











 (pdf)
(pdf)




 SciELO
SciELO  Google
Google  SciELO
SciELO  Google
Google 
 Permalink
PermalinkSuma Psicológica
ISSN 0121-4381
EFECTOS EN LA DISTRIBUCIÓN TEMPORAL DE LA RESPUESTA AS0CIADOS A LA VARIACIÓN EN LA PROBABILIDAD DE ENTREGA DE AGUA EN PROGRAMAS SEÑALADOS: ANÁLISIS DEL TIPO DE CONTACTO Y AJUSTE A RELACIONES DE CONTINGENCIA TEMPORAL
EFFECTS ON TEMPORAL DISTRIBUTION OF RESPONSE ASSOCIATED WITH THE VARIATION IN THE PROBABILITY OF WATER DELIVERTY USING STIMULI-CORRELATED TEMPORAL SCHEDULES: ANALYSIS OF THE TYPE OF CONTACT AND ADJUSTMENT TO TEMPORAL CONTINGENCY RELATIONS
Universidad de Guadalajara, México
La correspondencia relacionada con este artículo debe ser dirigida a: Carlos Torres, Centro de Estudios e Investigaciones en Comportamiento, Francisco de Quevedo #180 (Arcos Vallarta), Guadalajara, Jalisco, México. 44100, correo electrónico: jtorres@cencar.udg.mx
Received: Marzo 9 2011 Accepted: Septiembre 29 2011
Resumen
Se evaluaron los efectos de manipular los valores de P en la distribución temporal del responder bajo programas temporales. Se utilizaron 6 ratas albinas experimen-talmente ingenuas las cuales se dividieron en dos grupos. Cada uno de los grupos se expuso a un programa temporal de distinta longitud. El Grupo 1 T= 30 s., el Grupo 2 T=120". Cada grupo se expuso a cuatro fases distintas. El parámetro P se manipuló entre fases en los valores de 1.0, 0.5 y 0.1. Durante las fases experimentales se entregó agua de manera contingente al responder de las ratas de acuerdo al valor de P. En las primeras tres fases se señalizaron de manera diferencial los subintervalos de ∂t y Δt. Durante la fase cuatro (redeterminación de P a 1.0), la señalización se eliminó en ambos subintervalos. Posterior a las fases experimentales las ratas se sometieron a sesiones de sondeo en las cuales la entrega de agua fue independiente al responder. En la medida que los programas temporales prescriben contingencias temporales respecto de las ocurrencias de respuestas y eventos de estímulo, los resultados se analizan en términos de la modulación de las propiedades temporales de dichos eventos. En este sentido, se evidenciaron diferencias en la distribución temporal de las respuestas efectivas e inefectivas bajo condiciones de señalización y no señalización. Se analizan los resultados en términos del tipo de ajuste a las contingencias de ocurrencia característica de los programas definidos temporalmente, además se sugiere que los programas definidos temporalmente representan una herramienta metodológica pertinente para el análisis de las funciones contextual y suplementaria sugeridas por Ribes y López (1985).
Palabras clave: Programas temporales, distribución temporal del responder, ajuste conductual.
Abstract
One study was conducted to evaluate the effects of manipulating the values of P on performance under temporal defined schedules. It was used 6 experimentally naive albino rats that were divided into two groups. Each group was exposed to a temporal defined schedule with different cycle length. Group 1 was T = 30 s. and Group 2 was T = 120 s. Each group was exposed to four distinct experimental phases. The parameter P was manipulated between phases with the values of 1.0, 0.5 and 0.1. The first three phases the subintervals ∂t and Δt were signaled differentially. During the phase fourth (redetermination), signaling was eliminated in both subintervals. The results suggest that the T-cycle length determines the response patterns. In addition, differences were observed in temporal properties of effective and ineffective responses under signaling and no signaling conditions. The results were analyzed in terms of type of the behavioral adjustment to occurring contingencies featured into temporal defined schedules. We suggest that this procedure represent a relevant methodological tool for analyzing the contextual and supplementary functions suggested by Ribes and Lopez (1985).
Key words: Temporal schedules, temporal distribution of response, behavioral adjustment.
Los programas temporales propuestos por Schoenfeld, Cumming & Hearst (1956) se desarrollaron como una estrategia metodológica alternativa a los programas de reforzamiento propuestos por Ferster & Skinner (1957). En este tipo de programas, el análisis de la fuerza de la conducta se identificó a partir de estructurar la operación de reforzamiento con base en criterios temporales derivados de la manipulación paramétrica de: a) la longitud de un ciclo de tiempo repetitivo ( T ); b) la disponibilidad de reforzamiento dentro del ciclo (T o t testada ); y, c) la probabilidad de reforzamiento entre ciclos (P). El análisis experimental de estos parámetros permitió reproducir las ejecuciones características de los programas de intervalo y razón sin la prescripción explícita de contingencias diferenciales de tiempo o número (Schoenfeld & Cole, 1972).
A partir de los elementos definitorios del sistema T se pudo identificar una de las características más importantes asociadas a estos programas: la posibilidad de poder prescribir condiciones nacesarias y suficientes para la ocurrencia de eventos de estímulo en función de criterios exclusivamente temporales (e.g. ocurrencias no dependientes de la actividad del organismo), o a partir de criterios de oportunidad de respuesta asociados a la producción de eventos de estímulo en períodos específicos (e.g. ocurrencias dependientes de la actividad del organismo). Así mismo, en la medida que las condiciones de producción supusieron necesidad y suficiencia a partir de una ocurrencia de respuesta, la posibilidad de que un evento de estímulo no ocurriera como consecuencia de un desempeño no "adecuado" por parte del organismo (ocurrencias de respuestas no asociadas y/o competitivas respecto de las prescritas en la contingencia), podría dar cuenta del ajuste o no a las relaciones de contingencia establecidas en el programa.
Estas condiciones permitieron, bajo una perspectiva paramétrica, caracterizar un dominio experimental basado en criterios exclusivamente temporales para la ocurrencia de eventos de estímulo y respuesta, en el que se pudieron identificar como casos abstraídos de valores particulares de variables continuas a una diversidad de fenómenos estudiados empíricamente dentro del análisis experimental de la conducta (e.g. control del estímulo, evitación, superstición, etc.), y que han sido considerados en la literatura tradicional operante como fenómenos desvinculados entre sí (Schoenfeld & Cole, 1972; Cabrer, Daza & Ribes, 1975; Torres, 2009).
La manipulación de la disponibilidad de ocurrencia de estimulo ha sido asociada principalmente a los parámetros de T y P . El primero, se relaciona a la proporción del ciclo T en el que pueden ocurrir los eventos de estímulo. Esta proporción se obtiene al dividir el ciclo en dos subintervalos: a) el subintervalo tD, asociado a la ocurrencia de eventos de estímulo y b) el subintervalo tD en el que tradicionalmente no se programan contingencias asociadas a la presentación de eventos de estímulo. El parámetro P estipula la proporción de ciclos con disponibilidad para la ocurrencias de estímulos. Ambos parámetros son independientes entre sí y sus valores pueden varíar entre 0.0 y 1.0.
Con el fin de evaluar las interacciones entre estos parámetros, Torres, Villamil y Calleros (2009) desarrollaron un estudio en los que se manipuló entre fases el valor de P (1.0, 0.5, 0.3 y 0.1) e intra fases el valor de T (1.0, 0.5, 0.3 y 0.1) empleando programas temporales de distintas longitudes de ciclo T (30, 120 y 360 s.). Los resultados de este estudio sugirieron a los autores que los efectos de la manipulación de los parámetros fueron dependientes de la duración absoluta del ciclo T. La disminución en los valores de P mostró una función ascendente-descendente de la frecuencia de respuesta en programas temporales con T=30 s. Mientras que en el resto de los grupos (T=120 y 360 s.), siempre se observó una función decreciente de la frecuencia de respuesta conforme disminuyó el valor de P. Por su parte, la`T no mostró un efecto sistemático en la frecuencia de respuesta, aunque se pudo observar una disminución en el porcentaje de entregas de agua obtenidas conforme disminuyó el valor de este parámetro.
Los resultados de este estudio confirmaron los hallazgos de estudios previos en el área: a) frecuencias de respuesta relativamente más altas durante el subciclo tD respecto del subintervalo tD, sobre todo bajo condiciones de señalización diferencial, lo que implicaría una falta de control de estímulo (Skinner, 1938; Weissman, 1961, 1963); b) variación en la frecuencia de respuesta entre sesiones asociado en ocasiones con un porcentaje de pérdida en la entrega de agua disponible elevado y, d) el porcentaje de entrega de agua obtenida no se correlaciona de manera directa con la tasa o frecuencia de respuesta (Ribes & Torres, 1996; Ribes & Torres, 1997; Ribes, Torres & Piña, 1999; Ribes, Torres & Mayoral, 2000; 2002; Ribes, Mayoral, Torres & Ibañez, 2000).
Además, en el estudio de Torres, Villamil y Calleros (2009) en el que no se correlacionaron estímulos diferenciales con los subciclos, las frecuencias de respuesta obtenidas fueron menores a las obtenidas en programas temporales que emplean señales diferenciales asociadas a los subciclos ∂t y Δt (Ribes & Torres, 1996; Ribes, Torres y Piña, 1999), lo que parece apoyar el planteamiento de que los estímulos correlacionados adquieren funciones de "preparación" o priming auspiciando el respoder dentro de los ciclos T (Ribes & Torres, 1996; Ribes, Torres & Mayoral, 2002; Torres, Mayoral & Ribes, 2008).
De acuerdo con Ribes, Zepeda, Arenas y Mayoral (2009), la consistencia de los resultados obtenidos bajo los programas temporales pareciera sugerir que el tipo contingencias que se estructuran en estos procedimientos propician un desempeño variable en función de que:
La condición de disponibilidad limitada integra un elemento de variabilidad que es dependiente de la ocurrencia o no de la respuesta, propiciando cambios en los intervalos entre estímulos producidos que de facto son distintos a los programados. De aquí se desprende que la distribución de la respuesta dentro de cada ciclo T determina la producción o no de eventos de estímulo, lo que implicaría que distribuciones locales distintas de una misma frecuencia de respuesta estarían asociadas a diferentes porcentajes de entregas de agua.
Por otro lado, las variaciones de los parámetros de P y T tienen un efecto modulador de la disponibilidad temporal de estímulos intra y entre ciclos T. Bajo condiciones de producción contingente, la manipulación de estos parámetros pudiera incidir en la distribución temporal de la respuesta en términos de su coincidencia de ocurrencia con el tiempo mínimo especificado por ambos parámetros. Bajo condiciones de entrega no contingente, la distribución de estímulos es constante o con una variación restringida prescrita por el programa.
A partir de esto, resulta razonable suponer que la variabilidad observada entre sesiones y entre sujetos, en términos de la frecuencia de respuesta, pudiera deberse a la variabilidad temporal en la entrega del agua debido a la interacción de la aleatoriedad que imponen los siguientes factores: a) la probabilidad de reforzamiento, b) las condiciones de señalización, c) la ausencia de respuesta en ciclos en los que se encuentra disponible la entrega de agua.
Con el fin de analizar los supuestos anteriores, se propone el presente estudio para evaluar los efectos de variar la probabilidad de reforzamiento (P) en la distribución temporal de la respuesta comparando el desempeño en programas temporales contingentes y no contingentes de distintas longitudes bajo condiciones diferenciales de señalización.
Método
Sujetos
Se utilizaron 6 ratas albinas macho (cepa Wistar) de seis meses de edad, experimentalmente ingenuas y cuyo peso ad libitum osciló entre los 250 y 300 gramos. Las ratas permanecieron alojadas en jaulas plásticas individuales, y tenían un periodo de 30 minutos de acceso diario a agua purifcada después de cada sesión experimental. Las ratas fueron privadas de agua 22.5 horas diarias antes del inicio del experimento y tuvieron acceso libre a comida. Las sesiones experimentales se llevaron a cabo diariamente de 12:00 a 14:00 horas, seis días de la semana durante cuatro meses.
Aparatos
Se utilizaron seis cámaras de condicionamiento operante de las cuales dos de ellas fueron marca MED modelo ENV-007 colocadas dentro de cajas de aislamiento acústico marca MED, modelo ENV-014M, con extractores de aire modelo ENV-2212SM y cuatro fueron de construcción propia (30 cm de largo por 25.5 cm de ancho y 32 cm de altura) las cuales se encontraban dentro de cubículos sonoamortiguadores con extractores de aire de 115 w. Las cámaras tenían un dispensador de agua de (0.01 cc) en la parte central inferior de la pared operativa, así como una palanca al lado derecho del dispensador de agua. Esta palanca se ubicó a 2.8 cm de la pared derecha y a 2.5 cm encima del piso de rejilla. La palanca requería de 0.13 N de fuerza para operar. Se utilizaron tres focos de 28 v. de los cuales dos (luz verde y luz roja) se usaron para señalar los subintervalos ∂t y Δt, respectivamente, durante las primeras tres fases experimentales. Un tercer foco, cuya iluminación era blanca se colocó dentro del dispensador de agua que se encendía durante los 3s. del periodo de disponibilidad de agua.
Procedimiento
Se entrenó a las ratas a presionar la palanca mediante un procedimiento de automoldeamiento que concluyó cuando cada rata obtuvo 100 entregas de agua contingentes a la respuesta a la panca, durante 3 sesiones consecutivas. Las ratas se dividieron en dos grupos de cuatro ratas cada uno. Ambos grupos fueron expuestos a un programa temporal de estímulos (Schoenfeld & Cole, 1972), que consistió en ciclos T continuos, repetitivos. El Grupo 1 se expuso a un programa cuya duración del ciclo T fue de 30 segundos, mientras que el Grupo 2 se expuso a un programa cuya duración del ciclo T fue de 120 segundos. Para ambos grupos, el parámetro T se mantuvo constante en 0.5 (ver Tabla 1).
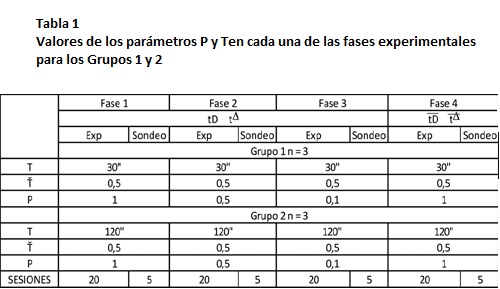
Diseño Experimental
Ambos grupos se sometieron a cuatro fases experimentales. Durante las primeras tres fases, los valores de probabilidad (P) se variaron de manera descendente entre fases (1.0, 0.5, 0.1). En la última fase (fase 4) se redeterminó el valor de (p) a 1.0. Cada fase experimental consistió de 20 sesiones, las cuales tuvieron una duración de 60 minutos cada una. Después de cada fase experimental las ratas fueron sometidas a periodos de sondeo constituidos por 5 sesiones. En las primeras tres fases experimentales, el subciclo tD fue correlacionado con una luz verde mientras que el subciclo tD se correlacionó con una luz roja. Cada ciclo T inició con el subciclo tD y terminó con el subciclo tD, repitiéndose en el mismo orden de secuencia hasta concluir con la cantidad de ciclos correspondientes para cada condición experimental. Durante el subciclo tD, la primer respuesta producía la entrega de agua, si las ratas no respondían en dicho subciclo, se perdía la entrega disponible para ese subciclo. Durante el subciclo tD, ninguna de las respuestas a la palanca tuvieron contingencias programadas. Durante las sesiones de sondeo, las condiciones experimentales eran las mismas que en las sesiones de las fases experimentales, con la diferencia de que la entrega de agua se presentó de manera no contingente a la respuesta y dicha entrega se daba siempre al final del subciclo. Durante las fases experimentales, la entrega de agua se correlacionó con la terminación de la luz verde que estuvo correlacionada con el subciclo tD. En caso de no haber respuesta en el subciclo tD la luz verde se apagaba al finalizar el subciclo y se encendía la luz roja correspondiente al subciclo tD. En las fases de sondeo, la luz verde se apagaba al concluir el subciclo. En los subciclos tD tanto en fases experimentales y de sondeo, la luz roja permanecía encendida durante el tiempo correspondiente al subciclo. En la última fase, de redeterminación del valor de (P), los subciclos tD y tΔ no estaban correlacionados con ninguna señal.
Resultados
Dentro del presente análisis de resultados, se describe la ejecución de las ratas en términos tanto de la tasa de respuesta como de las propiedades temporales de las ocurrencias que estructuran las relaciones de contingencia que se prescriben en los programas de estímulo (Schoenfeld & Cole, 1971). En lo particular, las propiedades temporales de la respuesta se analizaron a partir de distinguir dentro del patrón de ejecución los siguientes indicadores de ajuste:
a) Las respuestas efectivas (REF), identificadas como aquellas respuestas que cumplieron el criterio prescrito por el programa para la producción de la entrega de agua (EAP).
b) Las respuestas inefectivas (RIF), identificadas como las ocurrencias de respuestas que no tuvieron consecuencias programadas dentro de las contingencias prescritas.
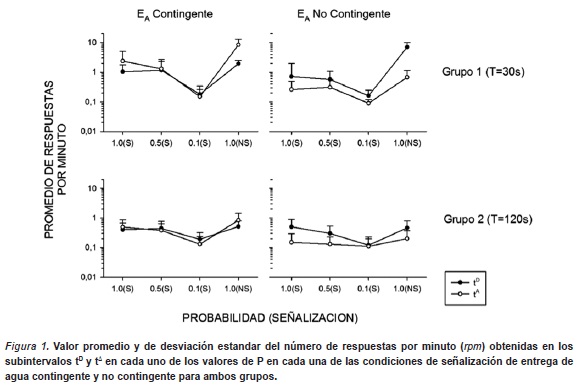
La Figura 1 muestra el valor promedio y de desviación estandar del número de respuestas por minuto (rpm) obtenidas en los subinterva-los tD y tD en cada uno de los valores de P en cada una de las condiciones de señalización de entrega de agua contingente y no contingente para ambos grupos. En el Grupo 1 (T=30s), se observó un decremento gradual de 3.0 rpm a 0.2 rpm asociado a la disminución de P. Este efecto se evidenció principalmente sobre las tasas de respuesas asociadas al subintervalo tD. Durante la condición de redeterminación (P=1.0, sin señal) hubo un incremento en la tasa de respuesta llegando a valores cercanos a 10 rpm. Además, los datos muestran que ante el valor de P=1.0, se observó una mayor tasa de respuesta durante el subintervalo tD que en tD. Por su parte, en el Grupo 2 (T=120s) mostró en general tasas de respuestas menores a las mostadas por el Grupo 1, manteniendose en un rango de 0.4 a 0.7 rpm. Sólo en el valor de P=0.1 se observó un decremento de la tasa llegando a valores de 0.2 rpm. En la condición de redeterminación se observó una mayor tasa asociada al subciclo tD. Bajo la condición contingente, las diferencias de ejecución entre los subintervalos fueron más evidentes. El subintervalo tD se mantuvo contante con valores cercanos a 0.2 rpm a lo largo de las variaciones de los valores de P, mientras que para el subintervalo tD se observó una función decreciente de la tasa conforme descendió el valor de P variando de 0.6 a 0.2 rpm., regresando a 0.6 rpm en la condición de redeterminación de P.
La Figura 2 muestra el tiempo promedio entre respuestas efectivas (TEREF) y la pausa postentrega de agua (PPEAP)[Y1], así como el porcentaje de entregas de agua producida [Y2] en cada una de las sesiones experimentales durante las cuatro fases experimentales para cada una de las ratas del Grupo 1. Los gráficos de esta figura también presentan los valores de duración del ciclo T y el valor de T/P que identifica el intervalo teórico entre entregas de agua (e.g. tiempo promedio entre entregas de agua, dada una respuesta). Los resultados mostraron variaciones en la ejecución de las tres ratas a lo largo de las primeras tres fases. Durante la Fase 1 (p=1.0) el valor de T y de T/P es de 30s., se observó en todas las ratas una función descendente en el TEREF, mostrando una tendencia a acercarse al valor del ciclo T en las últimas sesiones.Por su parte, la PPEAP mostró también valores irregulares durante la fase, acercandose al valor del ciclo T al final de la condición experimental. El porcentaje de entregas de agua producidas (EAP) mostró una función creciente en dos de las tres ratas (r32 y r34) cuyos porcentajes iniciales estuvieron entre el 20 y 40 % de las entregas programadas, llegando al final de la condición a un porcentajes de entre el 50 y el 100 %. La rata r33 mostró una mayor variabilidad a lo largo de la fase, sobre todo entre las sesiones 5 a la 15.
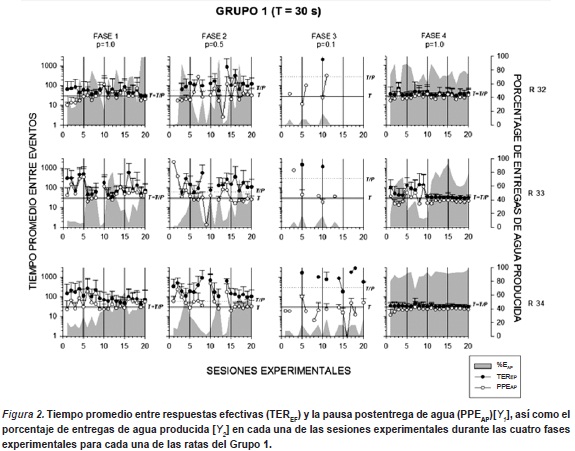
Durante las Fases 2 y 3 (p= 0.5 y 0.1, respectivamente), los datos mostraron que todas las ratas tuvieron una ejecución irregular a lo largo de las fases y un incremento en los TEREF y la PPEAP conforme disminuyó el valor de P, lo que a su vez se asoció a un decremento en el porcentaje relativo de EAP. En la fase 2, todas las ratas mostraron valores de TEREF por arriba de los 60s. (T/P), mientras que los valores de la PPEAP se mantuvieron cercanos a los 30 s. (T). Durante la Fase 3, el TEREF mostró valores superiores a los 1000 s. (T/P = 300) en todas las ratas, mientras que la PPEAP se mantuvo en valores cercanos a los 30 s. en dos ratas (r33 y r34). En ambas fases se observó un decremento en el porcentaje de EAP, llegando a valores entre 0 y 10 % durante la fase 3.
En la Fase 4 (p=1.0, sin estímulos correlacionados), en dos de las tres ratas (r32 y r34) se observó un patrón de ejecución regular respecto del TEREF y la PPEAP las cuales estuvieron en valores muy cercanos a los 30s. a lo largo de toda la fase. La rata r33 mostro variabilidad en ambas medidas durante la primera mitad de la fase, mostrando un ajuste a valores cercanos a los 30s. a partir de la sesión 10. Por su parte, se observó un incremento significativo en el porcentaje de EAP de todas las ratas, respecto de las fases experimentales anteriores. Las ratas r32 y r34 obtuvieron un porcentaje de EAP entre el 60 y el 95%, mientras que la rata r33 obtuvo un porcentaje entre el 20 y el 80%.
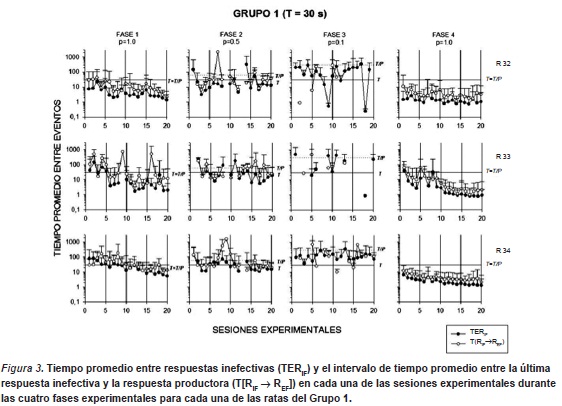
La Figura 3 muestra el tiempo promedio entre respuestas inefectivas (TERIF) y el intervalo de tiempo promedio entre la última respuesta inefectiva y la respuesta productora (T[RIF → REF]) en cada una de las sesiones experimentales durante las cuatro fases experimentales para cada una de las ratas del Grupo 1. Los gráficos de esta figura también presentan los valores de duración del ciclo T y el valor de T/P que identifica el intervalo entre entregas de agua teórico (e.g. tiempo promedio entre entregas de agua, dada una respuesta). Los datos mostraron funciones decrecientes en ambas medidas para todas las ratas en las Fases 1 y 4 (P=1.0), observándose un incremento en fases restantes (P=0.5 y 0.1). Durante la primera fase, todas las ratas mostraron valores de TERIF entre los 10 y 300s. durante las primeras 5 sesiones, disminuyendo hasta llegar a valores cercanos a los 2s. en las últimas 5 sesiones. En la Fase 2 (P=0.5) se observa un incremento en los valores de los TERIF en las tres ratas, fluctuando entre 6 y 200s. Estos valores se incrementan al pasar a la fase 3 (P=0.1) obteniendose valores entre 20 y 300s., con excepción de dos sesiones para la rata r32 y una para la rata r33 que obtuvieron valores de TERIF cercanos a 1s. Durante la Fase 4 (redeterminación de P=1.0), dos de las ratas (r33 y r34) mostraron funciones descendentes a lo largo de la fase en los valores de los TERIF llegando a valores cercanos a 1 s al final de la fase experimental. La rata r32 se mantuvo en valores cercanos a 1 s durante toda la fase experimental. Respecto del T[RIF → REF] las funciones obtenidas resultaron similares a las mostradas por los TERIF, en todas las ratas y en todas las fases, aunque éstas fueron sistemáticamente más altas.
La Figura 4 muestra el tiempo promedio entre respuestas efectivas (TEREF) y la pausa postentrega de agua (PPEAP)[Y1], así como el porcentaje de entregas de agua producida [Y2] en cada una de las sesiones experimentales durante las cuatro fases experimentales para cada una de las ratas del Grupo 2. Los gráficos de esta figura también presentan los valores de duración del ciclo T y el valor de T/P que identifica el intervalo entre entregas de agua teórico (e.g. tiempo promedio entre entregas de agua, dada una respuesta). En general los resultados mostraron variabilidad durante las tres primeras fases, disminuyendo ésta durante la fase de redeterminación del valor de P (Fase 4).
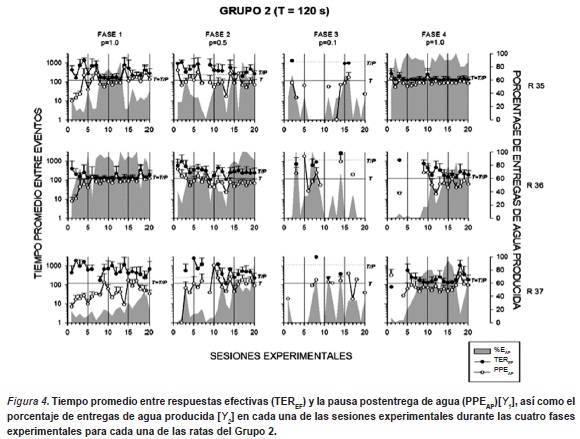
Durante la Fase 1 (p=1.0 y T/P= 120s.) dos de las tres ratas (r35 y r37) mostraron ejecuciones irregulares a lo largo de la condición experimental. Ambas ratas muestran valores de TEREF superiores a los 120s. durante las primeras sesiones de la condición. Sin embargo, después de la sesión 7, la rata r35 muestra un descenso en el valor de los TEREF llegando a valores cercanos a los 120s. Por su parte, la rata r37 mostró un patrón irregular en los TEREF a lo largo de toda la condición experimental mostrando en general valores por arriba de los 200s. En ambos sujetos, el porcentaje de EAP mostró funciones irregulares a lo largo de la fase observandose porcentajes entre 15 y 45% de entregas de agua obtenidas. Cabe resaltar en la rata r35 mostró un incremento considerable en el porcentaje de EAP, cuando sus TEREF se mantuvieron cercanos al valor de 120s., llegando a un porcentaje cercano al 80%. El sujeto r36, a diferencia de los sujetos anteriores, mostró una ejecución relativamente estable en los valores de los TEREF que se mantuvieron durante todas sesiones en valores cercanos a los 120s. Además, esta rata mostró valores de porcentaje de EAP superiores (entre 40 y 85%) a los observados en las ratas r35 y r37. Las PPEAP mostraron una función creciente hasta llegar a valores cercanos a 30s. en las tres ratas.
En la Fase 2 (p= 0.5 y T/P = 240s.) las tres ratas mostraron patrones variables de ejecución durante las primeras sesiones de la fase, observándose TEREF superiores a los 240s. Las ratas r35 y r36 mostraron una disminución en los valores de este indicador a partir de la sesión 8, observándose un ajuste irregular entre los 120 y 240s. durante el resto de las sesiones de la condición. La rata r37 mostró una variabilidad superior al resto de las ratas obteniendo valores de los TEREF superiores a los 300s. en la primera mitad de la fase, disminuyendo de manera irregular en la última mitad de la fase. La PPEAP siguio mostrando una tendencia a permanecer en valores cercanos a los 30s. en todas las ratas. El porcentaje EAP mostró una función de incremento gradual a lo largo de la condición en todas las ratas, variando en un rango de entre10 a 80% de entregas producidas.
La Fase 3 ( p= 0.1 y T/P = 1200s.) mostró una ejecución muy pobre en cuanto a TEREF. Las tres ratas mostraron que sólo en 3 o 4 sesiones se pudieron observar valores de este indicador, los cuales estuvieron en general cercanos a los 1200s. Por su parte, la PPEAP también mostró irregularidad entre e intra sujetos a lo largo de esta condición experimental. El porcentaje de EAP fue irregular a lo largo de la fase manteniendose en un rango de 0 a 60%, con la excepción de dos sesiones para la rata r36 que llegó a obtener el 100% de EAP.
Por último, en la Fase 4 (redeterminación P= 1.0, sin señalización) las ratas r36 y r37 obtuvieron ejecuciones irregulares durante la primera mitad de la condición (sesiones 1 a 10), en la que se observó TEREF con valores superiores a los 120s., mostrándose un decremento al inicio de la segunda mitad llegando a valores cercanos a los 120s. La rata r 35 mostró una ejecución regular a lo largo de toda la fase, observándose valores en los TEREF cercanos a los 120 s. durante las 20 sesiones. Con excepción de la rata r35, el resto de las ratas (r36 y r37) mostraron un incremento gradual del porcentaje de EAP (rango de 10 al 80%) a partir de la sesión 6 (r37) y la sesión 10 (r36). La rata r35 mantuvo un porcentaje de EAP por arriba del 80% a lo largo de toda la fase experimental.
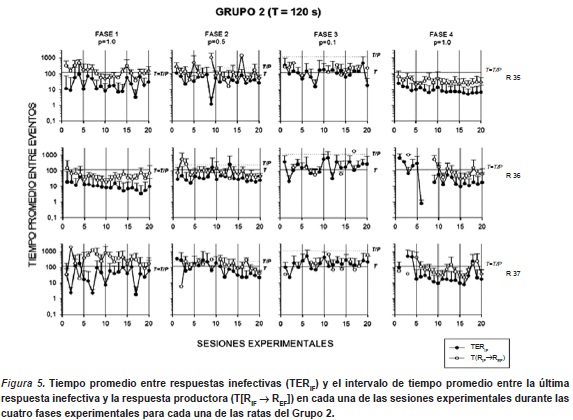
La Figura 5 muestra el tiempo promedio entre respuestas inefectivas (TERIF) y el intervalo de tiempo promedio entre la última respuesta inefectiva y la respuesta productora (T[RIF → REF]) en cada una de las sesiones experimentales durante las cuatro fases experimentales para cada una de las ratas del Grupo 2. Los gráficos de esta figura también presentan los valores de duración del ciclo T y el valor de T/P que iden-tifca el intervalo entre entregas de agua teórico (e.g. tiempo promedio entre entregas de agua, dada una respuesta). Los resultados generales con estos indicadores mostraron funciones decrecientes al interior de cada una de las fases, mientras que se observaron incrementos en las ejecuciones conforme disminuyó el valor de P (entre fases experimentales). Durante la primera fase, las ratas r35 y r37 mostraron ejecuciones irregulares respecto de los TERIF variando en un rango de 2 a 120s. Por su parte, la rata r36 mostró una ejecución descendente gradual a lo largo de la fase, variando en un rango de los 10 a los 20s. Estos valores se fueron incrementando en todas las ratas a lo largo de las Fases 2 y 3, asociadas a una disminución del valor de P. Durante la Fase 2 (P=0.5), los TERIF se mantuvieron en un rango entre los 30 y los 300s., mostrando los valores más bajos en las últimas 5 sesiones. Durante la Fase 3 (P=0.1), los TERIF fluctuaron entre 20 y los 500s. Por último, en la Fase 4 (redeterminación de P=1.0), las ratas r36 y r37 mostraron variabilidad en los valores de TERIF durante las primeras 10 sesiones (entre 10-700s.), tendiendo a estabilizarse a partir de la segunda mitad de la fase manteniendose en valores cercanos a los 10s. La rata r35 mostró una menor variabilidad en la ejecución (10 a 30s.), observándo una función gradual de decremento a lo largo de toda la fase, llegando a valores cercanos a los 10s. al final de la fase. Al igual que en el Grupo 1, los valores obtenidos del indicador T[RIF → REF] mostraron funciones similares a las obtenidas en los TERIF, en todas las ratas y en todas las fases, aunque éstas fueron sistemáticamente más altas.
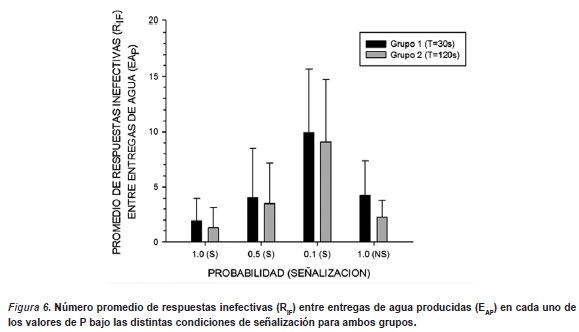
La Figura 6 muestra el número promedio de respuestas inefectivas (RIF) entre entregas de agua producidas (EAP) en cada uno de los valores de P bajo las distintas condiciones de señalización para ambos grupos. Los resultados muestran funciones parecidas de incremento gradual en el número de RIF conforme desciende el valor de P, observándose una disminución de éste indicador en la condición de redeterminación de P=1.0, sin señal. Aunque las funciones obtenidas en ambos grupos son similares, en el Grupo 1 (T=30s.) se observó una mayor frecuencia promedio de respuesta (entre 2 y 10 RIF), respecto del Grupo 2 (T=120s.) que obtuvo frecuencia promedio de respuesta levemente menor (entre 1 y 9 RIF). Además se pudo observar que en la condición de redeterminación, el promedio de RIF no regresó a los valores iniciales obtenidos en la primer condición, sino que se mantuvieron relativamente más altas en ambos grupos.
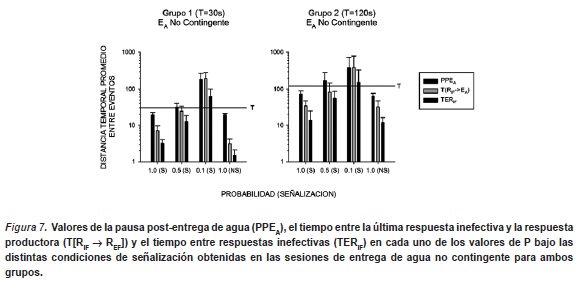
La Figura 7 muestra los valores de la pausa post-entrega de agua (PPEA), el tiempo entre la última respuesta inefectiva y la respuesta productora (T[RIF → REF]) y el tiempo entre respuestas inefectivas (TERIF) en cada uno de los valores de P bajo las distintas condiciones de señalización obtenidas en las sesiones de entrega de agua no contingente para ambos grupos. En general se observaron funciones ascendentes en cada uno de estos indicadores conforme disminuye el valor de P, mostrando un posterior descenso cuando se expone a la condición de redeterminación a 1.0 de P. Estas funciones fueron similares en ambos grupos, cuya diferencia tan sólo fue que para el Grupo 2 (T=120s.) los valores de los indicadores fueron levemente mayores al Grupo 1 (T=30s.).
Discusión
La presente discusión se ha dividido en tres secciones con el fin de acotar los resultados obtenidos en el presente estudio en función de: a) las condiciones de contingencia-no contingencia prescritas en los programas temporales empleados y; b) las posibles implicaciones que estos pudieran tener con las funciones contex-tual (f [cx]) y suplementaria ( f[su]) descritas en la taxonomía propuesta por Ribes y López (1985).
Indicadores de ejecución bajo condiciones de entrega contingente
Los resultados de este trabajo son consistentes con los datos obtenidos en estudios previos utilizando programas temporales (Ribes & Torres, 1996; Ribes, Mayoral, Torres & Ibañez, 2000; Ribes, Torres & Mayoral, 2000) al observar patrones de ejecución variables, sobre todo en las fases experimentales donde el parámetro P fue menor a 1.0. Sin embargo, en el presente estudio se pudo observar que, independientemente del valor del ciclo T (T= 30s. en el Grupo 1 y T=120s. en el Grupo 2), cuando P tuvo un valor de 1.0 (Fase 1 y Fase 4 para ambos grupos), la variabilidad en los TEREF mostró una tendencia a disminuir y acercarse a los valores prescritos por la contingencia experimental (valor del ciclo T). Este efecto fue más notorio partir de la segunda mitad de cada condición experimental. Este dato, aunado al incremento gradual en el porcentaje de entregas de agua producida (EAP) parecieran sugerir que, por lo menos bajo estas condiciones, un proceso gradual de ajuste a las contingencias prescritas por el programa por parte de las ratas en tanto que el valor del tiempo entre este tipo de respuestas se acerca al valor del ciclo T, cumpliendo así con el criterio de suficiencia para la suplementación de eventos de estímulo. Ahora bien, si incorporamos en el análisis el TERIF, podemos deducir que en la medida en que este indicador disminuye, la proporción de respuestas por ciclo aumenta, favoreciendo un incremento de la probabilidad de que una respuesta a la palanca cumpla con el criterio temporal prescrito por la contingencia del programa. Por su parte, otro indicador respecto del ajuste a las contingencias también nos lo otorga la PPEAP, ya que en ésta se evidenció una tendencia a igualar los valores de duración del ciclo (T) o respecto de los valores de los tiempos teóricos entre entregas de agua producida (T/P) en ambos grupos, lo que pudiera estar indicando sensibilidad al cambio de las contingencias programadas.
Cabe destacar que, en la medida de que el valor de P permanece constante en 1.0 durante las fases 1 y 4, asegura que en cada ciclo T exista la posibilidad (disponibilidad) de la producción de la entrega de agua, lo que implica una situación experimental relativamente constante. Ahora bien, el decremento en los valores de P (0.5 y 0.1) implica dos cambios importantes en el programa. En primer lugar, el tiempo relativo entre entregas de agua contingente a la respuesta efectiva, se incrementa en función del valor del ciclo T y del valor del parámetro P ó (T/P). En segúndo lugar, la aleatoriedad inherente en la asignación de ciclos T con disponibilidad en la entrega de agua supone ya no un criterio temporal específico a cubrir por una respuesta. Sino que, dadas estas condiciones, las respuestas productoras que tienen una mayor probabilidad de ocurrencia están comprendidas en un rango temporal cuyos límites parecen estar entre el valor del ciclo T (como límite inferior) y el valor teórico delimitado por T/P (como límite superior). Esto implíca una relación de contingencia más compleja en términos de contacto, en la medida en que la producción de estímulos es condicional ya no a una posición regular en tiempo de la respuesta (T), como ocurre en condiciones de P=1.0, sino que la disponibilidad para la producción de estímulo se vuelve irregular en tiempo, en función del decremento de P (T/P).
Los datos obtenidos en las fases 2 y 3 mostraron variabilidad en los tiempos entre respuestas tanto efectivas como inefectivas observándose ejecuciones por arriba de los identificados por el valor de T/P en ambos grupos. Estos resultados vinculados con la irregularidad observada en el porcentaje de EAP confirman que las ratas no hicieron contacto con el tipo de contingencias programado. La única excepción a esto se observó en la rata r36, cuyos TEREF se mantuvieron entre los 120 y los 240s. a partir de la segunda mitad de la Fase 2, lo que posibilitó un incremento en el porcentaje de EAP.
Por otro lado, en este estudio no se evidencia de manera conclusiva la función señal (Ribes, 1997) de los estímulos correlacionados respecto del desempeño de las ratas. Nuevamente, las fases 1 y 4 son las que posibilitarían algún tipo de comparación. Los datos en la fase 4 muestran que hay un mayor porcentaje de EAP, asociado a una mayor estabilidad en cuanto a los TEREF respecto de la Fase 1. También se observa que durante la Fase 1, los TERIF fueron mayores (entre los 30 y los 10s.) a los observados en la fase 4 (entre los 10 y valores cercanos a 1s.). Este efecto fue más evidente en el Grupo 1 (T=30s.). Esto pareciera indicar que ante la ausencia de estímulos correlacionados, los TERIF disminuyen, favoreciendo así el contacto con la contingencia programada. Sin embargo, también existe la posibilidad de que el aumento en la densidad de respuesta por ciclo este asociada con el incremento la densidad de EAP asociadas al cambio en el valor de P (de 0.1 a 1.0). Por lo tanto, se vuelve necesaria una evaluación experimental de dichos supuestos bajo condiciones controladas de señalización y cambio de valores de P.
Indicadores de ajuste bajo condiciones de entrega no-contingente
Como se mencionó anteriormente, una de las grandes virtudes de los programas temporales tiene que ver con la posibilidad procedimental de poder estructurar contingencias de ocurrencia de eventos de estímulo de manera independiente a la actividad del organismo. En este estudio se evaluó dicha característica a través de la presentación no contingente de entregas de agua durante las sesiones de sondeo.
Los resultados obtenidos en este estudio son consistentes con los obtenidos en estudios previos (Bruner, 1981; Ribes y López, 1979 a y b; Ribes y Torres, 1996) en la que se observa una densidad de respuesta menor en las sesiones de sondeo con la entrega de agua no contingente, respecto de las sesiones experimentales con entrega de agua contingente a la respuesta de la rata.
Con el fin de evaluar el nivel de ajuste a las contingencias programadas bajo condiciones de entrega no contingente, se analizaron tres tipos de relaciones temporales:
1. Tiempo entre la EA y la RIF precedente,
2. Tiempo entre la EA y la RIF consecuente, y;
3. Tiempo entre RIF. Los resultados mostraron que en ambos Grupos la ejecución mantuvo una función creciente conforme disminuyó el valor de P, independientemente del valor de T. Además, en todas se observó un decremento abrupto durante la fase de redeterminación. Esto pareciera sugerir que las ratas hicieron contacto con las contingencia establecidas, en la medida en que las propiedades temporales de la respuesta se acercaron a los tiempos entre entregas de agua (EA), lo que pudiera sugerir un ajuste diferencial del responder respecto de los eventos de estímulo programados.
Es importante señalar que este indicador de ajuste en este estudio se vuelve pertienente en la medida que las ratas primero fueron expuestas a condiciones de producción contingente en la entrega de agua, lo que favorece que se desarrollen relaciones funcionales respecto de la respuesta a la palanca, que al parecer no se pierden durante el periodo de no contingencia. Por lo tanto se vuelve relevante explorar la pertinencia de estos indicadores cuando estas las condiciones no impliquen una historia de suplementación.
Funciones de respuesta: indicadores de ajuste efectivo y diferencial
El dato primario en los programas de reforzamiento se ha obtenido históricamente a partir de la distribución en tiempo de la emisión de respuestas bajo condiciones específicas de reforzamiento (Ferster & Skinner, 1957). Consecuentemente, la tasa y frecuencia de respuesta emergieron como las unidades de medidas necesarias para sostener el criterio de reproducibilidad definitoria de la clase operante y los programas de reforzamiento se concentraron en la reproducción de ejecuciones conductuales lo menos variables posibles con el fin de poder replicar e identificar patrones típicos de ejecución que fueran característicos de las condiciones experimentales específicas (Carpio, 1992; Ribes, 1994; Ribes, 1999).
Sin embargo, al considerar al comportamiento psicológico como una estructura de relaciones de contingencia en las que participa el organismo, y no como una relación genérica entre clases de estímulo y respuesta, la necesidad de mostrar indicadores de "cuantas veces" ocurre la relación (criterio de frecuencia), o que tan "fuerte" es dicha relación (criterio de tasa), deja de ser pertinente como criterio demostrativo de ajuste. De aquí que se vuelve imprescindible identificar la naturaleza y las condiciones de las relaciones de contingencia a las que se expone y con las que interactúa el organismo a partir de su responder.
De esta manera, podríamos considerar que, en el caso particular de los programas temporales las contingencias de ocurrencia se establecen primordialmente con base en las propiedades temporales de ocurrencia de los eventos que participan en estos procedimientos, incluyendo las propiedades temporales de ocurrencia de la conducta del organismo en tanto que, si se considera la función suplementaria, la ubicación temporal de la respuesta respecto los eventos de estímulo con los que interactúa posibilita que se cumplan las condiciones de necesidad y suficiencia para su producción. Por su parte, cuando la ocurrencia de los eventos de estímulo no depende de manera directa de la conducta de los organismos, es probable que esto se vea reflejado respecto de la distancia temporal de las instancias particulares de respuesta, a partir de los patrones de presentación estipulados en las contingencias programadas.
A partir de lo anterior, nos gustaría destacar que en el presente trabajo, aunque el análisis se hizo respecto de una topografía particular de respuesta (presión de la palanca), se pudieron identificar dos tipos distintos de indicadores de ajuste a partir de las propiedades temporales del responder, en términos de su capacidad de producir o no cambios en las relaciones entre eventos de estímulo: a) Respuestas Efectivas, cuya ubicación temporal cumple con el criterio estipulado por la contingencia, y; b) Respuestas Inefectivas, que identifican a todas aquellas respuestas que no cumplen con el criterio de suficiencia para la producción de agua.
Los datos obtenidos en el presente estudio sugieren funciones diferenciales entre este tipo de respuestas. Como ya se ha venido mencionando, las respuestas efectivas permiten mediar las relaciones entre eventos de estímulo en la medida en que sus propiedades temporales se ajustan a las contingencias prescritas por el programa. Por su parte, las respuestas inefectivas parecieran tener una función de muestreo de la situación experimental. Esto se sugiere a partir de que el desempeño de las ratas incrementa en su efectividad conforme disminuye el TERIF. Además, este efecto se vuelve más notorio ante condiciones de ausencia de estimulos correlacionados, lo que parecería indicar una interacción entre su función de señal y este tipo de respuestas.
Los resultados obtenidos en este experimento sugieren que el análisis de los cambios en las propiedades temporales del responder respecto de los eventos de estímulo con los que se inte-ractúa, utilizando los programas temporales como estrategia metodológica, permiten describir cambios a nivel molecular (Ribes, 2007) que podrían indicar modos en que las ratas entran en contacto con las contingencias prescritas por los programas, además de que posibilitan un acercamiento cuantitativo al tipo de ajuste que se estructura en función de la producción y entrega independiente de eventos de estímulo.
De aceptarse los argumentos aquí propuestos, se implicaría la necesidad de identificar indicadores de contacto y ajuste a las contingencias, a partir de las estrategias metodológicas pertinentes de acuerdo con el tipo de interacción que se quiera estudiar (Ribes & López, 1985).
La distribución de los TEREF y TERIF observada en este estudio sugieren que existe un efecto diferencial entre condiciones de presencia y ausencia de estímulos correlacionados. Además, la regularidad en la ejecución obtenida en la fase de redeterminación (Fase 4) sugiere que pudiera existir un efecto asociado al cambio en los valores del parámetro P. A partir de lo anterior, resulta importante que en estudios posteriores se identifquen de manera más controlada las condiciones de "señalización" y de cambio de probabilidad con el fin evidenciar su efecto en el tipo de contacto y ajuste a las contingencias programadas.
Referencias
Brown, P.L. & Jenkins, H.M. (1968) Auto-shaping of the pigeon's key-peck. Journal of Experimental Analysis of Behavior, 11, 1-8. [ Links ]
Bruner, C.A. (1981) The effect of cycle lenght, inter-stimulus interval and probability of reinforcement in autoshaping. Revista Mexicana de Análisis de la Conducta, 7, 149-157. [ Links ]
Cabrer, F., Daza, C. & Ribes, E. (1975) Teoría de la conducta: ¿Nuevos conceptos o nuevos parámetros?. Revista Mexicana de Análisis de la Conducta, 1, 191-212. [ Links ]
Carpio, C. (1992) Transición paradigmática en psicología: Análisis de un caso. En Acta Comportamentalia, 0, 85-108. [ Links ]
Falk, J.L. (1971) The nature and determinants of adjunctive behavior. Psychology & Behavior, 6, 577-588. [ Links ]
Ferster, C.B. & Skinner, B.F. (1957) Schedules of Reinforcement. New York: Appleton-Century Crofts. [ Links ]
Kantor, J.R. (1924/1929) Principles of Psychology. Chicago: Knopf (reeditado por Principia Press, 1949). [ Links ]
Lee, R., Sturmey, P. & Fields, L. (2007) Schedule-induced and operant mechanisms that influence response variability: A review and implications for future investigations. The Psychological Record, 57, 429-455. [ Links ]
Ribes, E. (1994) Skinner y la psicología: lo que hizo, lo que no hizo y lo que nos toca hacer. En E. Ribes (Ed.) B.F. Skinner: in memoriam. México: Universidad de Guadalajara. [ Links ]
Ribes, E. (1995) Causalidad y Contingencia. Revista Mexicana de Análisis de la Conducta, 21, 123-142. [ Links ]
Ribes, E. (1997) The stimulus in behavior theory: event or function? Revista Mexicana de Análisis de la Conducta, 23, 147-160. [ Links ]
Ribes, E. (1999) Teoría del condicionamiento y lenguaje. México: Taurus, Universidad de Guadalajara. [ Links ]
Ribes, E. (2004) Acerca de las funciones psicológicas: Un post-scriptum. Acta Comportamentalia, 12, 117-127. [ Links ]
Ribes, E. & López, F. (1979a) Efectos de un estímulo delta en la adquisición de respuestas concurrentes bajo un programa definido temporalmente. Revista Mexicana de Análisis de la Conducta, 5, 27-40. [ Links ]
Ribes, E. & López, F. (1979a) La adquisición de operantes concurrentes bajo un programa señalado de reforzamiento definido temporalmente. Revista Mexicana de Análisis de la Conducta, 5, 41-56. [ Links ]
Ribes, E. & López, F. (1985) Teoría de la Conducta: Un análisis de campo y paramétrico. México: Trillas. [ Links ]
Ribes, E. Mayoral, A., Torres, C. & Ibañez, F. (2000) Effects of auditory stimuly correlated with different probabilities of water delivery in a limited –hold temporal schedule. Behavioural Processes, 52, 49-59. [ Links ]
Ribes, E. & Torres, C. (1996) Efectos de la variación en la probabilidad de reforzamiento correlacionada con dos estímulos neutros en un programa definido temporalmente. Revista Mexicana de Análisis de la Conducta, 22, 41-78. [ Links ]
Ribes, E., Torres, C. & Mayoral, A. (2000) Efectos de la ausencia y presencia con estímulos correlacionados con distintas probabilidades de reforzamiento y con extinción en programas definidos temporalmente. Revista Mexicana de Análisis de la Conducta, 26, 327-354. [ Links ]
Ribes, E., Zepeda, I., Arenas, S. & Mayoral, A. (2007) Efecto de la densidad local de entrega de agua en la frecuencia total de respuesta en un programa temporal señalado. En Revista Mexicana de Análisis de la Conducta, 33, 139-166. [ Links ]
Schoenfeld, W.N. & Farmer, J. (1970) Reinforcement schedules and the behavior stream. En The Theory of reinforcement schedules, W.N. Schoenfeld (Ed.), New York: Appleton-Century Crofts. [ Links ]
Schoenfeld, W.N., Cumming, W.W. & Hearst, E. (1956) On the classifIcation of reinforcement schedules. Proceedings of the Nacional Academy of Science, 42, 563-570. [ Links ]
Schoenfeld, W.N., Cole, B.K., Lang, J. & Mankoff, R. (1973) "Contingency" in behavior theory. En Contemporary Approaches to Conditioning and Learning, F.J. McGuigan & D.B. Lumsden (Eds.) New York: John Wiley and Sons. [ Links ]
Skinner, B.F. (1948) "Superstition" in the pigeon. Journal of Experimental Psychology, 38, 168-172. [ Links ]
Torres, C. (2009) Análisis de los parámetros asociados a la entrega del reforzador en programas temporales. En M.A. Padilla (Ed) Avances en la investigación del comportamiento animal y humano. México: Universidad de Guadalajara. p. 65-78. [ Links ]
Torres, C., Villamil, W. & Calleros, K. (2009) Efectos de la disminución de la probabilidad y la disponibilidad de reforzamiento en programas temporales de distinta duración. En Revista Mexicana de Análisis de la Conducta, 35, 87-100. [ Links ]
Waissman, A. (1961) Impairment of performance when a discriminative stimulus is correlated with reinforcement contingency. Journal of Experimental Analysis of Behavior, 4, 365-369. [ Links ]
Waissman, A. (1963) Behavioral effects of pairing and SD with decreasing limited-hold reinforcement schedule. Journal of Experimental Analysis of Behavior, 6, 265-268. [ Links ]

