











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Tectona grandis L., comúnmente conocida como teca, es una especie forestal de la familia Lamiaceae, cultivada en casi todas las regiones tropicales. Es considerada como una de las especies de madera dura más valiosa del mundo y es buscada por su belleza, fuerza, estabilidad y resistencia naturales (Kollert & Kleine, 2017). En Costa Rica, es la especie más reforestada, alcanzando aproximadamente 47 167 ha plantadas y más de 150 000 árboles dispersos en fincas (INEC, 2015).
Para el trópico Americano, Arguedas et al. (2013) reportan 34 patógenos que causan enfermedad en plantaciones de teca, entre los que mencionan especies del género Fusarium y Armillaria, relacionados con la pudrición de la raíz.
En estudios recientes realizados en México y Brasil, se han registrado árboles de teca que presentan muerte descendente con pudrición de la base del tronco y la raíz. Las hojas de los árboles enfermos muestran amarillamiento, marchitez y posterior defoliación y muerte. El cuello de la raíz muestra pudrición y desprendimiento de la corteza, y en ocasiones se observan sobre el síntoma cuerpos fructíferos de color gris a blanco brillante. De estos árboles, se ha aislado el patógeno Kretzschmaria zonata (Cibrián et al., 2014;Alfenas et al., 2021).
También se han reportado síntomas similares a los descritos anteriormente en plantaciones de teca en la provincia de Guanacaste, pero se desconoce el agente causal. Por tal motivo, el objetivo de esta investigación fue determinar, a nivel morfológico y molecular, el patógeno causante de la pudrición del cuello de la raíz de Tectona grandis en Costa Rica.
MATERIALES Y MÉTODOS
En tres plantaciones de teca ubicadas en la provincia de Guanacaste, específicamente en Cañas 10 ° 23’ 45’’ N, 85° 01’ 57’’ W (13 años), Limonal 10 ° 17’ 20’’ N, 85° 01’ 51’’ W (7 años) y Coyolito 10 ° 07’ 10’’ N, 84° 55’ 57’’ W (5 años), se registraron árboles con síntomas de pudrición en el cuello y la raíz.
Se cortaron secciones cercanas a la base de estos árboles enfermos, y se trasladaron discos de madera de tres pulgadas de ancho al Laboratorio de Patología Forestal del Instituto Tecnológico de Costa Rica, donde se lavaron con jabón antibacterial y abundante agua. Posteriormente, se cortaron secciones desde la zona de avance y, dentro de la cámara de flujo laminar, se desinfestaron con alcohol al 70 % durante un minuto, con hipoclorito de sodio al 5% durante dos minutos y, acto seguido, por tres enjuagues consecutivos con agua destilada estéril (ADE). Con toallas de papel absorbentes, se secaron los segmentos, se colocaron en placas de Petri que contenían un medio de cultivo papa dextrosa agar (PDA) con antibiótico (penicilina 0.2 g.L-1 + estreptomicina 0.2 g.L-1 + chloramphenicol 0.2 g.L-1) y se encubaron a 25 °C. Después de la incubación, se obtuvieron colonias individuales, transfiriendo puntas de hifas a medio de cultivo PDA con antibiótico.
La identificación morfológica se realizó utilizando microscopía óptica (MO, Nikon Eclipse -Ni-). Los peritecios se colocaron en ácido láctico, y se midió el ancho y la longitud de 30 ascosporas. Se calcularon los tamaños máximo y mínimo, la media, la desviación estándar y los intervalos de confianza del 95 %.
En el Laboratorio de Técnicas Moleculares Aplicadas a la Fitoprotección de la Universidad de Costa Rica, se realizó extracción de ADN, seguida de PCR con cebadores fúngicos universales (ITS4 e ITS5), en aras de identificar los aislados, los cuales se enviaron a secuenciar a Macrogen en Corea del Sur. Las secuencias de consenso se compararon con las accesiones del GenBank utilizando la herramienta NCBI Blast, y el análisis de unión de vecinos se llevó a cabo con la versión 11 de MEGA (Tamura et al., 2021).
Las pruebas de patogenicidad se realizaron en diez plantas de teca. Cinco plantas se inocularon con un aislado de Kretzschmaria zonata. Se realizó un corte en la base del cuello de la planta para exponer el cambium, y se puso en contacto con un tapón de micelio del hongo creciendo en PDA. La herida se cubrió con Parafilm para evitar la disecación del organismo. Las otras cinco plantas se inocularon con PDA estéril. Diez días después de la inoculación, se midió el avance de la enfermedad con un caliper digital.
Las plantas con síntomas se reaislaron para confirmar los postulados de Koch.
RESULTADOS
En las tres plantaciones de teca estudiadas, se encontraron síntomas similares en árboles enfermo. Se observó pudrición en la base del tronco, y en ocasiones se logró visualizar el cuerpo fructífero del patógeno (Figura 1a). También se observó una reducción del área foliar (Figura 1b), pudriciones en la raíz (Figura 1c) y líneas de pudrición interna en la madera del cuello (Figura 1d).

Figura 1 Síntomas encontrados en árboles enfermos de Tectona grandis. a) Pudrición en la base del tronco con cuerpos fructíferos del hongo, b) defoliación, c) pudrición de la raíz, d) líneas de pudrición encontradas en madera. Provincia de Guanacaste, Costa Rica.
Sobre la corteza de árboles con síntomas, se desarrollaron estromas duros de color gris oscuro con bordes blancos (Figura 2a). Dentro de los estromas, se observaron peritecios de color café oscuro (Figura 2b). Los aislamientos realizados en PDA mostraron que la colonia del hongo crecía de forma circuncéntrica, con una coloración gris en el centro y blanca en los bordes (Figura 2c). Se registraron ascosporas de color café muy oscuro, de forma elipsoidal con extremos frecuentemente estrechos y de tamaño (21.6-) 27.4 - 32.1 (-33.5) x (6.2-) 8.7 - 10.9 (-11.8) µm ((mínimo-) promedio menos desviación estándar - promedio más desviación estándar y (-máximo)). Además, dentro de ellas se observó una línea germinal recta (Figuras 2d a 2e).

Figura 2 Kretzschmaria zonata. a) Astroma, b) peritecios, c) crecimiento micelial en PDA, d y e) ascospora. Escala: 10 µm
La secuencia del gen ITS mostró un 99 % de similitud con K. zonata (número de accesión KY660541). Según el análisis filogenético de máxima verosimilitud, la secuencia Kretzschmaria zonata_CR1 se agrupó con K. zonata de otras regiones (Figura 3).
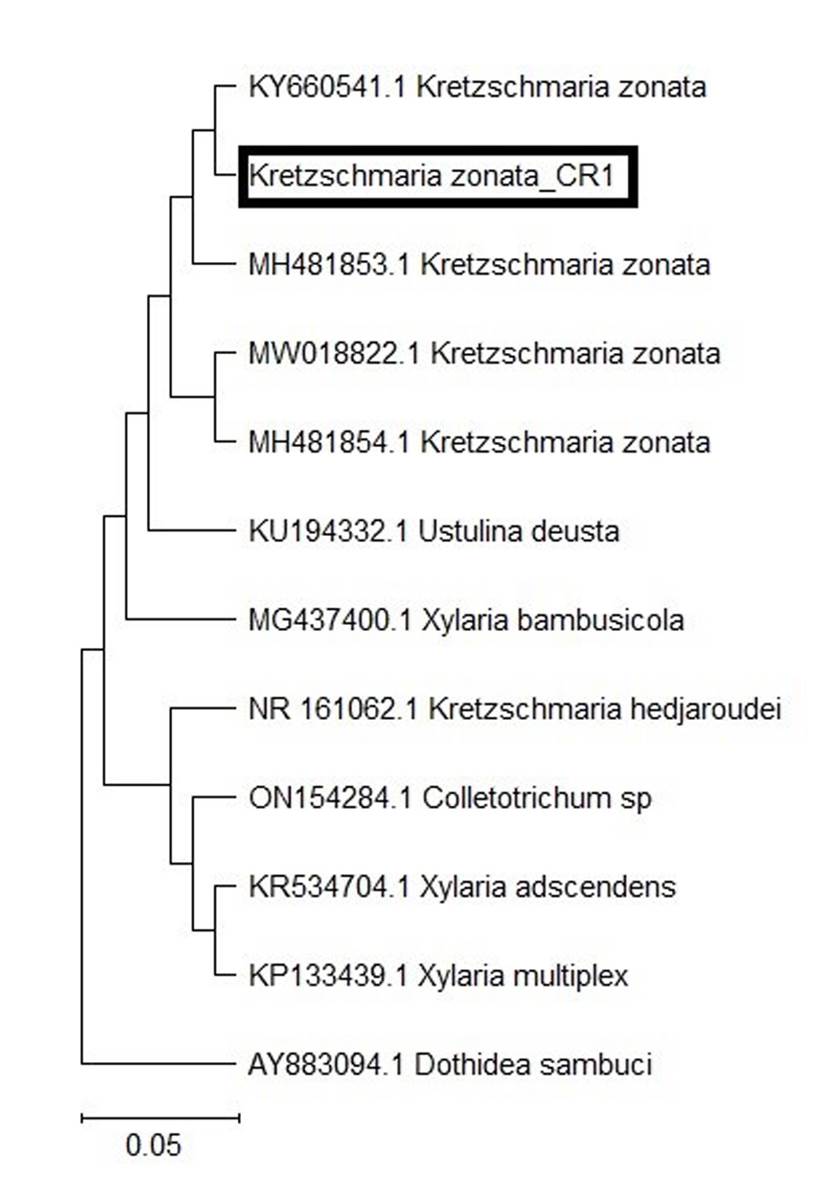
Figura 3 Árbol filogenético basado en secuencias de ADN ITS de aislados representativos de K. zonata y otras especies de Xylariaceaes. El árbol se enraizó con el aislado de Dothidea sambuci. K. zonata_CR1 está resaltada en el árbol.La barra de escala indica el número de sustituciones por sitio
En las pruebas de patogenicidad realizadas, los árboles inoculados con el aislado de K. zonata presentaron una lesión necrótica con una longitud promedio de 20.97 mm, mientras que, en los árboles testigo, la longitud de la lesión fue 5.09 mm 22 días después de la inoculación. Se encontró una diferencia altamente significativa entre ambos tratamientos, con un alfa de 0.05.
De los reaislamientos correspondientes, se recuperó el hongo Kretzschmaria zonata, cumpliendo con los postulados de Koch.
DISCUSIÓN
Las estructuras morfológicas del hongo, así como los análisis filogenéticos de su identificación molecular en la región del espaciador transcrito interno (ITS) del ADN, indican que el patógeno bajo estudio corresponde a Kretzschmaria zonata y coinciden con las características descritas por Alfenas et al. (2021) y Cibrián et al. (2014). Estos autores reportan que la especie ha causado los mismos síntomas de pudrición en la raíz y el cuello de árboles de T. grandis en Brasil y Campeche (México). También confirman que este patógeno puede afectar gravemente las plantaciones comerciales de teca, desarrollando una pudrición suave en la madera de los individuos, lo que disminuye la resistencia de la misma (afectando inclusive el duramen). Además, produce un anillamiento en el fuste que impide el flujo de savia desde la raíz hasta las zonas aéreas, provocando defoliación y, en algunos meses, la muerte de los individuos (Cibrián et al., 2014; Alfenas et al., 2021).
Del género Kretzschmaria se han descrito 28 especies distribuidas en regiones templadas y tropicales (Kirk et al., 2008). En la Reserva de la Biosfera Isla Cozumel, Quintana Roo, México, se registró K. zonata como fitopatógeno en árboles de Cedrela odorata (Raymundo et al., 2021). En China, se encontraron y describieron K. milleri y K. sandvicensis (Du et al., 2016). Por otro lado, en regiones templadas, se han analizado los síntomas causados por K. deusta en árboles de boques caducifolios. Los estudios aseguran que es un patógeno eficaz en la descomposición del tejido leñoso de la base y la raíz de sus hospederos (Innes et al., 2006; Büttner et al., 2017).
Para Costa Rica, este es el primer reporte de síntomas causados por K. zonata en plantaciones de teca, y es un avance importante para continuar con prácticas de monitoreo, así como en el establecimiento de pruebas estratégicas para el manejo integrado de la enfermedad.
CONCLUSIONES
En este estudio se identifica, por primera vez y a nivel morfológico y molecular, el patógeno Kretzschmaria zonata como el agente causal de la pudrición de la base del tronco y la raíz en árboles de Tectona grandis en Costa Rica. La patogenicidad se comprobó mediante los postulados de Koch. Este trabajo constituye un primer avance para el estudio de estrategias de manejo integrado de la enfermedad en las plantaciones de teca en el país.

