











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
El cacao se cultiva en el mundo en sitios en los cuales la precipitación varía desde 1200 hasta 4000 mm anuales [1, 2, 3] y la temperatura entre 21 y 32°C con un desempeño óptimo a nivel productivo [4]. Cuando el cultivo de cacao está expuesto a periodos prolongados de sequía se afectan de manera inmediata sus procesos fisiológicos (apertura de estomas, reducción de la fotosíntesis, metabolismo y respiración), como a largo plazo en procesos que indicen en la reducción de su crecimiento y rendimiento [5, 6, 7].
Las áreas con plantaciones de cacao en el departamento del Huila están expuestas a las variaciones climáticas producto del cambio climático [8, 9] debido a períodos prolongados de eventos globales como El Niño y La Niña [10], que aumenta la temperatura entre 1,4-4,8°C [11], generan cambios en los patrones de lluvias y regímenes de precipitación [12], y reducen la productividad [13]. La sensibilidad de las plantas de cacao al déficit hídrico se debe principalmente a características morfo lógicas de sus hojas, al presentar variaciones de area foliar que están relacionadas con la transpiración [14]; los vasos del xilema tienen mayor diámetro comparado con aquellas plantas que se encuentran en un estatus hídrico adecuado, y debido a este estrés las raíces se concentran entre 0,4-0,8 m de profundidad, que limita la absorción de agua en el suelo [15] y pueden aumentar la sensibilidad del cacao a la cavitación en periodos de sequía [16]. Si el aporte hídrico del suelo está por debajo del requerimiento hídrico estimado del árbol (1400-2000 mm/año) se verá afectado el proceso de floración (reducción en la tasa de producción de flores y el cuajado) [11], que influyen en el rendimiento del grano [13].
Actualmente en Colombia el cacao es cultivado en cuatro regiones agroecológicas: departamentos de Santander y Norte de Santander (altitudes entre 300 y 1200 msnm, precipitación entre 1500 y 2500 mm/año, temperatura entre 23 y 28oC), departamentos del eje cafetero, Caldas, Quindío, Risaralda, Antioquia, Santander, Norte del Tolima, Boyacá (altitudes entre 500 y 1200 msnm, precipitación entre 1500 y 2000 mm/ año, temperatura entre 23 y 26oC), Arauca, Meta, Urabá y Bajo Cauca (altitudes entre 0 y 500 msnm, precipitación > 2500 mm/ año, temperatura entre 26 y 30oC), Huila, Cauca, Valle del Cauca, Tolima, Costa Atlántica (altitudes entre 0 y 500 msnm, precipitación < 1500 mm/ año, temperatura entre 26 y 30 oC), pero estas regiones presentan condiciones de precipitación contrastantes (<1200 mm hasta 4000 mm anuales) [17].
Algunas de estas regiones no presentan restricciones en clima y suelo que afecten el rendimiento del grano de manera importante (suelo profundos > 100 cm, bien drenados, temperatura entre 24 y 28°C y precipitación anual entre 1800 y 2600 mm/año que garantizan las necesidades hídricas del cultivo), pero cultivos de cacao ubicados en Valles interandinos secos presentan restricciones en disponibilidad de agua con precipitaciones anuales < 1200 mm que pueden ser no aptas para la producción de cacao [13].
En el departamento del Huila la actividad cacaotera se concentra en 10 municipios, con mayor producción en Campoalegre, Rivera, Algeciras y Hobo. Esta región se caracteriza por condiciones de bosque seco tropical, con precipitaciones que varían entre 668 mm y 1500 mm anuales, una marcada estacionalidad de lluvias de una época seca larga (Julio, Agosto y Septiembre) y seca corta (Enero y Febrero), alta radiación solar mensual (1498,2 micromol m2/s) y brillo solar (6 horas de sol/día) que pueden incidir en el uso de agua por las plantas de cacao, en respuesta a las condiciones ambientales [18 ]
Estas fluctuaciones del clima han motivado a los productores a implementar diferentes arreglos agroforestales con el cultivo de cacao [19, 20], como una estrategia de adaptación a condiciones subóptimas o adversas en los periodos secos que favorezca la producción del grano, advirtiendo que existe un efecto del tipo de árbol sobre las estrategias de adaptación [21].
Los cultivos de cacao asociado con diversas especies de árboles producen condiciones de sombra que permitan un adecuado desarrollo del cultivo y buen desempeño fisiológico de la planta, aprovechando su condición umbrófila (tolerante a la sombra) [22]. Se han implementado sistemas de riego debido a la alta radiación y demanda hídrica, pero por sus altos costos de producción solo se ha implementado en el 0,5% del área cultivada [23, 24].
De acuerdo a esta situación, los sistemas agroforestales se convierten en una estrategia de adaptación a las condiciones hídricas limitantes especialmente por la alta radiación durante las épocas secas para la producción de cacao. El rol de los SAF para el mantenimiento del estatus hídrico en los árboles de cacao radica en la modificación de las condiciones microclimáticas lo cual incide notablemente en la transpiración.
El potencial hídrico es el trabajo necesario para llevar el agua ligada al nivel del potencial de agua pura (MPa = 0) a una misma temperatura y presión atmosférica [25]. Esta variable influye en el crecimiento celular, la fotosíntesis y la productividad de los cultivos, y es un indicador del estado hídrico del cultivo [26]. El monitoreo del flujo de savia (Vs) y del potencial hídrico del xilema (Ѱ MPa) en las especies arbóreas permite estimar la transpiración [27] y entender las respuestas específicas de la planta a la variación de los parámetros ambientales, la disponibilidad de agua y luz [28, 29], para proponer el desarrollo de sistemas de producción estables [30].
Con base en este escenario es necesario desarrollar estudios cuantitativos sobre la respuesta de las plantas de cacao (productividad) al déficit hídrico y maneras de mitigación de los efectos adversos [1, 21]. El objetivo del presente estudio fue determinar la influencia de la cobertura arbórea sobre el estatus hídrico (dinámica del flujo de savia y potencial hídrico) en árboles de cacao Theobroma cacao L. en el valle interandino del norte del Huila.
MÉTODO
La investigación se realizó en el municipio de Campoalegre, Huila, (Colombia) en una finca ubicada a 2° 40' 30,2" N y 75° 21' 24,6" W, 533 msnm, temperatura promedio anual de 25,6°C, humedad relativa promedio anual de 68%, brillo solar de 1679 horas/año y promedio de 6 horas de sol día; que corresponde a una zona de bosque seco tropical en la región fisiográfica del valle superior del río Magdalena. El régimen de lluvias es bimodal, con una precipitación media anual 1370 mm con dos picos de lluvia: marzo-mayo y octubre a noviembre.
Sistemas de producción cacaoteros
Se utilizaron cultivos en estado productivo de cacao clonados con el genotipo CCN51 que tenían un marco de siembra 3 x 3 m, edad promedio de plantación de 8 años, caracterizados por los siguientes sistemas de producción i. a libre exposición solar (SLE) y ii. bajo sistemas agroforestales (SAF). Para los cultivos bajo SAF se experimentaron dos opciones: i. con dominio de frutales (Annona muricata, Morinda citrifolia, Psidium guajava), niveles de radiación transmitida baja (BPAR), dosel del 66% de apertura, un índice de área foliar promedio de 2,3 y una densidad de árboles frutales de 103 árboles en 0,01 ha ii. con dominio de especies vegetales maderables (Ochroma pyramidale, Cordia alliodora, Cecropia peltata y Guarea guidonia), radiación transmitida media (MPAR), apertura de dosel de 22%, un índice de área foliar promedio de 3,1 y una densidad de árboles maderables de 12 árboles en 0,01 ha.
Las condiciones de clima y suelo fueron homogéneas para las parcelas de los sistemas de producción evaluados ya que se encontraban en la misma finca, una seguida de la otra.
Medición del flujo de savia y potencial hídrico
El flujo de savia (Vs) fue determinado utilizando sensores tipo SFM1 (ICT International, Armidale, Australia), instalados en el tronco principal de tres árboles de cacao en cada parcela (n=9), a 15 cm de altura del suelo. Cada sensor SFM1, está conformado por un sistema de recolección de datos (CR10X datalogger + multiplexor AM416, Campbell Scientific, Logan, UT, USA), programado para almacenar los datos cada diez minutos y posteriormente la información semanalmente en el Software ICT HRM (Versión: 2.0.5.4). Los sensores SFM1 se fundamentan en el método de relación de calor (Heat Ratio Method, HRM) [31]. En el tronco de cada árbol de cacao se instalaron dos agujas y una sonda de calor integrada a un microprocesador para determinar la velocidad del flujo de savia. Cada aguja está compuesta por dos termocuplas las cuales reciben el pulso de calor que incrementa la temperatura en el xilema conductor y es proporcional a la velocidad del pulso de calor (Vs/cm/h) [31, 32].
La Vs, se estimó utilizando la ecuación 1.
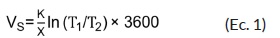
Donde:
K. = Difusividad térmica en el área del xilema conductor (se asumió un valor por defecto de 0,0025 cm2/s).
x. = Distancia entre la sonda de calor y cada una de las agujas (cm).
T. = Aumento de la temperatura (°C) inicial con respecto a la aguja ubicada en la parte inferior (T1) y superior ( T2)
Para determinar el potencial hídrico del xilema (Ѱ, Mpa) se utilizaron psicrómetros de tallo PSY (ICT International, Armidale, Australia) instalados a la misma altura del suelo del tronco principal del árbol donde se ubicaron los sensores tipo SFM1. Para el registro y almacenamiento de datos se instaló a cada psicrómetro un “datalogger” Em-50 (Decagon Devices, Pullman, Washington, USA), con frecuencia de registró de diez minutos. Los datos registrados fueron descargados cada semana en el Software PSY1 ICT (Versión: 2.0.5.9). El Ѱ Mpa se calculó según lo propuesto por [33].
Análisis estadístico
Los datos obtenidos en dos temporadas contrastantes de precipitación (época de máxima precipitación: del 17 de marzo al 13 de Abril del 2017 y época de mínima precipitación: del 21 de Septiembre al 11 de Octubre de 2017), de flujo de savia y potencial hídrico fueron analizadas ajustando modelos lineales mixtos (MLM) para estudiar el efecto de los factores fijos (i. sistema de producción, ii. Época y iii. Hora). Se incluyeron como efectos aleatorios el día y las repeticiones de los árboles de cacao. Los supuestos de normalidad y homogeneidad de la varianza se evaluaron mediante un análisis residual exploratorio. Las diferencias entre los sistemas de producción se analizaron con la prueba post-hoc de LSD de Fisher con una significación de α = 0,05. El MLM se realizó utilizando la función lme en el paquete nlme y las gráficas fueron realiza utilizando el paquete “ggplot2” el software de lenguaje R, versión 3.4.4 (R Development Core Team, 2020), por la interfaz en InfoStat [34].
RESULTADOS
Potencial hídrico
Al comparar el gradiente del potencial hídrico del xilema conductor (Ѱ), este fue mayor en el cultivo a libre exposición solar (SLE) en las dos épocas de monitoreo pero se acentuó más en la época seca cuando muestra valores de -3,08±0,1 Mpa. En los sistemas agroforestales (SAF) con maderables con medio nivel de radiación transmitida (MPAR) el valor fue intermedio (-2,29±0,1 Mpa), seguido del SAF con frutales con nivel de radiación transmitida baja (BPAR) que registró el menor valor de potencial hídrico durante la misma época (-0,75±0,1 Mpa).
Flujo de savia
En el flujo de savia (Vs) hay un comportamiento diferente en las dos épocas climáticas en el caso de SAF con frutales (BPAR). En la época seca se alcanza el valor extremo entre las 10 y las 15 horas. En el SAF con maderables (MPAR) las curvas muestran el mismo patrón y en algunos momentos del día se presentan diferencias significativas (P<0,005). En el cultivo a libre exposición solar (SLE) el comportamiento es muy parecido en las dos épocas a lo largo del día.
Se evidencia en la Figura 1, como el flujo de savia en el SAF con frutales (BPAR) durante la época de mínima precipitación tiene el mayor valor (0,5 L h-1) a las 12 horas, seguido del SAF con maderables (MPAR) (0,38 L h-1) entre las 10 y las 18 horas, frente al cultivo a libre exposición solar (SLE) con un menor valor (0,35 L h-1) entre las 12 horas.
Flujo de savia y potencial hídrico
Los valores del potencial hídrico (Ѱ) se correlacionan de manera negativa con el flujo de savia (Vs r= -0,65 P<0,001). En la Figura 1, se observa un comportamiento opuesto entre el potencial hídrico (Ѱ) y el flujo de savia (Vs), entre los sistemas de producción, durante la época seca. En las dos variables (Ѱ y Vs) el SAF con frutales (BPAR) muestra el menor potencial hídrico y mayor flujo de savia, seguido del SAF con maderables (MPAR) y con los menores valores en el sistema de producción a libre exposición solar (SLE).
En la época de máxima precipitación el mayor flujo de savia se observa en el sistema de producción a libre exposición solar (SLE) y el SAF con maderables (MPAR) (0,36 ±0,1 L h-1), con un menor valor en el SAF con frutales (BPAR) (0,25 L h-1), pero el potencial hídrico no tuvo el mismo comportamiento, siendo estadísticamente menor en el SAF (BPAR) (-0,3±0,1 Mpa), seguido del SAF (MPAR) (-0,6±0,1 Mpa) y el máximo valor en el cultivo a libre exposición solar (SLE) (-1,5 ±0,1 Mpa) (P<0,01 Figura 1).
Los resultados descritos sobre el comportamiento del flujo de savia (Vs) durante la presente investigación, son similares a los descritos por otros estudios [18, 22]. Las condiciones que probablemente pueden incidir sobre el comportamiento del estatus hídrico puede ser la variabilidad climática o la variación interanual [35, 36, 37] que ocasiona una alta demanda evaporativa [38] causando el cierre estomático. Estas situaciones han sido descritas en otros estudios específicamente para el cultivo del cacao [18, 38] en los cuales mencionan que Vs depende de los incrementos de la radiación y del déficit de vapor de presión. Esto indica el efecto del dosel de sombra sobre procesos de apertura y cierre estomático lo cual se refleja en la dinámica de Vs [38].
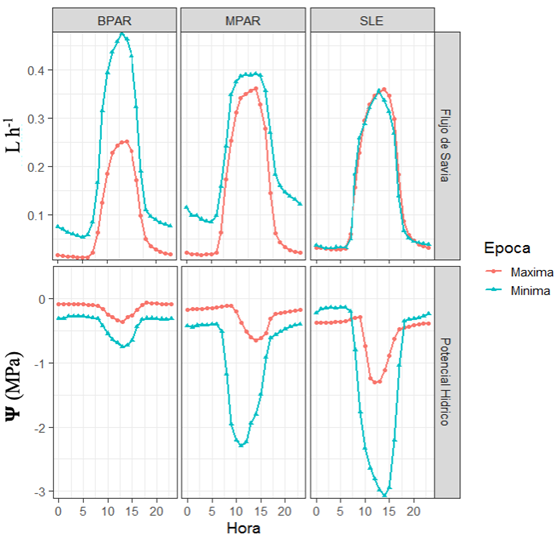
Figura 1 Comportamiento del flujo de savia (Vs) y del potencial hídrico (Ѱ, Mpa) en árboles de cacao en SLE y SAF (BPAR con frutales y MPAR con maderables), en máxima y mínima precipitación.
Para el caso del comportamiento del Ѱ que se encontró durante la investigación, se puede relacionar con una respuesta fisiológica de la planta a la disponibilidad de agua en el suelo y su distribución en la planta [22] que es limitada por variables atmosféricas [15, 18], específicamente para la época de mínima precipitación. La disminución del agua en la zona de raíces implica ejercer una mayor presión para movilizar el agua desde gradientes diferenciales de presión en el complejo suelo-planta-atmósfera [39, 40, 41, 42].La transpiración excesiva se controla por los estomas; se evitan daños de embolia [43, 44], aunque se reducen la fotosíntesis y el rendimiento (producción) de la planta, causados por la sequía [45]. Se podría considerar que a partir de nuestros resultados probablemente las plantas de cacao tienen un compartimiento isohídrico debido a que cierran sus estomas y se presenta una caída en el potencial hídrico del suelo, es importante advertir que en nuestro estudio el genotipo evaluado es el CCN 51, ya que se ha mencionado que el genotipo puede incidir [46].
Bajo estas condiciones las plantas de cacao en el sistema a libre exposición solar (SLE) durante la época de mínima precipitación, pudieron presentar un estrés hídrico severo (potencial hídrico < -1,76 Mpa) [47], por la sensibilidad de la especie al déficit hídrico [8]. El mecanismo de las plantas ante el déficit hídrico en los tres sistemas de producción analizados reflejo la pérdida de agua de los tejidos del tallo durante la mañana y una recuperación nocturna, exhibiendo una fase opuesta al flujo de savia, estos cambios están estrechamente relacionados con factores climáticos [48, 43, 49, 50, 41] en árboles perennes en el trópico [39].
Los valores de potencial hídrico (-0,4 hasta -0,75 Mpa) encontrados en el SAF con frutales (BPAR) durante las dos épocas evaluadas fueron menores a 1 MPa que fisiológicamente indican un estrés hídrico moderado [47]. Esta acción se relaciona con la influencia del dosel sobre el microclima que permite reducir la evapotranspiración comparada con la situación a libre exposición solar [22]. Los resultados hallados comprueban que la cobertura arbórea de los árboles asociados con cacao en Sistemas Agroforestales (SAF), reduce el estrés hídrico, favorece el balance hídrico del cultivo [52], el flujo de savia y en consecuencia mejorar los procesos fotosintéticos y el rendimiento del cultivo [8, 9] en épocas de mínima precipitación.
CONCLUSIONES
La tendencia del flujo de savia (Vs) durante el día en los periodos contrastantes de precipitación siguió un patrón similar tanto BPAR y MPAR, comparado con SLE. Al comparar las épocas se evidencia que el Vs fue mayor en la época seca (mínima precipitación) con una magnitud de 0,5 L h-1 bajo el SAF con frutales el cual tenía un bajo nivel de radiación transmitida (BPAR). Sin embargo en el sistema a libre exposición solar (SLE) el comportamiento del Vs no fue estadísticamente diferente.
El potencial hídrico (Ѱ) mostró los valores más altos (-0,75, -2,29 y -3,08 MPa) durante la época seca en los tres arreglos, pero la mayor magnitud se presentó bajo el sistemas a libre exposición solar (SLE), seguida por la condición SAF maderables con radiación transmitida media (MPAR) y por los SAF con frutales con nivel de radiación transmitida baja (BPAR), donde las variaciones entre las épocas climáticas no son tan extremas.
Los resultados evidencian un efecto positivo de los SAF sobre el estatus hídrico en el cultivo de cacao durante las dos épocas evaluadas, porque mejoran las condiciones microclimáticas que favorecen el balance hídrico del cultivo y reducen el estrés hídrico para que no se limite el flujo de savia, la fotosíntesis y por ende el rendimiento del cultivo. Además de estas los servicio ecosistémicos directos, que convierten los SAF en una estrategia adecuada de adaptación al cambio climático por enriquecer y mantener la biodiversidad asociada a las áreas con plantaciones de cacao.

