetative buds in the plants (Tab. 2). A greater number of flower and vegetative buds (Fig. 2) was observed for the 2012/13 and 2013/14 harvest seasons, respectively. When analyzing the number of flower and vegetative buds as linked to the shoot length, a statistical difference was found between short and long shoots. Long shoots showed a greater number of flower and vegetative buds, having on average 53% more buds than short shoots (Fig. 2; Tab. 3). For the proportion between the number of vegetative and flower buds, 1.43, and 1.41 vegetative buds were observed for each flower bud in long and short shoots, respectively (Tab. 3).
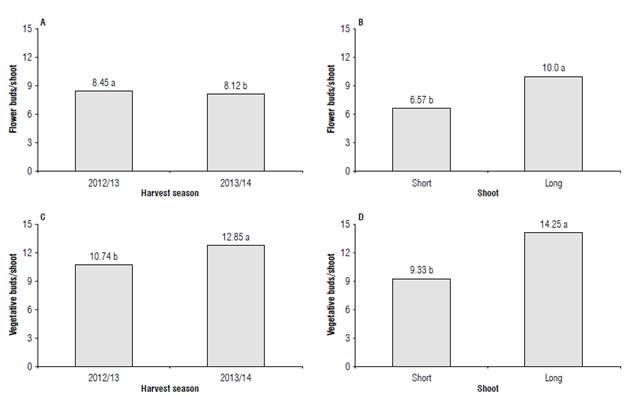
FIGURE 2 Total mean number of A and B) flower and C and D) vegetative buds as a function of the harvest season and secondary shoot length (short shoot: 15 to 30 cm and long shoot: 31 to 50 cm) for blueberry cv. Bluegem. Means followed by the same letter In the figure are not significantly different according to the Tukey's HSD test (P≤0.05).
TABLE 3 Bud distribution in levels and total buds (TB) for blueberry cv. Bluegem as a function to the harvest season (H) and shoot length (S).
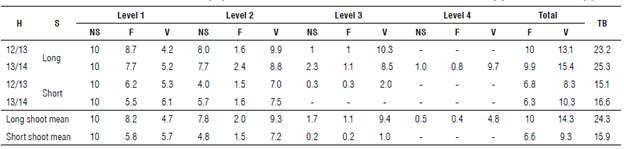
Mean of three plants (30 shoots). NS - Number of shoots in which levels are present; F- number of flower buds in the level; V- number of vegetative buds in the level.
Analyzing the levels in which flower buds were intercalated with vegetative ones for every ten evaluated shoots on average, 78% and 48% showed intercalated buds in long and short shoots, respectively. In the third level, the intercalation was of 17% in long shoots and 2% in short shoots, while for the fourth level an intercalation of 5% was observed only in long shoots (Tab. 3). The highest number of flower buds was found in long and short shoots of the first level (Tab. 3), which, on average, showed 8.2 and 5.8 flower buds, accounting for 82% and 88% of all the buds found on these shoots, respectively.
Regarding the total distribution of buds on the shoots as a function of shoot length (Tab. 3), we verified that for every 1.53 cm (0.65 buds cm-1) there was a bud in both short- and long shoots. The similarity in the values indicated that the total number of buds per shoot was directly associated with the shoot length, and so was the proportion between flower and vegetative buds.
A distinct statistical tendency was identified for the number of primordia, flowers, and fruits. An interaction between the shoot length and the bud position was observed for the number of primordia and fruit, as well as the single effect of the harvest season on the number of primordia. The number of flowers was affected by the three-way interaction shoot length x bud position x harvest season.
When analyzing the interaction between the position of the bud (apical, medial, and basal) and the shoot length (short and long) for the number of floral primordia (Tab. 2), we found differences between the shoot lengths only in relation to the medial and basal positions, since long shoots showed the greatest number of floral primordia. When analyzing the interaction of the short shoot as a function of the bud position, we observed that buds were found in a greater number in the apical portion, followed by the basal and the medial ones. This behavior was different from that observed in the long shoot, in which the medial portion stood out (Tab. 4). We observed no significant differences for the number of primordia as a function of the harvest season (mean 7.98), showing that the results referring to the number of floral primordia were stable and were not influenced by the edaphoclimatic characteristics of the region.
TABLE 4 Number of floral primordia of blueberry cv. Bluegem as a function of the shoot length and bud position.
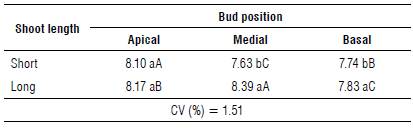
Means followed by the same letters (lowercase in the column and uppercase in the same line) are not significantly different according to theTukey's HSD test (P≤0.05). CV - coefficient of variation.
The number of flowers was significantly influenced by all the factors involved in the research (Tab. 2). However, it did not differ for the apical bud position for the two harvest seasons assessed. We observed a similar behavior for the medial and basal bud positions in the two harvest seasons, with a greater number of flowers in long shoots (Tab. 5). When analyzing the interaction for short shoots, only the position of the buds affected the number of flowers, as the apical position was on average 17.2% greater than the basal one (Tab. 5). The behavior for the long shoot was similar, with a difference between the apical and basal bud position, although we found no differences between the apical and medial positions, and the medial and basal positions, except for the medial position in the 2013/14 harvest season (Tab. 5).
TABLE 5 Number of blueberry cv. Bluegem flowers as a function of the interaction between the bud position and the harvest season on the shoot length.
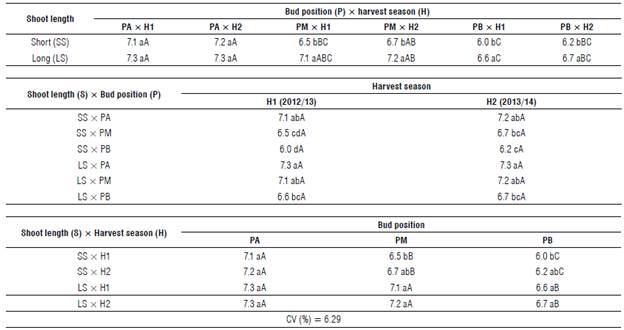
Means followed by the same letters (lowercase in the column and uppercase in the same line) are not significantly different according to the Tukey's HSD test (P≤0.05). PA - apical; PM - medial; PB - basal. H1 - 2012/13; H2 - 2013/14. CV- coefficient of variation.
The relationship between the shoot length (S) and the bud position (P) as a function of the harvest season can be observed in Table 5. No statistical difference was observed between the harvest seasons; however, the interaction between the bud position and the shoot length had a similar behavior. In both cases, we found the highest number of flowers on the long shoot - apical bud position (mean 7.30), which did not differ from the long shoot - medial bud position (mean 7.15) and the short shoot - apical bud position (mean 7.15). Nevertheless, the lowest number of buds was found on the short shoot - basal bud position, which did not differ statistically from the short shoot - medial bud position for both harvest seasons, and from the long shoot - basal bud position in the 2013/14 harvest (Tab. 5).
The number of flowers for the interaction between shoot length (S) and harvest season (H) as a function of the bud position (P) can be seen in Table 5. The interaction between the harvest season and the shoot length did not influence the number of flowers in the apical bud position of the shoot. However, this interaction had a similar behavior in the medial and basal positions, considering that in both cases (medial and basal positions) the greatest number of flowers was observed in long shoots, regardless of the harvest season. This number of flowers was similar to that observed in short shoots from the harvest 2013/14 (Tab. 5). When analyzing the number of flowers as a function of the bud position on the shoot, in all cases the lowest number of flowers was found in the basal position whereas the highest number of flowers was found in the apical one. However, we observed no differences between the apical and the medial positions for long shoots (Tab. 5).
Regarding the number of fruits for the interaction between the position of the bud and the shoot length (Tab. 2), the apical bud position was not influenced by the shoot length and produced on average 7.10 fruits. For the medial and basal position of buds on the shoots, the behavior was similar. The short shoot showed a greater number of fruits, on average 12.3% and 11.0% greater than the long shoot, respectively (Tab. 6).
TABLE 6 Number of blueberry cv. Bluegem fruits as a function of the bud length in the bud position.
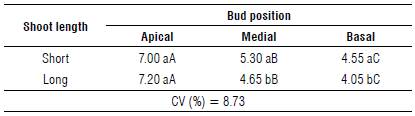
Means followed by the same letters (lowercase in the column and uppercase in the same line) are not significantly different according to the Tukey's HSD test (P≤0.05). CV - coefficient of variation.
When analyzing the same shoot length, we saw that the behavior was the same regardless of the length; in both cases, the values of the apical position of the buds were higher than those of the medial position, which in turn were higher than those of the basal position. We observed for the short shoot that the medial and basal bud positions showed 24.3% and 35.0% fewer fruits than the apical one, respectively. For the long shoot, the apical position of the buds showed 35.4% and 43.8% more fruits than the medial and basal positions, respectively (Tab. 6).
Discussion
The observed behavior for the number of buds is of extreme importance for fruit production and quality, as vegetative buds are those that will support the production of pho-toassimilates (Radünz et al., 2016). In Georgia, USA, the maximum number of flower buds formed in each shoot piece of 20 cm is 14 for the cultivar Premier belonging to the Rabbiteye group (Ojiambo et al., 2006).
In blueberry bushes, flower buds are found in the upper part of the shoot while the vegetative buds are found in the basal part (Karimi et al., 2017). The results from this work can be attributed to the climatic conditions found in the farming region, especially those in southern Brazil, where mild winters are frequent (Fachinello et al., 2011). Originally, all the buds of blueberry bushes are vegetative and, depending on the day length and temperature, they can turn into flower buds (Song & Walworth, 2018). The existence of intercalated buds in the shoots might be related to the adjustment of the cultivar to the region since it is an exotic species for the climatic conditions of Brazil.
When analyzing the total number of buds per shoot, similarities were seen, leading to the belief that the total number of buds per shoot is directly associated with the shoot length and so is the proportion between flower and vegetative buds. So, it is possible to estimate the number of buds for a given shoot length. There might be a correlation between the shoot thickness and the fruit size, as it represents a natural evolution of the species where larger and thicker shoots may provide better conditions to support higher yields (Braha & Zajmi, 2015).
The apical portion of the shoots showed a greater number of flowers when compared to the basal portions of the shoots. These results are associated with the greater capacity of solar radiation interception in the external portions of the plant canopy, considering that the architecture of the plant is a factor that influences its functions and characteristics (Retamal-Salgado et al., 2017). The vegetative morphology of the plant is correlated with its reproductive characteristics. Because of its evolutionary traits, the development of flowers happens in places where they can be better found by pollinators (Stournaras & Schaefer, 2017). Plant morphology also influences the number of flowers as basal flowers are located in the canopy of the plants and play a role in the attractiveness and arrival of pollinators (Stournaras & Schaefer, 2017). Floral induction takes place through environmental stimuli such as temperature and photoperiod that, in turn, might influence the number of differentiated primordia (Wilkie et al., 2008).
Conclusion
The harvest season and the shoot length influenced the total number of buds and the number of flower and vegetative buds, since a greater number of vegetative buds were observed. The bud position on the shoot influenced the number of floral primordia but was not influenced by the harvest season. The number of flowers and fruits, in general, was greater in apical buds and in long shoots, compared to the other positions and shoot lengths.
These results can be used by farmers as a decision-making tool to define the moments in which cultural practices are carried out. However, further studies are necessary for the detailed evaluation of floral differentiation and the reasons why it occurs in an intercalated form. Additionally, research that addresses the formation of flowers and fruit set and their relationship with the climatic characteristics of the region is also suggested here.

