










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkORINOQUIA
On-line version ISSN 0121-3709
Orinoquia vol.14 no.1 Meta Jan./June 2010
Artículo Original /Original Article
Estudio clínico y hematológico de una infección experimental con Aeromonas hydrophila y Edwardsiella tarda en tilapia, Oreochromis sp
Clinical and hematological study of experimental infection with Aeromonas hydrophila and Edwardsiella tarda on tilapia Oreochromis sp
Mónica A. Vásquez-Piñeros1*, Iang S. Rondón-Barragán2, Luis F. Restrepo-Betancur3, Pedro R. Eslava-Mocha4
1Biologa Marina, Esp Acuicultura, cMSc, Grupo de Investigación en Sanidad de Organismos Acuáticos, IALL - Universidad de los Llanos
2 Médico Veterinario Zootecnista, cMSc Docente Universidad del Tolima, Departamento de Sanidad Animal, Grupo de Investigación en Inmunología y Fisiopatología Animal,
3 Docente Universidad de Antioquia, Facultad de Ciencias Agrarias, Grupo GRICA.
4 Médico Veterinario MSc. Docente Universidad de los Llanos *Email: mandreavasquez@gmail.com
Recibido: Febrero 17 de 2010. Aceptado: Mayo 18 de 2010
RESUMEN
Las bacterias oportunistas como A. hydrophila y E. tarda se relacionan con brotes de septicemia hemorrágica en tilapias de cultivo en nuestro país, siendo los agentes de mayor participación en enfermedades infecciosas en cultivos de Oreochromis sp en el Departamento del Meta. Este artículo reporta un estudio clínico y hematológico de tilapias híbridas (50 - 70g) durante la infección experimental con Aeromonas hydrophila y Edwardsiella tarda; realizada por medio de inoculaciones intraperitoneales de 7 x 108 UFC/ml y 2 x 108 UFC/ml, de A. hydrophila y E. tarda respectivamente. A partir de las 11 horas los peces infectados presentaron: palidez, letargo, boqueo, aumento de la frecuencia opercular y pérdida de eje de nado. En el examen hematológico, los animales infectados presentaron linfopenia severa, aumento de monocitos, hemólisis, abundantes células blásticas, marcada carriorrexis, hipocromía y anisocitosis, características de una anemia hipocrómica y eritroclasia descrita en infecciones bacterianas en peces. Al examen de necropsia, los peces inoculados con las bacterias evidenciaron: hemorragias en la base de las aletas y órganos internos, palidez e hígado moteado, aumento de líquido abdominal y líquido sanguinolento en el intestino, a diferencia de animales control que no presentaron estas lesiones. Adicionalmente, se observó hepatomegalia en animales infectados, aunque no hubo diferencias significativas en el índice hepatosomático de estos animales con respecto a animales control. En el tratamiento de E. tarda, en el bazo se observaron nódulos blanquecinos de distribución multifocal. Durante el presente estudio tilapias hibridas infectadas con E. tarda y A. hydrophila, mostraron un cuadro clínico compatible con septicemia hemorrágica, posiblemente ocasionado por a productos extracelulares (PEC) tales como hemolisinas y aerolisinas con actividad hemolítica y citotóxica en peces, entre otros factores. Este trabajo constituye una aproximación a un modelo de estudio de enfermedad bacteriana de tilapias con cepas patógenas aisladas en la región.
Palabras clave: Bacteria, hematología, septicemia, tilapia híbrida.
ABSTRACT
Opportunistic bacteria as A. hydrophila and E. tarda are related to haemorrhagic septicaemia outbreaks in cultured tilapias in our country, being the aetiological agents of highest participation on the diagnosis of infection disease in cultures of Oreochromis sp., in Departamento del Meta. This article reports a clinical and hematological study of tilapia hybrid (50-70g) during the experimental infection with Aeromonas hydrophila and Edwardsiella tarda made by intraperitoneal inoculation of 7 x 108 CFU/ml and 2 x 108 CFU/ml of A. hydrophila and E. tarda, respectively. From 11 hours infected fish showed paleness, lethargy, gasping, increased respiratory frequency and loss of swim axis. At hematological examination, infected animals showed severe lymphopenia, increased monocytes, hemolysis, increased blastic cells, karyorrhexis, hypochromia and anisocytosis, which is characteristic of a hypochromic anemia and erythroclasia described in fish bacterial infection. At necropsy examination, fish inoculated with bacteria showed: hemorrhages on the base of fins and internal organs, pale and mottled liver, increased abdominal fluid and blood-stained fluid in intestine, unlike control animals that did not show these lesions. Additionally, hepatomegaly was observed in experimental infected animals, even though there were not significance differences in the hepatosomatic index of these animals with respect to control. In treatment of E. tarda, whitish nodules with multifocal distribution were observed in spleen. During the present study, hybrid tilapias infected with E. tarda y A. hydrophila, showed a clinical condition compatible with haemorrhagic septicaemia, possibly associated to extracellular products (ECP), such as hemolysins and aerolisyns, with hemolytic and cytotoxic activity in fish, among other factors. This work is an approximation to a model of study of bacterial disease of tilapia with pathogenic strains isolated in the region.
Key words: Bacteria, hematology, septicaemia, Oreochromis sp., tilapia hybrid.
INTRODUCCIÓN
Aeromonas hydrophila y Edwardsiella tarda, se consideran entre los agentes etiológicos más importantes de enfermedades bacterianas en peces. Estas bacterias oportunistas hacen parte de la flora intestinal normal de peces y otros animales acuáticos (Cipriano, 2000); así que la presencia de estas, no es indicativo de enfermedad, factores ambientales y fisiológicos asociados a estrés contribuyen a la presentación de brotes de enfermedad y en consecuencia a altas pérdidas económicas dentro de los cultivos de peces. La patogénesis de la enfermedad por A. hydrophila y E. tarda es multifactorial y puede involucrar el producto de numerosos y diferentes genes que actúan individual y conjuntamente, codificando la síntesis de diferentes factores de virulencia y permitiendo a las bacterias invadir las células del hospedero y diseminarse vía sanguínea a diferentes (Chopra et al., 1999).
Tanto A. hydrophila como E. tarda son responsables de procesos de septicemia hemorrágica, un cuadro de enfermedad que afecta gran variedad de especies poiquilotermas. La septicemia hemorrágica se caracteriza por la diseminación generalizada en forma de bacteremia, seguida por la síntesis de toxinas, necrosis tisular y posterior fase clínica (Inglis, 1993). Las lesiones inducidas incluyen: hiperemia de capilares, exudación, hemorragias petequiales focales y leucocitosis, que es acompañada generalmente por niveles importantes de macrófagos circulantes pigmentados derivado de centros melanomacrófagos, así como exoftalmia asociada con edema pre-orbital (Roberts, 1989).
Aeromonas hydrophila y otras aeromonas motiles están entre las bacterias más comunes en los hábitats de agua dulce del mundo. Cepas de A. hydrophila aisladas de procesos infecciosos, han mostrado la presencia de diferentes genes que intervienen en la patogenicidad de esta bacteria como los son: la serina proteasa, asociada con la activación de la aerolisina; el colesterol aciltransferasa GCAT (por sus siglas en ingles); genes codificadores de lipasa como plc, lipH3, pla, lip y genes de DNasa (Chacón, 2003). Adicionalmente se ha determinado la producción de una variedad de productos extracelulares (PEC) biológicamente activos, como enterotoxinas citotóxicas que causan un daño importante en el epitelio, principalmente de intestino. Estas enterotoxinas también se han descrito por los investigadores como b-hemolisinas y aerolisinas.
Otros factores de virulencia reportados para A. hydrophila son: la producción de proteasas que contribuyen a la utilización de nutrientes para la proliferación celular y que causan daño al tejido, presencia de adhesinas, pili, flagelina, la presencia de moléculas secuestradoras de hierro denominas sideróforos (ligando específico de hierro de bajo peso molecular) (Murugappan et al., 2006), y la presencia de la capa S, arreglos de un tipo de proteína de superficie que forman una estructura supramolecular que encierra la bacteria y permite la interacción con tejidos y fluidos corporales, influencia la interacción hospedero-patógeno y se ha probado que su presencia aumenta la letalidad en peces (Kostrzynska et al., 1992).
Edwardsiella tarda es un agente causal de la septicemia en peces de aguas cálidas, en Estados Unidos y Asia se ha reportado principalmente en la anguila japonesa Anguilla japonica, tilapia nilótica Oreochromis niloticus, bagre de canal Ictalurus punctatus, y lenguado japonés, Paralichthys olivaceus (Mohanty y Sahoo, 2007). En Colombia, se ha reportado principalmente en Oreochromis sp., de cultivo (Iregui et al., 2004). Esta bacteria se ha diseminado ampliamente en animales acuáticos, estanques y lodos, lo que provee una oportunidad para la re-infección en peces de agua dulce de cultivo (Inglis et al., 1993). Se ha reportado que E. tarda invade las células epiteliales de intestino y piel; es capaz de producir hemolisinas, condroitinasa, sideróforos, es resistente al suero y a procesos fagocíticos (Morohshi et al., 2004), debido a la producción de enzimas, como: catalasa, peroxidasa y superóxido dismutasa (Mohanty y Sahoo, 2007), que detoxifican especies intermediarias de oxígeno y evitan los mecanismos oxidativos de destrucción, incluyendo la explosión respiratoria.
Adicionalmente, se ha reportado para ambos patógenos que la expresión de varios factores de virulencia y la división celular de estas bacterias, puede ser controlado por Quorum sensing, que es el mecanismo de control de expresión génica en respuesta a la expansión de la población bacteriana, factor que contribuye significativamente a la patogénesis (Morohshi et al., 2004; Chopra y Houston, 1999). Por tanto la proliferación de estas bacterias depende íntimamente de la intercomunicación entre ellas, exclusión competitiva así como de la disminución de la inmunocompetencia del hospedero.
La tilapia híbrida es la principal especie de cultivo del país; para el año 2004 Colombia se ubicó dentro de los diez principales productores de tilapia híbrida (FAO 2006). Este mercado en crecimiento de los últimos años, estuvo acompañado de una mayor intensificación de los sistemas de cultivo de esta especie, incremento en densidades, alimentación, así como en la carga orgánica en las aguas del cultivo; presentándose una considerable presión sobre estos sistemas de cultivo y posicionando a la tilapia como una de las especies más afectadas por infecciones por Aeromonas spp., Edwardsiella spp. y Streptococcus spp. (Rey, 2002).
En el Departamento del Meta, la mayor prevalencia en el diagnóstico de infecciones bacterianas en peces, fue reportada sobre A. hydrophila y E. tarda (Iregui et al., 2004)
Este trabajo tiene como objetivo aproximarse a un modelo de estudio de la dinámica de la enfermedad por bacterias oportunistas (A. hydrophila y E. tarda), aisladas de tilapias de cultivo del Departamento del Meta, a través de infecciones experimentales y el análisis de cuadros clínicos, lesiones macroscópicas y características hematológicas causadas.
METODOLOGÍA
Animales experimentales
Veinticuatro alevinos de Oreochromis sp., con peso entre 50-70 gramos provenientes de una granja comercial del Departamento del Meta fueron trasladados al Laboratorio de Bioensayos del Instituto de Acuicultura de los Llanos (IALL), Universidad de los Llanos. Los alevinos fueron ubicados en acuarios de vidrio de 64 L; mantenidos a una densidad no mayor a 1 g/L, con aireación constante, sin filtro y bajo condiciones controladas. Los animales se sometieron a un periodo de aclimatación de 15 días, alimentados al 1 % de la biomasa/día, con recambio de agua del 50 % diario.
Cepas bacterianas
Se realizó cultivo y aislamiento de A. hydrophila y E. tarda a partir de riñón, hígado y bazo de tilapias de cultivos del Departamento del Meta, con signos clínicos de enfermedad. La enfermedad se diagnosticó en el Instituto de Acuicultura de los Llanos; las cepas fueron caracterizadas de acuerdo a su fenotipo y pruebas bioquímicas. La especie de cada cepa se identificó por medio del Kit comercial BBL Crystal® (BD Diagnostic Systems, USA). Para su conservación las cepas bacterias fueron congeladas a -20°C en viales de caldo BHI (Brain Heart Infusion) con glicerol al 10%.
Virulentación y preparación de inóculos
Las cepas fueron virulentadas mediante pasajes por tilapia roja a través de la inoculación intraperitoneal de 100 µl de una solución madre de aproximadamente 109 UFC/ml (según Escala de Mac Farland), para posterior aislamiento de las bacterias a partir de órganos como: hígado, riñón y bazo, en agar BHI suplementado con sangre de ovejo al 5 %, incubando a 28°C por 24 horas (Jiménez et al, 2008).
Una vez obtenida la cepa virulenta, las concentraciones experimentales se cuantificaron a través de diluciones seriadas (1:10) en solución salina al 0.85 % y posterior incubación a 28°C por 24 horas para conteo en placa en agar triptona soya (TSA) (Jiménez et al., 2008)
Para las inoculaciones se tomaron dosis infectivas por encima de la dosis letal 50 para quince días en peces, reportada como 105 UFC/ml (Rey, 2002), para asegurar la infección y manifestación temprana de la enfermedad.
Infección experimental
Se dispusieron 3 tratamientos para la infección experimental: el primer grupo (T1), se inoculó intraperitonealmente con una solución de 7 x 108 UFC/ml de A. hydrophila; el segundo grupo (T2), se inyectó intraperitonealmente con 2 x 108 UFC/ml de E. tarda; y el grupo control (T0), se trató con solución salina estéril al 0.85 %, bajo el mismo protocolo de los grupos tratamiento. Cada grupo constó de 8 animales por unidad experimental. Durante el periodo de desafío los peces se mantuvieron a una densidad de 20 g/l con el fin de generar condiciones de estrés. Los alevinos se mantuvieron en observación por un periodo de 24 horas, durante el cual se realizó la evaluación clínica. Se tomaron muestras de animales con pérdida total del eje de nado y al final de este periodo se realizó la eutanasia y se tomaron muestras de los animales que no mostraron signo alguno.
Evaluación clínica
Los peces se sometieron a examen clínico, durante la aclimatación y posterior a la inoculación durante todo el periodo experimental, a través de la observación de diferentes patrones de comportamiento, de acuerdo a lo descrito por Francis-Floyd (1988) y Francis-Floyd (1999).
Evaluación hematológica
Una vez los animales experimentales presentaban pérdida total del eje de nado, o a las 24 horas post-infección (para animales que aun no presentaban este signo), fueron tranquilizados con agua fría (4°C) y desensibilizados por sección de la medula espinal a la altura del borde caudal del opérculo. La sangre fue colectada por punción de la vena caudal, a la altura de la inserción de la aleta anal, 1 a 1.5 cm por debajo de la línea lateral, mediante la utilización de una aguja número 28 con heparina sódica como anticoagulante.
El hematocrito se determinó mediante el uso de capilares para microhematocrito, se centrifugó a 12000 rpm por 5 minutos. Así mismo, se evaluó el leucocrito. Adicionalmente, se realizaron extendidos, los cuáles fueron teñidos con Hemacolor® (Merck Chemicals, Germany), para realizar los recuentos diferenciales y determinar porcentaje de linfocitos, neutrófilos, eosinófilos, monocitos y basófilos. Se tuvo en cuenta los cambios en la morfología celular.
Hallazgos de Necropsia
Los animales experimentales se sometieron a examen de necropsia siguiendo los lineamientos descritos por Reimschuessel et al (1988), y Yanong (2003). Para esto, los peces fueron insensibilizados mediante sección de la medula espinal, dorsal a los opérculos. Se tomó el hígado y se determinó el índice hepatosomático mediante la fórmula:
IHS=Peso hígado/peso cuerpo x 100.
Análisis estadístico
Se utilizó un diseño experimental completamente al azar de efecto fijo balanceado mediante ANOVA y MANOVA donde las variables respuesta fueron hematocrito, leucocitos, porcentaje de neutrófilos, linfocitos, monocitos, basófilos, eosinófilos e índice hepatosomático(IHS). El modelo se ajustó de acuerdo a la siguiente ecuación:
Yij = µ + πi + εi (i)
Donde µ es la media, πi el efecto tratamiento, y ej(i) el error experimental.
Se utilizaron las pruebas de contraste de Tukey y análisis canónico de índice ortogonal con un 95 % de confiabilidad. Se validaron los supuestos del modelo y se empleó análisis descriptivo exploratorio unidimensional para hallar: media, desviación y coeficiente de variación. Los datos se transformaron mediante arco seno. Los resultados fueron analizados con el software SAS/STAT ®versión 9.0 para Windows.
RESULTADOS
Las cepas aisladas de A. hydrophila de animales experimentales, mostraron características de b-hemolisis en agar BHI suplementado con sangre de ovejo al 5 % (Fig. 1), mientras la cepas de E. tarda no mostraron esta característica.
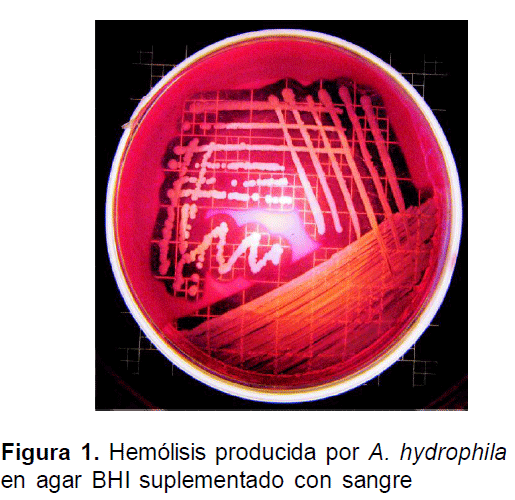
Los animales sometidos al desafío bacteriano presentaron palidez, letargia, boqueo en superficie y en algunos, pérdida total del eje nado a las 11 horas pos-infección para A. hydrophila y de 13 horas pos-infección para la E. tarda. Además, se evidenció aumento de la frecuencia opercular en los grupos experimentales a excepción del grupo control.
En la evaluación hematológica, se observó una coloración más pálida de la sangre y de consistencia más acuosa en animales infectados, siendo de mayor severidad en el grupo inoculado con A. hydrophila. En los frotis sanguíneos, se evidenciaron abundantes formas blásticas de hematíes, así como normoblastos en ambos tratamientos y una disminución marcada en el número de plaquetas comparadas con el grupo control. Los hematíes mostraban hipocromía, cariorrexis y anisocitosis. La hemolisis fue más marcada en el tratamiento con A. hydrophila. Por otra parte, fue posible evidenciar monocitos fagocitando bacterias de E. tarda (Fig. 2), que fueron identificadas en cúmulos rodeados por glóbulos rojos así como de monocitos activos con granulaciones claras evidentes en su citoplasma.
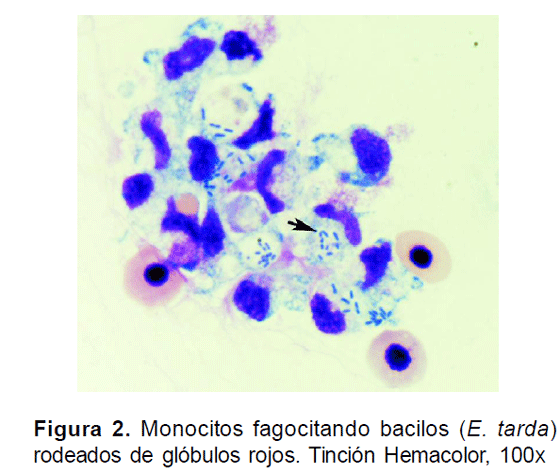
Los valores de hematocrito, leucocrito, recuento diferencial de leucocitos, e índice hepatosomático, se presentan en la Tabla 1. Los valores de hematocrito no presentaron diferencias significativas (p>0.05) entre tratamientos; sin embargo, los valores más bajos fueron hallados para los individuos desafiados con E. tarda. Igualmente, no se hallaron diferencias significativas (p>0.05) en el leucocrito, aunque tuvo tendencia a aumentar en número en animales inoculados con las bacterias. En el recuento diferencial, los linfocitos de los grupos tratamiento de A. hydrophila y E. tarda, tuvieron una disminución muy significativa (p<0.001) con respecto al control, mientras los monocitos tuvieron un porcentaje significativamente más alto en animales inoculados con E. tarda (p<0.05) que en animales control, el recuento de neutrófilos mostró una alta desviación estándar, debido a la gran variabilidad de los conteos de estas células. En la revisión de los frotis, en los animales inoculados con ambas bacterias, se evidenció un grado significativo de hemolisis. No existió diferencia significativa (p>0.05) en el índice hepatosomático de los animales experimentales respecto al control.
En el examen de necropsia de los animales inoculados con A. hydrophila se observó palidez del hígado, patrón moteado en la coloración del mismo, hepatomegalia, consistencia friable y cambios congestivos, tales como arborización (Fig. 3). En el tratamiento con E. tarda, fue posible evidenciar en el bazo de uno de los ejemplares nódulos blanquecinos de distribución multifocal (Fig. 4). De manera general, para los dos desafíos bacterianos se evidenció hemorragia en la base de las aletas, hemorragias en órganos internos, ascitis, un aumento de líquido abdominal de carácter seroso amarillento y líquido sanguinolento en intestino.
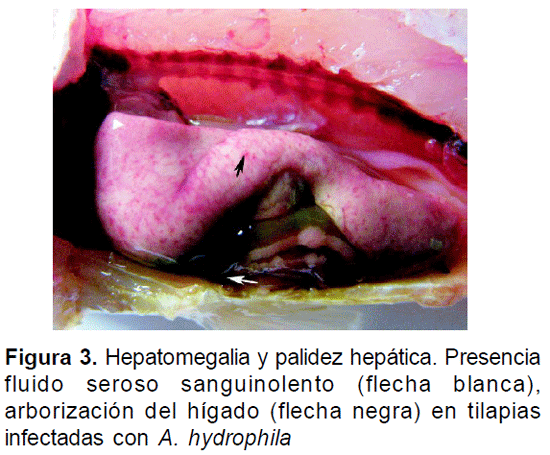
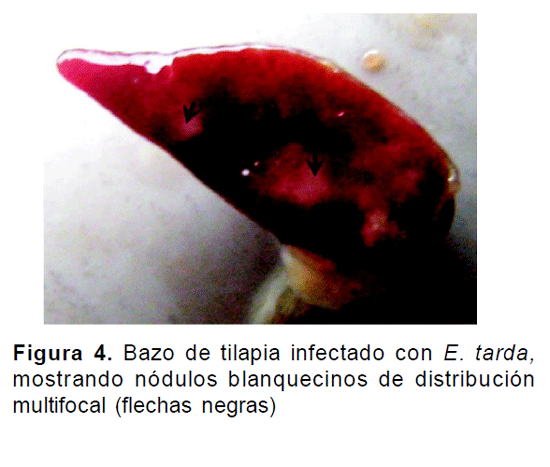
DISCUSIÓN
Tanto la A. hydrophila como la E. tarda han sido asociadas con brotes de septicemia hemorrágica en tilapias de cultivo en el Departamento del Meta y otros regiones productoras piscícolas del país (Iregui et al., 2004; Rondón et al., 2007).
Las cepas aisladas de A. hydrophila de animales experimentales, mostraron características de b-hemolisis. Se ha reportado tanto la secuenciación de genes como la producción de PEC como b-hemolisina y aerolisina, que han presentado actividad hemolítica (Chopra y Houson, 1999; Allan y Stevenson, 1981), que pueden ser responsables del aumento de la fragilidad vascular acompañada de ruptura de pequeños vasos y hemorragias en otros órganos. Además, estudios de mutagénesis sugieren que tanto las hemolisinas como las aerolisinas, actúan induciendo la formación de poros en las membranas de las células afectadas, y el efecto de estas dos parece ser sinérgico (Wang et al., 2003).
Las cepas aisladas de E. tarda no mostraron características hemolíticas en el agar suplementado con sangre, aunque se observó cierto grado de hemolisis en los frotis sanguíneos de peces inoculados con la bacteria. Frente a esto, Chen et al., (1996) caracterizaron y secuenciaron el gen de hemolisinas en cepas de E. tarda, reportando diferentes hemolisinas a las de A. hydrophila. La ausencia de hemolisis observada de la E. tarda en agar, aunque no se demostró en el presente estudio, puede ser debida a la baja o nula sensibilidad de los eritrocitos de ovejo a las hemolisinas de la bacteria, así como a las condiciones de cultivo en el medio (Watson y White, 1979). Adicionalmente Chacón et al (2003) y Santos et al (1999), demostraron que la hemólisis de cepas virulentas depende de la sangre usada en el agar.
Los signos clínicos encontrados en la infección con A. hydrophila, son similares a aquellos evidenciados en tilapias hibridas (Oreochromis sp.) y cachama blanca (Piaractus brachypomus), inoculados experimentalmente tanto con A. hydrophila, como con PEC de A. hydrophila, dentro de los que se encuentran: anorexia, letargia, aumento de la frecuencia opercular y boqueo en superficie, (Rey et al., 2009), signos encontrados en infecciones naturales por Aeromonas mótiles (SAM) en tilapia (Plumb, 1997).
El aumento de la frecuencia opercular en peces desafiados con los patógenos experimentales, puede deberse tanto a la respuesta de estrés, como al desarrollo de anemia que conlleva a la insuficiencia de oxígeno en los tejidos (Mohanty & Sahoo, 2007) y por tanto a un incremento compensatorio de la respiración.
Existen puntos de vista encontrados acerca de la utilidad y pertinencia de la hematología y el leucograma para fines clínicos en peces; principalmente por algunos fenómenos que limitan la confiabilidad de los rangos, dadas las características de maduración intravascular de células sanguíneas. Como lo describen, Groff y Zinkl (1999), existe gran dificultad en el recuento diferencial y total de leucocitos en peces debido a la confusión y la diferenciación de las células inmaduras, como por ejemplo, trombocitos confundidos con linfocitos o monocitos confundidos con hemoblastos. Por esta razón, en el presente trabajo, el recuento total de células blancas no se consideró debido a la alta variabilidad en el conteo, pues aunque se lisan los glóbulos rojos, puede existir error de diferenciación al confundir sus remanentes nucleares con leucocitos. No obstante, se han hecho diferentes trabajos tendientes a establecer rangos "normales" en los valores leucocitarios en peces, tanto tropicales como de aguas templadas. Adicionalmente, el análisis hematológico permite detectar y evaluar algunas condiciones, como son: anemia, desordenes hemostáticos, también como el comportamiento de algunas infecciones, entre otros.
En la evaluación hematológica no se observaron diferencias significativas (p>0.05) en el hematocrito de todos los grupos experimentales; sin embargo, los animales infectados con E. tarda presentaron niveles más bajos y una mayor variabilidad. La reducción en el hematocrito se ha observado en diferentes infecciones bacterianas experimentales en peces, como en tilapias nilóticas infectadas con E. tarda (Benli y Yildiz, 2004), carpas infectadas con Pseudomonas fluorescens (Yildiz, 1998) e infectadas con A. hydrophila (Harikrishnan et al., 2003), en Oreochromis aureus infectados con Corynebacterium sp., (Silveira-Coffigny et al., 2004), Anguilla anguilla infectadas con A. hydrophila (Yildiz et al., 2005); lo cual puede deberse a una alteración severa de la hematopoyesis por las bacterias o sus productos (Yildiz et al., 2005).
Adicionalmente, se ha demostrado que la disminución en el hematocrito y la disminución del conteo total de glóbulos rojos y concentración de hemoglobina en carpas desafiadas con A. hydrophila, está asociada a una anemia hipocrómica microcítica causada por la bacteria y a la destrucción de glóbulos rojos por actividad leucocítica en una anemia eritrocítica con subsecuente eritroblastosis (Harikrishnan et al., 2003). Así mismo, la anemia microcítica por reducción del contenido de hemoglobina, hemoglobina corpuscular media y recuento total de glóbulos rojos, fue también reportada en Oreochromis aureus desafiados con Corynebacterium sp., (Silveira-Coffigny et al., 2004). A pesar de que no existió una diferencia significativa (p>0.05) en el hematocrito entre los grupos experimentales, se observó una mayor presencia de formas blásticas, en los animales desafiados con las bacterias, lo cual puede estar relacionado con la hipocromía y anisocitosis, así como la eritroblastosis y presencia de formas blásticas de células blancas reportados para peces infectados en este estudio.
En el recuento diferencial fue posible diferenciar: trombocitos, monocitos, linfocitos y neutrófilos, de manera poco frecuente se observaron, eosinófilos y basófilos, similar a lo observado en otros trabajos similares en Oreochromis sp., (Ueda et al., 2001; Silveira-Coffigny et al., 2004). También se logró observar hemólisis en células sanguíneas de los peces inoculados con las bacterias.
En los cuadros bacterianos agudos tanto en mamíferos como en peces, la línea celular sanguínea predominante es el neutrófilo, siendo la neutrofilia y la linfopenia características en hemogramas de animales septicémicos. En el presente trabajo se evidenció linfopenia muy significativa y proliferación de monocitos (denominados monocitos/macrófagos). Sin embargo, no se puede confirmar un fenómeno de neutrofilia dada la alta variación y baja confiabilidad de los valores porcentuales hallados en este estudio.
Los extendidos sanguíneos de animales desafiados con A. hydrophila y E. tarda, mostraron un número significativo de eritrocitos inmaduros y anisocitosis de los mismos, también como hipocromía y cariorrexis marcada. Similar a los hallazgos de Silveira-Coffigny et al., (2004) que demostraron que O. aureus infectados con Corynebacterium sp., presentaban eritrocitos inmaduros, deformes y microcistos y eritrocitos con el núcleo segmentado; posiblemente, por un cuadro de anemia hipocrómica microcítica. En estos extendidos, además fue posible evidenciar agregados celulares de interacción de eritrocitos con bacterias y leucocitos, similares a rosetas, las cuales se considera que incrementan la capacidad fagocitaria de los macrófagos, gracias a la polarización de los eritrocitos hacia ellos (Passantino et al., 2002).
En general en animales infectados, existe evidencia de un proceso hemorrágico que conduce a anemia (compatible con la morfología celular descrita) y pérdida de plasma; sin embargo, en el presente estudio no se correlacionó la anemia con los valores de hematocrito y leucocrito, (ya que no se presentaron diferencias significativas esperadas en peces infectados), siendo estos parámetros, tomados aisladamente, insuficientes para apoyar un diagnóstico en procesos septicémicos.
Los hallazgos de necropsia fueron similares a los reportados por Rey et al (2008), en infecciones experimentales: cambios hemodinámicos sistémicos y locales como: congestión, hemorragias, ascitis, permeabilidad vascular incrementada, expulsión de fluido sanguinolento a través del ano, edemas e intususcepción (esta última no se observó en el presente trabajo).
En peces inoculados con E. tarda, se han reportado lesiones de septicemia, como: hiperemia con congestión sanguínea de aletas, con hemorragias petequiales en diferentes partes del cuerpo. Internamente con hiperemia general del peritoneo, hígado moteado y con edemas (Inglis, 1993), lo cual está acorde con lo observado en el presente trabajo. En el bazo de uno de los ejemplares, la observación de nódulos blanquecinos de distribución multifocal fue similar a lo descrito por Rondón et al., (2007) quienes demostraron el desarrollo de lesiones similares en infecciones naturales por E. tarda. De la misma manera, Muroga (1997) ha reportado la presencia de puntos blancos similares a granulomas, que contienen grupos de bacterias con fagocitos alterados y cubiertos por tejido fibroso que se desarrollan en el bazo y riñón en Yellowtail infectados experimentalmente con Pasteurella piscida. A pesar que no se realizó examen histopatológico, existen reportes de la formación de piogranulomas en bazo de tilapia hibrida infectadas naturalmente con E. tarda (Rondón et al., 2007), que pueden estar asociados con la evidencia a la necropsia de dichos nódulos en el presente estudio.
La infección experimental de tilapias con E. tarda y A. hydrophila en el presente estudio, demostró un cuadro clínico compatible con septicemia hemorrágica, dado que existieron signos evidentes y alteraciones de patología clínica, incluyendo: hipocromía, anisocitosis y eritroblastosis, que pueden estar asociados a factores de virulencia, tales como PEC, que se puede esperar tengan un papel importante en las enfermedades como la septicemia hemorrágica de peces, tal como se ha demostrado en estudios de patogénesis en Oreochromis sp., y P. brachypomus inoculados con A. hydrophila y sus PEC mostrando en ambas inoculaciones cambios hemodinámicos sistémicos y locales (Rey et al., 2009). Dentro de estos productos extracelulares se destacan la hemolisina y aerolisina con actividades hemolíticas y citotóxicas con toxicidad letal en peces (Khalil y Mansour, 1997; Allan y Stevenson, 1981), otros factores como GCAT producido por A. hydrophila también pueden contribuir con la lisis eritrocítica, digiriendo sus membranas plasmáticas y reportado en enfermedades de peces como la furunculosis (Chopra y Houston, 1999).
Aun cuando estadísticamente no se hallaron diferencias significativas (p>0.05) en el índice hepatosomático, los animales experimentales sometidos a desafíos bacterianos mostraron un patrón de aumento de tamaño del órgano y existió una tendencia a incrementar el índice hepatosomático de peces infectados con E. tarda. Un incremento marcado de este parámetro se ha evidenciado en lenguados, Paralichthys olivaceus inoculados con E. tarda luego de 72 horas post-inoculación, sin embargo, el aumento del peso del hígado no estaba asociado a cambios degenerativos ni a la respuesta inflamatoria en el tejido, sino a la gran transcripción de genes o síntesis de proteínas estimuladas por la infección con E. tarda. Igualmente, se ha demostrado que el contenido de ADN por unidad de peso del hígado disminuyó en peces expuestos a E. tarda, comparado con animales control, sugiriendo un incremento en el tamaño de cada célula de peces infectados (Miwa y Mano, 2000). Estos autores sugieren que el aumento del tamaño del hígado puede ser resultado directo de la hipertrofia de los hepatocitos, y en algunos casos a una hiperplasia leve de los mismos.
Este trabajo contribuye a la aproximación de modelo en el estudio de la dinámica de enfermedades septicémicas en tilapias de cultivo en Colombia, como un paso inicial en la evaluación sistemática de la fisiopatología y dinámica de las enfermedades bacterianas en peces usando cepas obtenidas de brotes o casos de campo, tomando en cuenta que las cepas de microorganismos presentan diferentes variaciones interespecíficas y que en ocasiones la presencia de una misma bacteria puede remitir a cuadros clínicos diferentes dependiendo de las condiciones del medio así como de la interacción con otras bacterias o microorganismos asociados.
AGRADECIMIENTOS
Los autores agradecen el soporte del proyecto 276-06 IICA-MADR-Unillanos en el aislamiento de las cepas bacterianas, también agradecen el apoyo del Laboratorio de Microbiología de la Universidad de los Llanos, así como a David Pardo Hernández y Miguel Bonilla, por su colaboración durante la fase experimental.
REFERENCIAS
Allan B, Stevenson R. Extracellular virulence factors of Aeromonas hydrophila in fish infection. Can J Microbiol 1981; 27: 1114-1122. [ Links ]
Benli K, Yildiz Y. Blood parameters in Nile tilapia (Oreochromis niloticus L.) spontaneously infected with Edwardsiella tarda. Aqua Res 2004; 35: 1388-1390. [ Links ]
Chacón M, Figueras G, Castro G, Soler L, Guarro J. Distribution of virulence genes in clinical and environmental isolates of Aeromonas spp. Antonie van Leeuwenhoek 2003; 84: 269-278. [ Links ]
Chen J, Lai S, Huang S. Molecular cloning, characterization and sequencing of the hemolysin gene from Edwardsiella tarda. Arch Microbiol 1996; 165: 9-17. [ Links ]
Chopra A, Houston C. Enterotoxins in Aeromonas-associated gastroenteritis. Review. Microbes infect 1999; 1:1129-1137. [ Links ]
FAO. State of world aquaculture. Production: environments, species, quantities and values. Fishery and Aquaculture Department. Corporate Document Repository. Fisher Technical paper Nº500 2006. Roma. 16 p. [ Links ]
Francis-Floyd R. Behavioral diagnosis. Vet Clin North Amer: Small Anim Pract 1988; 16:303-314. [ Links ]
Francis-Floyd R. Clinical Examination of Fish in Private Collections. Vet Clin North Am Exotic Anim Pract 1999; 2:247-264. [ Links ]
Groff JM, Zinkl JG. Hematology and clinical chemistry of cyprinid fish. Common carp and goldfish. Vet Clin North Amer Exot Anim Pract 1999; 2(3):741-776. [ Links ]
Harikrishnan R, Nisha Rani M, Balasundaram C. Hematological and biochemical parameters in common carp, Cyprinus carpio, following herbal treatment for Aeromonas hydrophila infection. Aquaculture 2003; 221: 41-50. [ Links ]
Inglis V, Roberts R, Bromage N. Bacterial diseases of fish. Blackwell Publishing. 1993. 312p. [ Links ]
Iregui C, Hernández E, Jiménez A, Pulido A, Rey AL, Comas J, Peña LC, Rodríguez M. Primer mapa epidemiológico de las lesiones y enfermedades de los peces en Colombia. Universidad Nacional de Colombia - Ministerio de Agricultura. 2004. 100pp. [ Links ]
Jimenez A, Iregui C, Figueroa J. Caracterización y evaluación in vivo e in vitro del lipopolisacarido de Aeromonas hydrophila Acta biol Colomb 2008, (13): 147-162. [ Links ]
Khalil A, Mansour E. Toxicity of crude extracellular products of Aeromonas hydrophila in tilapia, Tilapia nilotica. Lett Appl Microbiol 1997; 25: 269-273. [ Links ]
Kostrzynska M, Dooley J, Shimojo T, Sakata T, Trust T. Antigenic diversity of the S-layer proteins from pathogenic strains of Aeromonas hydrophyla and Aeromona veronii biotype sobria. J Bacteriol 1992; 174 (1): 40-47. [ Links ]
Plumb J. Infectious disease of tilapia En: Costa-Pierce A, Rakocy E. Tilapia Aquaculture in the Americas. World Aquaculture Society. Auburn. 1997. Vol I. 212-228. [ Links ]
Miwa S, Mano N. Infection with Edwardsiella tarda causes hypertrophy of liver cells in the Japanese flounder Paralichthys olivaceus. Dis Aquat Org 2000; 42: 227-231. [ Links ]
Mohanty B, Sahoo P. Edwardsiellosis in fish: a brief review. J Biosci 2007; 32: 1331-1344. [ Links ]
Morohoshi T, Inaba T, Kato N, Kanai K, Ikeda T. Identification of quorum-sensing signal molecules and the LuxRI homologs in fish pathogen Edwardsiella tarda. J Biosci Bioeng 2004; 98(4): 274-281. [ Links ]
Muroga K. Recent advances in infection disease of marine fish with particular reference to the case in Japan. En: Disease in Asian Aquaculture III. Proceedings of the third symposium on disease in asian aquaculture. Thailand. 1997. 21-31. [ Links ]
Murugappan R, Rekha S, Thirumurugan R. Characterization and quantification of siderophores produced by Aeromonas hydrophila isolated from Cyprinus carpio. Pak J Biol Sci 2006; 9(3): 437-440. [ Links ]
Passantino L, Altamura M, Cianciotta A, Patruno R, Tafaro A, Jirillo E, Passantino GF. Fish immunology. I. Binding and engulfment of Candida albicans by erythrocytes of rainbow trout (Salmo gairdneri Richardson). Immunopharmacol Immunotoxicol 2002; 24(4): 665-678. [ Links ]
Reimschuessel R, May E, Bennett R, Lipsky M. Necropsy examination of fish. Vet Clin North Amer Trop Fish Med: Small Anim Pract 1988; 18:427-433. [ Links ]
Rey A, Verjan N, Ferguson H, Iregui C. Pathogenesis of Aeromonas hydrophila strain KJ99 infection and its extracellular products in two species of fish. Vet Record 2009; 164(16):493-499. [ Links ]
Rey A. Caracterización de las enfermedades de tilapia roja (Oreochromis sp) cultivada en el departamento del Tolima. Sistematización de la información. Tesis de Maestría en producción animal [2002]. Se localiza en la Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional de Colombia. [ Links ]
Roberts RJ.2001. Fish Pathology. Third edition. Saunders, London.472pp. [ Links ]
Rondón-Barragán I, Ramírez-Duarte W., Gutiérrez G, Eslava-Mocha P. Edwardsiellosis en tilapia. En: Memorias XIII Jornada de Acuicultura, Instituto de Acuicultura de los Llanos, Universidad de los Llanos, Septiembre 21 de 2007. Pág. 84-88. [ Links ]
Santos J, Gonzalez C, Otero A, García M. Hemolitic activity and siderophore production in different Aeromonas species isolated from fish. Appl Environ Microbiol 1999; 65: 5612-5614. [ Links ]
Silveira-Coffigny R, Prieto-Trujillo A, Ascencio- Valle F. Effects of different stressors in haematological variables in cultured Oreochromis aureus S. Comp Biochem Physiol 2004; 139: 245-250. [ Links ]
Tizard I. Veterinary immunology. 8 edition. Saunders Elsevier. 2009. 600p. [ Links ]
Ueda IK, Egami MI, Sasso WS, Maushima ER. Cytochemical aspects of the peripheral blood cells of Oreochromis (tilapia) niloticus. Linnaeus, 1758 (Cichlidae, Teleostei): Part II. Braz J Vet Res Anim Sci 2001; 38: 273- 277. [ Links ]
Wang G, Clark C, Liu C, Pucknell C, Munro T, Kruk A, Caldeira R, Woodward D, Rodgers F. Detection and Characterization of the hemolysin genes in Aeromonas hydrophila and Aeromonas sobria by multiplex PCR. J Clin Microbiol 2003; 41(3): 1048-1054. [ Links ]
Watson J, White F. Hemolysins of Edwardsiella tarda. Can J comp Med 1979; 43: 78-83. [ Links ]
Yanong RPE. Necropsy techniques for fish. In Practical Gross Necropsy of Exotic Animal Species. Seminars in Avian and Exotic Pet Medicine., Echols S., ed. W.B. Saunders Co., Philadelphia, PA. 2003. pp. 89-105. [ Links ]
Yildiz Y, Bekcan S, Benli K, Akan M. Some blood parameters in the eel (Anguilla anguilla) spontaneously infected with Aeromonas hydrophila. Case study. Israel J Vet Med 2005; 60:91-92 [ Links ]
Yildiz Y. Effects of experimental infection with Pseudomonas fluorescens on different blood parameters in carp (Cyprinus carpio L). J Israeli Aquaculture 1998; 50: 82-85.
[ Links ]
