











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
Las mariposas diurnas (Lepidoptera; Rhophalocera) forman el grupo de los insectos vistosos (Salazar, 1995) por la belleza de sus formas y colores. La elegancia de sus vuelos y su comportamiento dócil son requeridos por coleccionistas y artesanos, se utilizan como atractivo turístico y son empleados para la sensibilización ambiental, por lo cual tienen un gran potencial para la educación ambiental y los bionegocios; convirtiéndolas en una opción para mejorar los ingresos complementarios de las familias rurales amazónicas, ya que pueden ser criados en cautiverio (Constantino, 1996; Vásquez, Zárate, Pinedo, et al. (2017); Ruiz et al., 2015).
Estos grupos de insectos representan importancia ecológica, su presencia en el trópico es un indicador de endemismo debido a las formaciones vegetales que los albergan (Salazar, 1995), su taxonomía está bastante avanzada en comparación a otros taxones de insectos, y por su interrelación específica con las plantas entre las larvas herbívoras y los adultos polinizadores han sido sugeridos como indicadores de diversidad de plantas (Brown, 1991; Beccaloni y Gaston, 1994). Así mismo, influencian en la interacción de la dinámica poblacional de la vegetación, la relación entre grupos discretos de lepidópteros con grupos discretos de plantas ayuda a comprender que las mariposas están coevolucionando (Rausher y Feeny, 1980; Jennersten, 1988; Kremen, 1992; Vásquez, Zárate, Huiñapi, et al., 2017).
Las mariposas son altamente sensibles a los cambios y disturbios ambientales; los inventarios de sus comunidades constituyen una valiosa herramienta para valorar el estado de conservación o alteración del medio natural (Kremen et al., 1993; Andrade-C, 1998), pero en el departamento de Loreto (Perú), solo existen tres estudios de lepidópteros; uno sobre las mariposas del alto Río Napo con un registro de 673 especies (Lamas, Robbins y Harvey, 1996), el segundo dentro de la Reserva Nacional Allpahuayo Mishana, con un registro de 518 especies (Campos y Ramírez, 2005) y el tercero en los bosques de tierra firme de la comunidad de San Rafael con un registro de 179 especies (Novoa y Montero, 2010). Sin embargo, no se conoce si han ocurrido cambios en la composición y abundancia de las especies de mariposas después de una década del primer estudio realizados por Campos y Ramírez (2005) en los bosques sobre arena blanca.
Los objetivos del presente estudio fueron evaluar la diversidad y composición de las mariposas (Lepidoptera: Morphinae y Satyrinae) en los bosques sobre arena blanca, denominados localmente como varillales en la Reserva Nacional Allpahuayo Mishana (Perú) con fines de educación ambiental y bionegocios.
Material y métodos
Área de estudio
El trabajo fue realizado desde enero hasta diciembre del 2015 en los bosques sobre arena blanca denominados varillales, en el Centro de Investigaciones Allpahuayo, ubicado aproximadamente en el km 26,5 de la carretera Iquitos - Nauta, distrito de San Juan Bautista, provincia de Maynas, región Loreto de la República del Perú (3°38’01.99”S; 73°25’08.86”W). El área de estudio corresponde a Bosques sobre arena blanca, clasificados en varillales altos y varillales bajos (García el al., 2003). Los varillales altos alcanzan un porte de 10 a 15 metros, mientras que en los varillales bajos fluctúan entre 8 a 10 metros. Las coordenadas geográficas de los sitios de muestreos están indicadas en la Tabla 1 y se grafican en la Figura 1.
Tabla 1 Ubicación de los sitios de muestreos de las mariposas de la Reserva Nacional Allpahuayo Mishana (RNAM), Loreto, Perú.
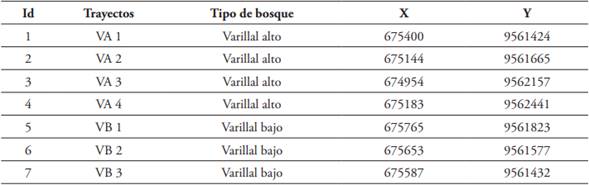
Fuente: elaboración propia.

Muestreo de mariposas
Para el muestreo se colocaron cebos en rodajas del plátano maduro (Musa paradisiaca L.) macerado en jugo de caña (Saccharum officinarum L.) como atrayente a lo largo de siete transectos de 500 m lineales dentro de los bosques de varillal bajo y varillal alto según el método de Campos y Ramírez (2005). Los cebos fueron colocados un día antes de la recolecta, cada punto fue codificado como VB1, VB2, VB3 en el varillal bajo y VA1, VA2, VA3 y VA4 en el varillal alto. Los especímenes fueron atrapados con la red entomológica durante la alimentación sobre los cebos, el muestreo se realizó durante una semana por cada mes en el transcurso de un año de 7:00 y 12:00 h y por la tarde desde las 14:00 hasta 16:30 h. Las muestras capturadas fueron colocadas en sobres entomológicos para su traslado al laboratorio del Centro de Investigaciones del Instituto de Investigaciones de la Amazonía Peruana (IIAP) para su determinación taxonómica, posteriormente fueron depositados en envases de plástico rectangulares con naftalina para su conservación.
Diversidad y composición de mariposas
La diversidad de mariposas se determinó en función a la riqueza de las especies y del índice de Shannon-Wienner (Moreno, 2001) por que se utiliza frecuentemente en ecología de comunidades y permite realizar comparaciones con otros trabajos; además permite conocer la homogeneidad en la cantidad de individuos por especies. Para la composición se procedió a identificar las especies a través de comparaciones de muestras del Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos (MHNUNMSM) y de claves taxonómicas y referencia de mariposas de la zona de estudio (Lamas, Campos y Ramírez, 2003).
Procedimiento de los datos
Se determinó la diversidad alfa de las mariposas en los transectos, a partir de la riqueza de especies y del índice de Shannon-Wienner (Moreno, 2001). Los valores de diversidad alfa de los transectos fueron comparadas para determinar su diferencia con la prueba de T de Student para los datos paramétricos de cantidad de especies y la prueba de Mann Whitney para los datos no paramétricos del índice de diversidad de Shannon-Wienner, estas pruebas se realizaron en el programa SigmaPlot. El cálculo de la diversidad de Shannon-Wienner se realizó con el programa DivEs v.3.0.
Resultados
Diversidad de mariposas
La diversidad de mariposas según el índice de diversidad de Shannon-Wienner, es de baja a media en las dos subfamilias estudiadas para ambos tipos de bosques, la subfamilia Morphinae fue menos diversa que la sub-familia Satyrinae. Para Satyrinae, reportamos un valor de 1,34 en bosques de varillal alto y 1,36 para varillal bajo; mientras para el caso de Morphinae el valor fue de 0,86 en varillal alto y 1,01 en varillal bajo.
Composición y abundancia de las mariposas
En los siete transectos, se colectaron 2662 individuos incluidos en 16 géneros y 38 especies. 2112 individuos correspondieron a 24 especies de Satyrinae y 550 individuos a 14 especies de Morphinae. La especie más abundante fue Pierella lena con 148 individuos (Figura 3D) y Cissia penelope la menos abundante con un total de 34 individuos, ambas especies de la subfamilia Satyrinae. En relación a la cantidad de individuos por tipos de bosque variaron de 0 a 35 para la subfamilia de Satyrinae y sus especies estuvieron presentes en todos los sitios de ambos tipos de bosques (Figura 2 y Tabla 2), para el caso de Morphinae, la especie más abundante fue Morpho helenor con un total de 91 individuos encontrados (Figura 3B) y la menos abundante fue Morpho telemachus con un solo individuo (Figura 3C). Respecto a la cantidad de individuos por sitios de muestreo, estos variaron de 0 a 29 para la subfamilia de Morphinae, pero no todas sus especies estuvieron presentes en los sitios de muestreos (Figura 2 y Tabla 3).

Fuente: elaboración propia.
Figura 2 Abundancia de las especies de Morphinae y Satyrinae encontradas en los Varillales bajo y alto de la Reserva Nacional Allpahuayo Mishana.
Tabla 2 Composición y abundancia de especies de Satyrinae encontradas en la Reserva Nacional Allpahuayo Mishana (RNAM).
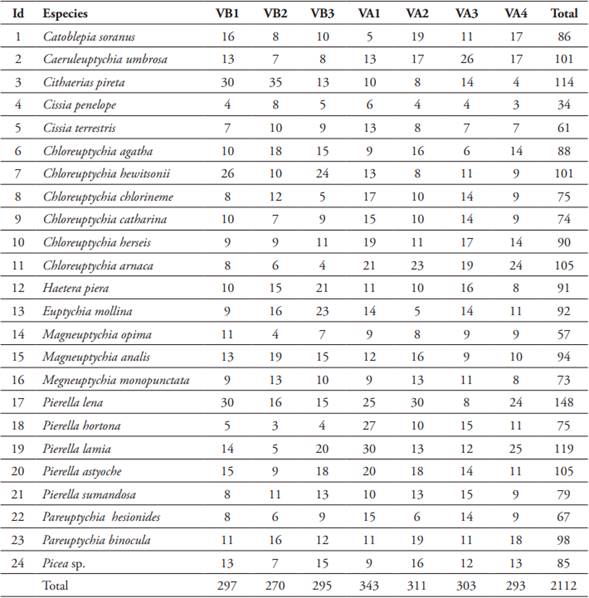
Nota: VB1: varillal bajo sitio 1, VB2: varillal bajo sitio 2, VB3: varillal bajo sitio 3, VA1: varillal alto sitio 1, VA2: varillal alto sitio 2, VA3: varillal alto sitio 3, VA4: varillal alto sitio 4.
Fuente: elaboración propia.
Tabla 3 Composición y abundancia de especies de Morphinae encontradas en la Reserva Nacional Allpahuayo Mishana (RNAM).
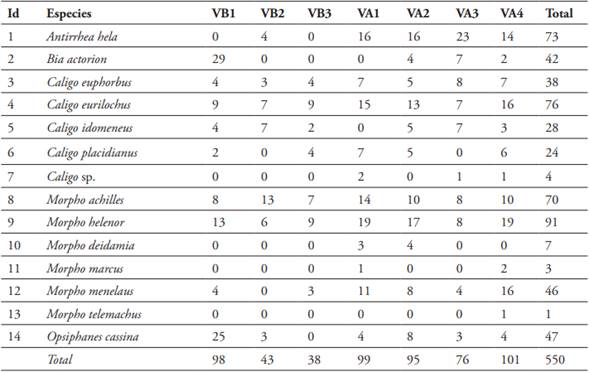
Nota: VB1: varillal bajo sitio 1, VB2: varillal bajo sitio 2, VB3: varillal bajo sitio 3, VA1: varillal alto sitio 1, VA2: varillal alto sitio 2, VA3: varillal alto sitio 3, VA4: varillal alto sitio 4.
Fuente: elaboración propia.

Comparación de la diversidad alfa del Varillal alto y Varillal bajo
La prueba de T de Student demuestra que hay una diferencia significativa entre la riqueza de especies de mariposas entre el Varillal alto y el Varillal bajo (p = 0,0129). Mientras que la prueba de Mann Whitney indica que no hay diferencia estadística significativa entre los valores del índice de Shannon-Wienner (p = 0,057).
Especies de interés para la conservación, educación ambiental y bionegocios
Entre las especies de interés para la conservación, educación ambiental y los bionegocios, encontramos a Morphinae que incluye a las seis especies del género Morpho ya que estas mariposas poseen una belleza muy particular, siendo las especies más abundantes Morpho helenor (Figura 3B), Morpho achilles y Morpho menelaus (Figura 3A). En el caso de las mariposas “búho”, presentan unos diseños muy particulares en sus alas ventrales de gran atracción, 4 especies fueron muy frecuentes Caligo euphorbus, Caligo eurilochus, Caligo idomeneus y Caligo placidianus. Entre los Satíridos destacan Cithaerias pireta aurorina (Figura 3E) y Haetera piera negra (Figura 3F) mariposas exclusivas del sotobosque cuya belleza radica en sus colores y formaciones alares transparentes. Todas estas especies se podrían incluir para futuros planes de manejo con fines turísticos y de exportación.
Discusión
El presente estudio muestra una riqueza de 24 especies de Satyrinae para el Varillal alto y bajo en la Reserva Nacional Allpahuayo Mishana (RNAM); por su parte, Campos y Ramírez (2005) reportan 37 especies para la misma zona, esta superioridad puede deberse a la aplicación de diferentes atrayentes (cebos) y trampas de colecta. En otros lugares amazónicos se reportan mayores registros de especies: 103 especies para la zona de Pakitza (Robbins et al., 1996), mientras que Lamas et al. (1996) indica 51 especies para el Alto Río Napo en Huamanpata (Cordillera subandina del Departamento de Amazonas). Lamas y Campos (2006) reportaron 29 especies, siendo inferior a lo encontrado por Campos y Ramírez (2005) en la RNAM (37 especies) y ligeramente superior a nuestros resultados: 24 especies en la misma zona, a pesar de la mayor cantidad de hábitat muestreados (9 tipos de hábitat). En cambio, nuestros datos son muy similares a los reportados por Novoa y Montero (2010) quienes reportaron 26 especies de Satyrinae para la zona de San Rafael. Estos registros reportados para la selva son superiores a los de la sierra y costa, tal como indican Cerdeña et al. (2014) quienes encontraron solo siete especies para Tacna (en el extremo sur del Perú); por su parte Pyrcz et al. (2011) encontraron 19 especies en Apurímac; y para el caso de la costa, Lamas (1976) solo registró 4 especies para Tumbes (en el extremo norte del Perú). La amazonia de Colombia muestra registros interesantes para una zona de baja Amazonia del Putumayo: 27 especies de Satyrinae (Salazar, 1995). En otra zona de Bosque bajo a medio denso, con depósitos aluviales registraron 50 especies de Satyrinae (Salazar et al., 2010); sin embargo, la zona de mayor riqueza lo reporta Andrade-C et al. (2015), para la Región Amazónica Colombiana con 78 especies de Satyrinae.
Para los Morphinae reportamos 14 especies, siendo Morpho telemachus un nuevo registro para la RNAM, el cual no se reportó en el estudio anterior de Campos y Ramírez (2005). Esto constituye un aporte de la presente investigación. Sin embargo, este registro es inferior a lo reportado por Campos y Ramírez (2005) quienes indican 24 especies para la misma zona. Por su parte, Novoa y Montero (2010) reportan datos muy similares a nuestros resultados para la zona de San Rafael quienes encontraron 16 especies de Morphinae, siendo este dato similar a lo indicado por Lamas et al. (1996) que reporta 15 especies para el Alto Río Napo en Loreto. Los registros más altos los indica Lamas (1984) y Robbins et al. (1996) para la zona de Tambopata y Pakitza con 32 y 31 especies respectivamente. Siendo el registro más bajo con una sola especie la de Huamampata (Lamas y Campos, 2006) y Tumbes (Lamas, 1976). La mayor cantidad de especies registradas en estos sitios de muestreos asumimos a dos factores principales: la metodología empleada y la diversidad de hábitat. En cuanto a los registros de Morphinae de las zonas de Colombia, Salazar (1995) reporta 8 especies de Morphinae y 5 especies de Brassolinae para la zona de baja Amazonia del Putumayo. En la zona de Bosque bajo a medio denso, con depósitos aluviales Salazar et al. (2010) registraron 2 especies de Morphinae y 15 especies de Brassolinae, por su parte Andrade-C et al. (2015) indican 26 especies de Morphinae y 28 especies de Brassolinae para la Región amazónica colombiana. Estos datos son superiores a nuestros resultados; sin embargo, es importante señalar que para la captura estos autores emplearon diversas redes entomológicas telescópicas, además de trampas Van Someren-Rydon realizaron visualización directa con binoculares de campo.
Cithaerias pireta aurorina (Satyrinae) fue una de las tres especies abundantes de nuestro estudio coincidiendo con los resultados de Campos y Ramírez (2005). Haetera piera negra fue abundante en el muestreo de Campos y Ramírez (2005), nosotros la reportamos en baja densidad; no obstante, las especies Pierella lena y Pierella lamia fueron más abundantes en nuestro caso, contrario a lo reportado por Campos y Ramírez (2005), esto se debe principalmente a la sincronización del ciclo de vida de las mariposas, la que está influenciado por la disponibilidad de su alimento tal como lo menciona Melo (2011). Así también consideramos que los patrones temporales de abundancia de mariposas se deben a una variación en la dinámica de sus plantas alimenticias, especialización de la alimentación o a una variación temporal en la mortalidad de las larvas (Hamer et al., 2005; Checa et al., 2009).
La cantidad de especies es diferente entre el varillal alto y el varillal bajo (T de Student; p = 0,012) y la homogeniedad entre la cantidad de individuos por especie (Shannon-Wienner) son iguales (Mann Whitney; p = 0,057); esto también representa una nueva información en comparación con la publicación de Campos y Ramírez (2005). Es posible que la estructura del varillal alto este influenciando en su mayor cantidad de especies, esta condición puede deberse a la mayor altura de las plantas de 10 a 15 m, mientras que las plantas del varillal bajo presentan una altura de 8 a 10 m. propiciando que una comunidad vegetal pueda albergar más especies mientras mayor cantidad de nichos disponga; nuestros resultados indican similitud con el trabajo de Campos y Ramírez (2005), lo cual muestra que después de una década las especies se siguen conservando, indicando que los bosques de la Reserva Nacional Allpahuayo Mishana (RNAM) están cumpliendo su función de conservación de la diversidad de las mariposas.
Entre las especies de mayor impacto en los bionegocios tenemos a los del género Morpho destacando las especies Morpho menelaus, Morpho helenor y Morpho achilles por ser las más comunes; datos similares fueron encontrados por Núñez (2009) para Iguazú en Argentina que indica a Morpho helenor y Morpho menelaus las especies más comunes en esta zona. Consideramos necesario desarrollar estudios relacionados a sus aspectos biológicos para ser manejados en condiciones de cautiverio con fines de conservación y bionegocios.
Conclusiones
Entre los varillales evaluados se encontró una baja y media diversidad de mariposas, siendo la subfamilia Satyrinae ligeramente superior a la sub-familia Morphinae. La composición de los Satyrinae supera en casi el doble de especies a los Morphinae, siendo P. lena la más abundante y la menos abundante C. penelope (Satyrinae); en los Morphinae la más abundante fue M. helenor y la menos abundante M. telemachus. En cantidad de individuos por tipos de bosques, los Satyrinae fueron superiores, su presencia ocurrió en todos los sitios evaluados, mientras que, para los Morphinae, la más abundante fue M. helenor y la menos abundante M. telemachus. La diversidad alfa muestra que el varillal alto presenta mayor riqueza de especies que el varillal bajo (T de Student; p = 0,0129); sin embargo, en la cantidad de individuos por especie las dos zonas son homogéneas (Mann Whitney; p = 0,057).
En relación a las especies de interés para la educación ambiental y los bionegocios, encontramos que las especies más bellas y exuberantes fueron M. menelaus, M. achilles y M. helenor, las más frecuentes entre las mariposas “búho” (C. idomeneus, C. eurilochus, C. euphorbus y C. placidianus) cuyas alas presentan diseños muy particulares. Los Satyrinae que destacaron por la belleza con colores muy llamativos fueron C. pireta aurorina y H. piera negra. Es necesario elaborar planes de manejo incluyendo a todas estas especies con el propósito de implementar actividades de conservación y manejo para su aprovechamiento con fines educativos, turísticos y de exportación.

