











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Los macroinvertebrados acuáticos altoandinos revisten interés científico por presentar adaptaciones particulares (Jacobsen, 2004). Esto es debido a que los ecosistemas acuáticos que los albergan, como los ríos altoandinos, presentan condiciones diferentes a las presentadas en otros ríos del mundo, tales como temperatura, materia orgánica, congelamiento, menor tensión de oxígeno, etc. No obstante, estas condiciones de climas extremos, su conocimiento es limitado, y específicamente por encima de los 4000 msnm, estos ambientes son aún poco estudiados. Esta información ha sido mejor documentada en ensambles bentónicos de cuencas bajas, donde el pulso hidrosedimentológico es el principal factor que estructura a las comunidades (Junk et al, 1989; Neiff, 1990). En el Perú, algunos trabajos han evaluado tales ambientes en bofedales y ríos altoandinos (Arana y Cabrera, 2017; Carrasco et al., 2020; Pumasupa et al., 2021; Quispe Mamani et al., 2021)
Los macroinvertebrados bentónicos, especialmente los insectos en sus estadios larvales son de gran importancia en un gran número de ambientes de aguas continentales (Hanson et al., 2010). Su importancia radica en que forman parte importante de casi todos los procesos ecológicos en los sistemas acuáticos (Hanson et al., 2010; Atkinson et al., 2019). Estudios sobre sus funciones en el ecosistema han sido realizados para diversos ambientes y se conoce que el ensamble de las comunidades de macroinvertebrados podría estar influenciado por gradientes altitudinales (Jacobsen et al., 2003: Lujan et al., 2013; Arana et al., 2021), variables ambientales (Jacobsen y Marín, 2008), interacciones ecológicas (Hurtado et al., 2005; Molina et al., 2008; Quimbayo Díaz et al., 2023) o por impactos antrópicos (Tognelli et al., 2016; Ramírez et al., 2018; Urdanigo et al., 2019; Ríos-Touma y Ramírez, 2019; Ríos-Touma et al., 2022).
Por ello, en este trabajo se busca incrementarla información sobre el ensamble de macroinvertebrados presentes en ambientes altoandinos y explicar sus variaciones por efecto de diversos factores relacionados a la estacionalidad como variables limnológicas, granulométricas e hidrológico-hidráulicas. Nosotros hipotetizamos que, en ambientes extremos, la estacionalidad debido al incremento del caudal modifica los ensambles bentónicos incrementando su densidad, y discutimos las características únicas de estos ambientes.
MATERIALES Y MÉTODOS
El estudio fue realizado en un tramo del río El Salto ubicado a 4552 msnm, en el distrito de Cayarani, Provincia de Condesuyos, departamento de Arequipa (14°59'41" S, 72°17'59" W a 14°59'39" S, 72°18'1" W) (Fig. 1). Según el mapa Nacional de Ecosistemas Minam (2018), el área de estudio pertenece al ecosistema altoandino de pajonal puna seca. Este ecosistema presenta vegetación herbácea constituida principalmente por gramíneas de porte bajo y pajonales dominados por algunas asociaciones arbustivas dispersas y gramíneas que crecen amacolladas, dispersas, y con de tallo y hojas duras. El río El Salto es un sistema fluvial de cabecera de cuenca que recibe contribuciones de numerosos bofedales de alto contenido orgánico. El patrón geomorfológico del canal se caracteriza por baja declividad, con canal estrecho rectilíneo y poca profundidad.
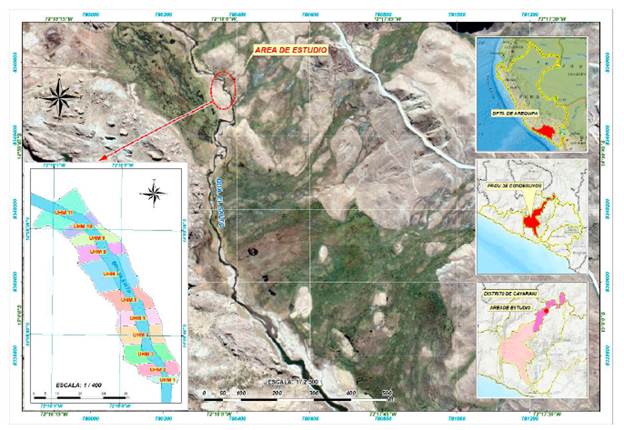
Se utilizó la metodología de tramo representativo, para seleccionar el tramo estudiado (Parasiewicz y Dunbar, 2001; Parasiewicz et al., 2009). Se recorrieron 3 km de río reconociendo el patrón de unidades hidromorfológicas (UHM), tomándose un tramo que reflejó la distribución proporcional de las mismas. Las UHM son tramos de río con características homogéneas, definidas por una determinada interacción entre sus características hidráulicas (velocidad, ancho de cauce, granulometría) y la topografía del lecho (profundidad). Estas características y otras variables de hábitat configuran un mesohabitat, una zona de río con particularidades que promueven la presencia y densidad diferencial de la biota acuática, según su conformación (Parasiewicz y Dunbar, 2001, Parasiewicz et al, 2009). Se establecieron 11 unidades (UHM), de una extensión variable según sus características hidromorfológicas, las cuales se encontraban una a continuación de la otra en un tramo de 90 m de longitud, siendo clasificadas según Parasiewicz et al. (2009), Bisson et al. 2006 y Dolloff et al. (1997) (Fig. 2 y Tabla 1). Se realizaron muestreos en dos temporadas: temporada seca (S) en setiembre de 2020 y temporada húmeda (H) en marzo de 2021.
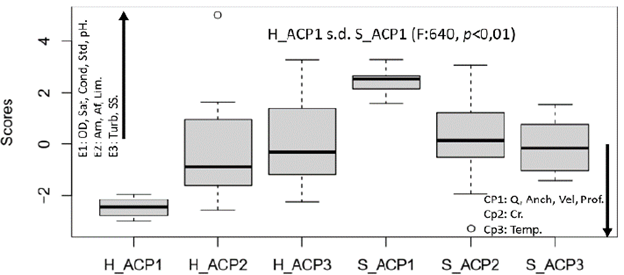
Figura 2 Dispersión de los escores de los ejes retenidos en el Análisis de Componentes Principales de las variables limnológicas, geomorfológicas e hidrológicas medidas en el río El Salto - Arequipa, en temporada seca (S) y húmeda (H). H_ACP1, H_ACP2 y H_ACP3, componentes principales 1, 2 y 3 en temporada húmeda. S_ACP1, S_ACP2 y S_ACP3, componentes principales 1, 2 y 3 en temporada seca. Las flechas indican correlación positiva (hacia arriba) y negativa (hacia abajo) de cada variable con su respectivo componente (EX1, EX2 y EX3), respectivamente. Temp: Temperatura, Turb: Turbidez, SS: Sólidos en suspensión, Prof: Profundidad, Q: Caudal, Anch: Ancho de cauce, Vel: Velocidad, Cond: Conductividad, Std: Sólidos totales disueltos, Sat: % Saturación, OD: Oxígeno disuelto, Cr: Canto rodado. Am: Arena media. Af: Arena fina. Lim: Limo.
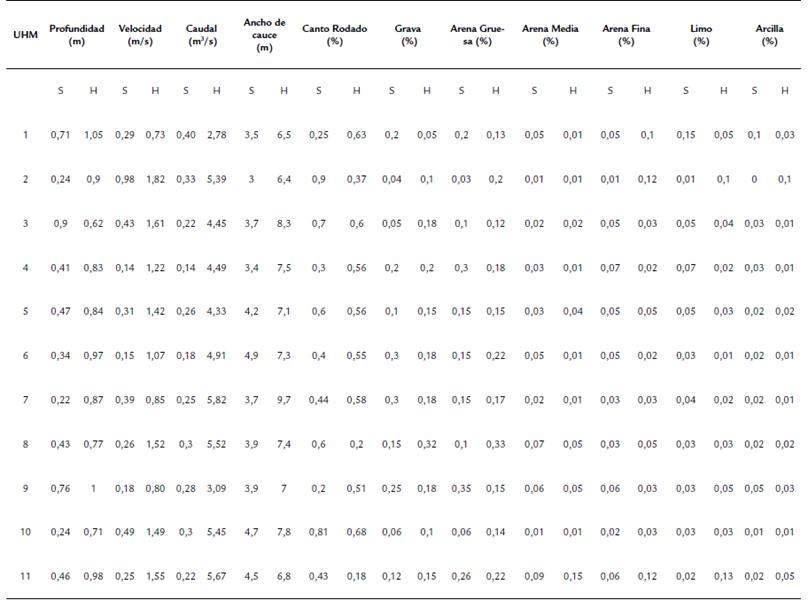
En cada UHM se registraron variables limnológicas fisicoquímicas como el oxígeno disuelto, porcentaje de saturación, conductividad, sólidos totales disueltos (STD), temperatura del agua, pH, turbidez y sólidos en suspensión (SS) utilizando un multiparámetro digital Bante900P, fotocolorímetro Hach DR900 y turbidímetro Lutron TU-2016 respectivamente. También se realizó la determinación del tenor granulométrico del sedimento (% de cantos rodados, grava, arena gruesa, arena media, arena fina, limo y arcilla) mediante el método de tamizado en mojado (Suguio, 2003) siguiendo la escala de Wentworth (1922) modificada. Mediante este método se obtienen resultados porcentuales en una escala cualitativa. También fueron medidas variables hidrológico-hidráulicas como caudal, velocidad, profundidad y ancho de cauce, tanto en temporadas seca como húmeda. La velocidad de flujo se midió utilizando un correntómetro Flowatch JDC de 0,01 de resolución. El ancho del cauce se midió con un distanciómetro digital Leica Disto D100. Se colectaron muestras de macrobentos en cada UHM, utilizándose una red Surber de marco metálico de 30 x 30 cm con abertura de malla de 250 micras. La red Surber fue colocada contracorriente y el sustrato que se encontraba dentro del marco metálico fue lavado y los organismos que quedaron retenidos en la red fueron almacenados en frascos de plástico conteniendo alcohol al 70%. En cada UHM se colectó dos veces con la red Surber. Los frascos posteriormente fueron llevados al laboratorio para realizar el tamizado de las muestras con malla de 250 micras y se identificaron los organismos hasta llegar al menor taxón posible utilizando las claves de identificación de Merritt et al. (2008), Domínguez y Fernández (2009) y Prat y Rieradevall (2014).
Las variables limnológicas fisicoquímicas, geomorfológicas e hidrológicas fueron resumidas en un análisis de componentes principales (ACP). Los ejes que retuvieron la mayor variabilidad fueron seleccionados con el criterio de Broken stick (Peck, 2016), siendo posteriormente analizados con permanova (Anderson, 2017), para determinar diferencias entre las temporadas evaluadas. Los valores de abundancia de organismos obtenidos con la red surber fueron convertidos a densidad (número de individuos por m2) y luego fueron promediados para obtener un valor de densidad para cada UHM en cada temporada. Con la densidad se calcularon los índices comunitarios como: diversidad (Shannon), Riqueza (Margalef), Equidad (Pielou) y Dominancia de Simpson (D) en el programa Past 4.08 (Hammer et al., 2001). Los valores de densidad por UHM fueron evaluados mediante Escalamiento multidimensional no métrico (nMDS) utilizando la distancia de Bray-Curtis como medida de distancia. Las diferencias entre los ejes significativos del nMDS en temporada seca y húmeda se evaluaron a través de permanova. (Anderson, 2017). La correlación entre las matrices abiótica y biótica se evaluó seleccionando los ejes significativos del ACP y nMDS y utilizando nmds (Peres-Neto y Jackson, 2001). Tanto esta técnica como los análisis multivariados se realizaron utilizando el paquete vegan (Oksanen et al, 2018) de la consola RStudio (RStudio Team, 2020) y R (R Core Team, 2014).
RESULTADOS
El ACP retuvo tres ejes que explicaron el 71,64 % de la variabilidad (CP1=36,282 %, CP2= 22,36 % y CP3= 13,22 %, respectivamente). En el CP1 fueron retenidas el oxígeno disuelto, porcentaje de saturación, pH, conductividad, STD, correlacionadas positivamente, y el caudal, el ancho de cauce, la velocidad y la profundidad correlacionadas negativamente. El CP2 retuvo la variabilidad de las variables granulométricas y el CP3 la variabilidad de la turbidez, SS y temperatura (Figura 2). Se encontraron diferencias estadísticamente significativas entre la temporada seca y la temporada húmeda en CP1, (F: 640,1, p<0,01). Las variables retenidas en el CP1 tuvieron un rango de variación marcado entre la temporada seca y la temporada húmeda (tablas 1 y 2). Así, la velocidad de corriente (0,35 a 1,28 m/s) como el caudal (0,26 a 4,72 m3/s) fueron menores en temporada seca que en temporada húmeda. Por otro lado, el porcentaje de saturación, la conductividad, el total de sólidos disueltos y el pH, fueron mayores en la temporada seca que en la húmeda. A pesar de que el eje en que fueron retenidas no mostró diferencias estadísticamente significativas entre las temporadas de muestreo, la temperatura, la turbidez y sólidos en suspensión también tuvieron rangos de variación relativamente altos. (Tabla 1 y 2). En cuanto a las variables granulométricas se observó que para ambas temporadas la partícula predominante fue el canto rodado, seguido de la arena gruesa y grava. Los valores de las variables limnológicas, granulométricas e hidrológico-hidráulicas para las dos temporadas evaluadas se observan en las tablas 1 y 2. Respecto a la clasificación hidromorfológica de las UHM, se observó el cambio de unidades de flujo lento a unidades de flujo rápido de la estación seca a la estación húmeda. Así las UHM de tipo tabla y poza (flujo lento) pasaron a run y riffle (UHM1, 4 y 9), mientras que las UHM 6 y 7 pasaron de tabla a ruffle respectivamente.
Tabla 1 Tipos de UHM y valores de las variables limnológicas medidas en las unidades hidromorfológicas (UHM) del 1 al 11 en el río El Salto - Arequipa, en temporada seca (S) y húmeda (H). TSD= Sólidos totales disuelto, NTU = Unidades nefelométricas de turbidez, SS= Sólidos en suspensión. Definición de UHM según Parasiewicz et al. (2009), Bisson et al. 2006 y Dolloff et al. (1997).
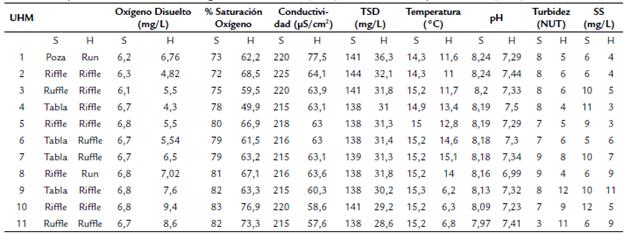
Tabla 2 Valores de las variables geomorfologicas e hidrologicas-hidraulicas medidas en 11 unidades hidromorfologicas (UHM) del rio El Salto - Arequipa, en temporada seca (S) y Humeda (H).
El ensamble macrobentónico estuvo conformado por 29 morfoespecies en total, siendo 23 especies registradas en temporada seca y 20 en la húmeda (Tabla 3). La densidad varió de 43756 individuos en temporada seca a 8722 individuos en la húmeda. Los valores de diversidad, riqueza y equidad fueron mayores en temporada húmeda, mientras que la dominancia fue mayor en temporada seca. (Fig. 2)
Tabla 3 Densidad total (Individuos por m2) de macroinvertebrados registrados en 11 unidades hidromorfologicas (UHM) en el rio El Salto - Arequipa, en temporada seca (S) y humeda (H).
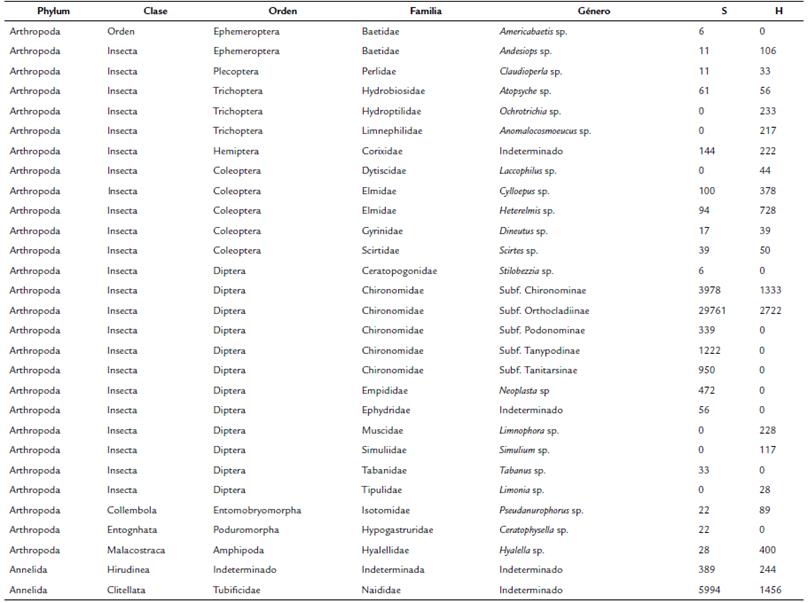
Las mayores densidades fueron registradas para la subfamilia Orthocladiinae (Familia Chironomidae) (29761 y 2722 individuos en temporada seca y húmeda, respectivamente) y para morfoespecies de Haplotaxida (5994 y 1546 individuos en temporada seca y húmeda, respectivamente). Fueron también registrados órdenes de insectos coleópteros, trichópteros y hemípteros, con mayores valores de densidad en la temporada húmeda. También se registró un mayor número especies de dípteros en la temporada seca.
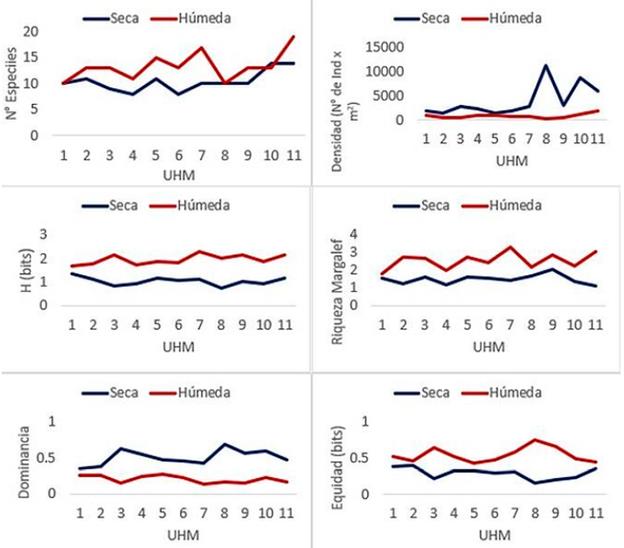
Figura 3 Número de especies, abundancia, diversidad, riqueza, dominancia y equidad de los macroinvertebrados registrados en 11 unidades hidromorfológicas (UHM) del río El Salto - Arequipa en temporada seca (S) y húmeda (H).
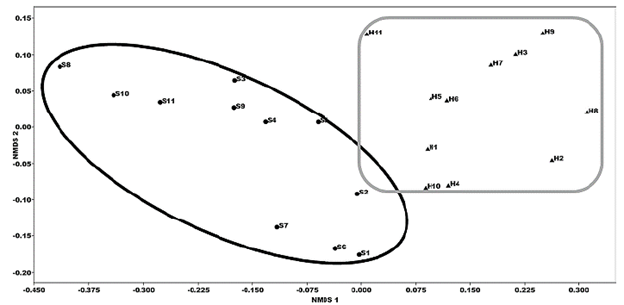
El análisis de ordenación (nMDS, stress=0.081) separó a las unidades de muestreo de ambas temporadas indicando cambios en la estructura del ensamble macrobentónico. (Fig. 3). Solos los escores del eje 1 del nMDS presentaron diferencias estadísticamente significativas entre las temporadas seca y húmeda (F:2162, p>0,001). Las matrices biótica y abiótica estuvieron altamente correlacionadas Procrustes suma de cuadrados (m2) = 0,3981, correlación en rotación simétrica Procrustes = 0,7759, p<0,001.
DISCUSIÓN
Debido al régimen hidrológico, en nuestro estudio, el caudal es la variable maestra que modifica los sólidos en suspensión, turbidez, pH, oxígeno disuelto, sólidos disueltos, temperatura, entre otras variables. Estas alteraciones promueven cambios en el ensamble del macrobentos en cuanto a su estructura. Este patrón de cambio ha sido referido en otros ríos altoandinos (Pérez y Segnini, 2005; Ríos-Touma, 2008; Villamarín et al., 2020).
En temporada húmeda, con el incremento del caudal, se evidenció una disminución en la densidad del macrobentos comparada a la temporada seca. Sin embargo, los índices comunitarios de diversidad y la riqueza fueron mayores, debido a la disminución de la dominancia y aumento de la equidad. Esta relación de disminución de la densidad con el incremento de variables hidrológico-hidráulicas es un patrón común a ríos con recurrencia de inundaciones periódicas (Arenas-Ibarra et al, 2012). La relación inversa entre la densidad y riqueza de macroinvertebrados con las variables hidrológicas-hidráulicas, así como entre la abundancia y riqueza con la profundidad y la velocidad de flujo en ríos altoandinos han sido anteriormente documentados (Molina et al., 2008; Oyanedel et al., 2008). Nuestros resultados confirman que las variables hidrológicas-hidráulicas modificarían fuertemente la estructura de las comunidades, tanto por los cambios que generan en la limnología fisicoquímica del agua, así como por su influencia en la deriva de diversas especies, teniendo un rol esencial en la colonización de los macroinvertebrados en diferentes microhábitats (Statzner y Higler, 1986; Ríos-Touma et al., 2012).
La conductividad, STD y turbidez presentan valores más altos en temporada seca, esto se explicaría debido a que el ambiente estudiado presenta un clima extremo (periodos de heladas donde las temperaturas descienden a 0°C o menos) y con características particulares e impredecibles (sequías) (Flecker y Feifarek, 1994). Más aún, el río El Salto está asociado a un bofedal, el cual lo alimenta y modificaría los valores de las variables limnológicas por la carga orgánica alta que presentan este tipo de ambientes (Osaki y Tsuji, 2015).
La abundancia y número de morfoespecies registradas en este análisis resultó ser baja comparada con otros ambientes altoandinos. Se sabe que en general, estos valores tienden a decrecer cuando incrementa la altitud y la latitud (Villamarín et al., 2020). Sin embargo, según Maldonado et al., (2011) este patrón de disminución de la diversidad con el aumento de la elevación en ríos andinos es aún poco claro. Es necesario entonces tomar en cuenta diversas características como sus conexiones a otros cuerpos de agua como bofedales, lo cual determina la formación de ensamblajes característicos (Carrasco et al., 2020), además de los cambios ambientales por actividades antrópicas (García- Ríos, 2016; Wang et al., 2021; Ríos-Touma et al., 2022) y adaptaciones morfológicas propias de los organismos a los cambios en las variables limnológicas (Hanson et al., 2010). Como ejemplo de esta complejidad, en este estudio, para la temporada seca se registró mayor abundancia de familias con altos requerimientos de oxígeno, como las EPT (familias Baetidae, Gripopterygidae, Hydrobiosidae, Hydroptilidae, Limnephilidae), sin embargo, las mayores concentraciones de oxígeno no necesariamente se dan en temporada seca, siendo que en cinco UHM la concentración de oxígeno fue mayor en temporada húmeda. Esto, en parte, estaría relacionado a la mayor solubilidad del oxígeno en bajas temperaturas y a la presión atmosférica. Estas condiciones se han presentado en diversos ambientes como quebradas del altiplano, donde se han presentado cambios drásticos en la saturación de oxígeno (Jacobsen et al., 2003). Por tanto, el cambio registrado no se manifiesta solamente en algunas variables, si no en el conjunto de características físico, químicas y biológicas que estructuran a las comunidades y condicionan sus cambios estacionales (Qazi y Ashok, 2012). Esta correspondencia entre los cambios en el sistema abiótico y el componente biótico tuvo alta correlación en el test procrustes de nuestro estudio. Por otro lado, si bien la clasificación hidromorfológica se realiza con criterio experto, es sugestivo que el cambio de tipo predominante de UHM de flujo lento (temporada seca) a flujo rápido (temporada húmeda) refleje fidedignamente los cambios en las condiciones de hábitat, abriendo un horizonte promisorio hacia la búsqueda de una clasificación más cuantitativa, que podría auxiliar en la comprensión de la relación caudal-hábitat-biota, como se realiza en estudios de preferencia de hábitat (Parasiewicz et al., 2013)
La mayor abundancia en ambas temporadas fue registrada para la familia Chironomidae (Orden Diptera), siendo mayor incluso en temporada seca. Los dípteros de esta familia son característicos de ambientes altoandinos y esto ha sido registrado en diversos ambientes (Ferru y Fierro, 2015; Custodio et al., 2018), debido a que tienen un rol muy importante en todos los niveles de las cadenas tróficas (Cummins et al., 2008). La subfamilia Orthocladiinae ha sido las más abundante en este estudio, lo cual también fue registrado por Acosta y Prat (2010) en las cabeceras del río Cañete, quienes resaltaron la complejidad de este grupo, la dificultad en su resolución taxonómica a nivel de géneros y más aún de especies.
El presente estudio confirma las tendencias anteriormente postuladas en cuanto a las comunidades de macroinvertebrados y su relación con las variables limnológicas, granulométricas e hidrológicas-hidráulicas. Éstas están más bien relacionadas a la temporalidad diferencial del ambiente lótico. Se confirma plenamente que estas comunidades están sometidas a fuertes cambios en el sistema físico que las estructuran estacionalmente. Si bien es cierto que las comunidades se encuentran adaptadas tanto morfológica y fisiológicamente a estos cambios, disturbios antrópicos en mayor escala podrían modificar esta dinámica, volviendo los ambientes acuáticos más vulnerables a pérdidas de biodiversidad.

