











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introduction
The lesser capybara (Hydrochoerus isthmius, Goldman, 1912) is a rodent distributed in Colombia, Venezuela and Panama. In Colombia, it is present in the Andes, the Caribbean and the Pacific regions (1). For many years it was classified as a subspecies of Hydrochoerus hydrochaeris. However, it is currently considered as a different species (2, 3).
The habitats of these animals are diverse and they have as a common element a wetland and a grazing area that can be pastures, gallery forests, or shrubs (4). It is a rodent with a digestion similar to ruminants, and can compete with cattle for resources (4, 5). Currently, the species is listed as Data Deficient in view of the absence of information on population and on its extent of occurrence (1).
Its reproductive capacity, high growth rate, hardiness, social behavior and quality of its meat, makes the lesser capybara a promising resource for biomedical innovations and better productive management for protein in captivity (6). However, studies regarding its ecology, production, sustainability and morphophysiology are scarce in its distribution range (3, 4, 6).
One of the least explored knowledge areas for these mammals is the digestive morphophysiology, highlighting some descriptive studies done on different organs of H. hydrochoerus in Brazil (7, 8). The pancreas is a digestive organ located in the abdomen. In mammals, it contains exocrine glands that produce enzymes important to digestion, including trypsin, chymotrypsin, amylase and lipase. The endocrine component of the pancreas consists of islet cells (islets of Langerhans) that create and release important hormones like insulin and glucagon (9).
Based on this, and due to pancreas importance on the absorption, secretion and excretion (10), the aim of this study was to describe histologically and histochemically the pancreas of adult individuals of H. isthmius, including the identification of cellular components belonging to the exocrine and endocrine portion of this digestive organ.
Materials and methods
Study site
The study was conducted in the countryside of Buenavista, located south-east of the department of Cordoba between latitude 8º13'13' N and longitude 75º29'11'' W, at 72 m.a.s.l.
Environmental conditions
The ecosystem corresponds to tropical dry forest. The climate in this region is of unimodal type with two hydrological periods. The annual temperature is 27.8 ° C with average rainfall of 2,181 mm.
Study animals
Three adult individuals of the species Hydrochoerus isthmius. The corpses were opportunistically collected after finding these dead animals as a result of feral dog attacks between June and July of 2015.
Samples collection
The post-mortem examination of the individuals was carried out by making an incision in the midline (alba line) where the viscera were exposed, from the tongue to the anus. The stomach and small intestine (duodenum, jejunum and ileum) were removed from the rest of the organs, exposing the pancreas, and allowing its complete extraction, which allowed to appreciate its external morphological characteristics as well as its location and its ducts.
Sample analysis
Fragments of the pancreas were collected, until obtaining significant samples of each individual. Then, the samples were stored in vials with 10% buffered formalin and Bouin liquid for 24 hours, as tissue fixative, and after that they passed to the dehydration process in five ascending alcohol concentrations starting at 70%. Later, they were initially diaphanized in a 1:1 mixture of 100% alcohol + Xylol and then in two xylol solutions, embedded in paraffin and sectioned to a 4 μm thickness with a hand rotary microtome.
Hematoxylin & Eosin, PAS, Gomori, Grimelius and Masson Fontana modified staining techniques were used to remove the paraffin. Histological and histochemical characteristics were visualized and identified in the different portions of the pancreas (3, 4, 5). The samples were processed in the histopathology laboratory of the University of Cordoba, Colombia and in the laboratory of Veterinary Histology of the Department of Veterinary Medicine in the Federal University of Viçosa, Minas Gerais, Brazil.
Ethical aspects
Samples were collected taking into account the procedures of sampling, management and conservation, as well as the ethical, technical, scientific and administrative standards for animal research contained in Law 84 (National Congress of Colombia, 1989). This study has a scientific research license from the Autonomous Corporation of the Valleys of Sinu and the San Jorge river (CVS) through resolution No. 2-1033 of May 22, 2015.
Data analysis
The data obtained for this study was descriptive, since the identification and characterization of the different cellular components in the pancreas was performed, based on the cellular characteristics observable with the different staining methods proposed.
Results
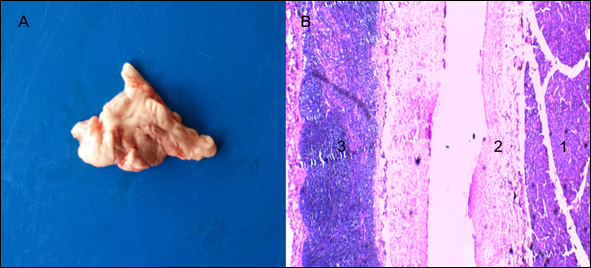
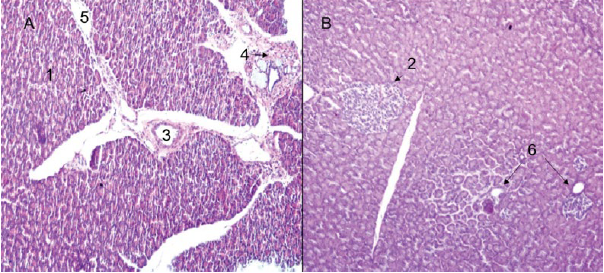
The pancreas (Figure 1A) of H. isthmius presented a duodenal mesenteric pattern. Separated lobes and segments are found; in some cases, largely, by elements of fat and mesenteric stroma. Likewise, lymphoid tissue was observed in the capsule of connective tissue that covers them (Figure 1B).
Source: own work
Figure 1 Hydrochoerus isthmius pancreas. A) Pancreas with mesenteric pattern; B) Panoramic view of the pancreatic parenchyma with presence of lymphoid tissue in the organ capsule (4X) H & E. 1. Exocrine portion. 2. Septum of loose connective tissue. 3. Lymphoid nodule.
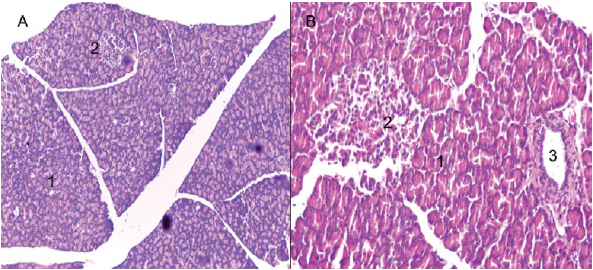
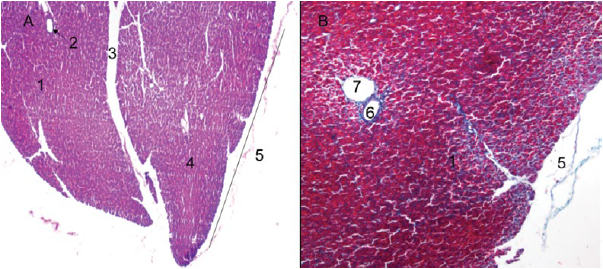
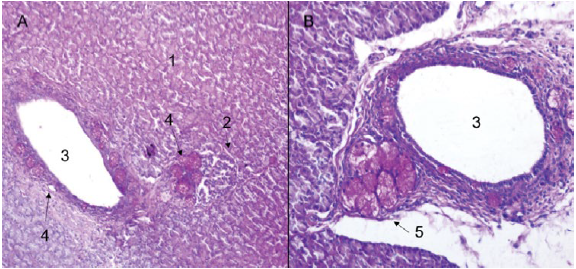
On the other hand, an exocrine and endocrine portion was identified for the pancreas (Figures 2A and 2B). The exocrine portion was described as a composite tubuloacinar gland, which presented clusters of interdigitating branches, that were grouped into lobules constituting the parenchyma, which was covered by a thin layer of loose or areolar connective tissue (Figures 3A and 3B). The parenchyma presented numerous serous acini, few mucous adenomers (mainly related to interlobular ducts and pancreatic islets), intralobular and interlobular ducts (Figures 4A and 4B). Likewise, pancreatic Langerhans islets and the diffuse neuroendocrine system were observed: this one constitutes the endocrine portion of the pancreas (Figure 5A).
Source: own work
Figure 2 H. isthmius pancreas. A) Panoramic view of the pancreatic parenchyma (10X); B) Pancreatic islet (40X), H & E. 1. Exocrine portion. 2. Pancreatic islets. 3. Interlobular excretory duct.
Source: own work
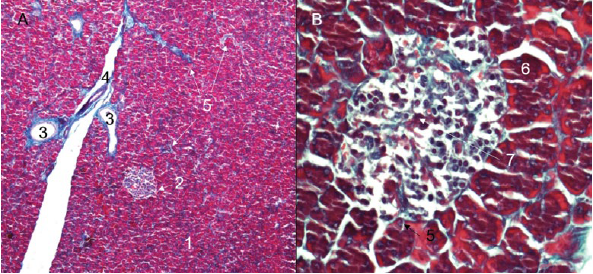
Figure 3 Lesser capybara pancreas. A) Pancreatic lobes (4X), H-E. B) Details of the thin capsule of loose connective tissue covering the pancreas (40X), Gomori trichrome. 1. Exocrine portion. 2. Interlobular excretory duct. 3. Septum of loose connective tissue. 4. Pancreatic lobe. 5. Pancreatic capsule. 6. Artery. 7. Vein.
Source: own work
Figure 4 H. isthmius pancreas. A) Panoramic of an excretory duct, mucous glands and periductal pancreatic islet (10X). B) Detail of interlobular excretory duct with presence of periductal PAS positive mucous glands (40X), PAS. 1. Exocrine portion. 2. Pancreatic islets. 3. Interlobular excretory duct. 4. PAS positive mucous glands. 5. Periductal PAS - Positive mucous glands.
Source: own work
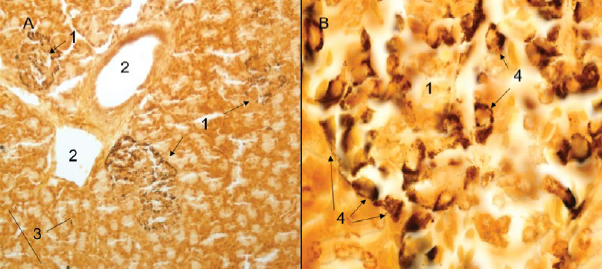
Figure 5 A) Panoramic view of pancreatic parenchyma with presence of periductal pancreatic islets composed of argyrophyll cells and presence of isolated endocrine cells (40X). B) Detail of a pancreatic islet with presence of argyrophyll cells (100X), Grimelius. 1. Pancreatic islet. 2. Interlobular excretory duct. 3. Isolated endocrine cells. 4. Argyrophyll cells.
The pancreas presented connective formations of sustentation, like the capsule that surrounds it. We found a conformation of dispersed, disorganized fibers between the fibroblasts. From there depart the connective septa that are directed towards the interior of the organ, which contain varying amounts of an amorphous substance, blood vessels and lymphatics, as well as adipose tissue, nerves and interlobular pancreatic ducts (Figures 3A and B, and 6A).
Source: own work
Figure 6 A) Interlobular septum of loose connective tissue (10X), H-E. B) Pancreatic parenchyma with interlobular pancreatic islet presence and intralobular or intercalary excretory ducts (10X), PAS. 1. Exocrine portion. 2. Pancreatic islet. 3. Interlobular excretory duct. 4. Septum of loose connective tissue. 5. Vein. 6. Interlobular excretory duct.
From the interlobular septa, delicate connective projections arise, which contain blood vessels into the lobes, wrapping the secretory acini and reaching the pancreatic islets (Figures 7A). Acinar cells are indistinct; and, in the middle of the pancreatic acini, intralobular or intercalar ducts were observed (Figures 6B, 7A). The centroacinar cells were observed: flattened, oval and cubic, depending on the contents of the conduit.
Source: own work
Figure 7 A) Pancreatic parenchyma with presence of interlobular septum of loose connective tissue and interlobular pancreatic islet (10X), Gomori trichrome. B) Detail of the pancreatic islet showing that the cytoplasm of the beta cells adopts a blue-violet (40X) coloration, Gomori trichrome. 1. Exocrine portion. 2. Pancreatic islet. 3. Interlobular excretory duct. 4. Artery. 5. Connective projections. 6. Beta cell. 7. Capillary network.
The endocrine portion of the pancreas was constituted by pancreatic islets of Langerhans (Figures 5B, 7B), which were presented in a variable number and generally in a rounded form. These tend to be unevenly distributed throughout the pancreas, being present in interlobular, periductal and occasionally intralobular sites. However, it was possible to observe a lack of islets sections (Figures 3A). In the middle of the constituent endocrine cells, a dense network of blood capillaries was observed (Figures 7B). It was also possible to observe beta cells, using the Gomori trichromic immunohistochemical staining (Figure 7B).
When staining argyrophyll (Grimelius) and argentafines cells (Masson Fontana modified), a reaction was observed for argyrophyll cells; on the contrary, we found no reaction for argentafine cells.
Discussion
The lesser capybara presents a pancreas with macroscopic characteristics compatible with a mesenteric pattern similar to that of the rabbit, mouse and rat (11). In this model, the lobes and segments are separated, sometimes broadly, by elements of fat and mesenteric stroma; and lymphoid aggregates have been reported in the pancreas of H. hydrochaeris (8, 12). These aggregates have also been observed during this study.
The structural features of the exocrine portion of the pancreas of H. isthmius are similar to those of other domestic mammals (13, 14). Mucous glands were associated with interlobular ducts and in some cases with pancreatic islets. Such glands are not commonly reported in the pancreas; however, some studies report the presence of different cell types producing mucin in the pancreas (15, 16, 17), whose main functions are lubrication, hydration and protection of the epithelium; in the same way, mucin has been shown to play an important role in innate immunity to microbial infections, and has highly immunogenic characteristics, such as blood group antigens (17).
The pancreatic parenchyma is covered by a thin layer of loose or areolar connective tissue, similar to the other domestic mammals (13, 18). Such feature contrasts with the report in H. hydrochaeris, described as a layer of dense connective tissue covering the pancreas (12).
The acinar cells are indistinct; in the middle of the pancreatic acini, intralobular or intercalar ducts were observed. These cells have a basal or centrobasal nucleus and, in the case of H. isthmius, as in other rodents, they can have two nuclei (18, 19). Likewise, centroacinares cells were observed with flattened, oval and cubic forms, depending on the content of the canal: this is compatible with the reported form for the other rodents (20, 21).
The endocrine portion is composed of pancreatic islets that are presented in varying numbers and generally in a rounded form, which is consistent with H. hydrochaeris (12). The pancreatic islets tend to be distributed unevenly throughout the pancreas, which makes possible to observe sections with lack of islets; however, it is not possible to state that there are pancreatic lobes with total islet absent, due to the conformation and three-dimensional location of the same inside the lobe. These are present in interlobular, periductal and occasionally intralobular sites, in agreement with reports for rodents (18, 20).
The pancreas has isolated endocrine cells, which are part of the diffuse neuroendocrine system (12, 18, 20). These cells are argyrophylls or argentaphins, with the exception of immunoreactive insulin producing cells, cholecystokinin and somatostatin, which do not stain with silver (22). When staining the argirophyl (Grimelius) and argentafines cells (modified Masson Fontana), a reaction was observed for argyrophyll cells and, on the other hand, there was no reaction for argentaffin cells (22, 23).
Conclusions
The histological and histochemical techniques used allowed us to identify the exocrine and endocrine portion of the pancreas in this rodent, which has similar characteristics to other mammals. It is suggested to complement this study with some special techniques for the identification of specific endocrine cells, such as Beta, Alpha, Delta, Epsilon, PP or Gamma.

