











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Las pitayas o pitahayas forman parte del orden Cactales, familia Cactaceae, subfamilia Cereoideae, tribu Hylocereae, subtribu Hylocereinae, con representación de dos géneros de importancia agrícola: Hylocereus (Berger) Br y R., que corresponde a frutos de epidermis roja y Selenicereus (Berger) Br. y R., con frutos tetraploides de epidermis amarilla (Nobel 2002; Rodríguez 2002) y con por lo menos, 20 especies reconocidas (Barthlott y Hunt 1993; Tel-Zur et al. 2004). S. megalanthus Bauer (K Schumann ex Vaupel) Moran, 1953, comúnmente conocida como pitaya amarilla, tiene su origen en Colombia, Perú, Bolivia, Ecuador y Venezuela; corresponde a plantas perennes, terrestres trepadoras y epifitas, que requieren de soporte para su producción comercial. Su propagación comercial es vegetativa a partir de tallos o esquejes, la producción de frutos inicia entre uno a dos años después del establecimiento y puede tener una vida productiva mayor a 10 años (Rodríguez 2002).
Colombia es el principal productor y exportador de pitaya amarilla a nivel mundial, con áreas sembradas en Boyacá, Valle del Cauca, Bolívar, Caldas, Cundinamarca, Huila, Quindío, Risaralda, Santander y Tolima (Betancourt et al. 2010). Las zonas más adecuadas para el cultivo se encuentran entre 1.400 y 1.700 msnm, con temperaturas entre 14 a 26 °C, suelos con pH entre 5,5 y 6,5 y precipitaciones de 1.300 a 2.200 mm anuales (Rebolledo et al. 2003). Aunque el municipio de Inzá, Cauca se considera zona ecológica marginal para este cultivo, se establecieron de novo cultivos de pitaya amarilla a partir de 2015, como alternativa de diversificación y como una respuesta a la reducción de la rentabilidad del cultivo de café, tradicionalmente sustento económico. A pesar de los altos costos para el establecimiento de este cultivo, el proceso se llevó a cabo sin asistencia técnica o estudios que permitieran estimar las limitantes para su producción y comercialización. El objetivo de este estudio fue caracterizar la entomofauna asociada a cultivos de pitaya amarilla de tres predios productivos, con características de la composición del paisaje agrícola contrastantes, del municipio de Inzá, Cauca, Colombia.
Materiales y métodos
Sitio de estudio
El municipio de Inzá, Cauca, se encuentra ubicado a los 02º33’24”N y 76º04’00”O; con cabecera municipal ubicada a una altitud de 1.800 msnm, temperatura promedio anual de 19 °C y precipitación media anual de 1.526 mm (Gobernación del Cauca 2013). El estudio se llevó a cabo en tres predios productivos para pitaya amarilla localizados en distintas zonas altitudinales y con diferentes arreglos agronómicos de producción. 1) Predio Piedra Mesa (02º30’10,63”N, 076º02’58,22”O) de la vereda Turminá, a una altitud de 1.804 msnm, con el cultivo pitaya amarilla en pendiente de 33 % y tutorado vivo de árboles maderables como flor amarillo (Handroanthus chrysanthus (Jacq.) S, O. Grose, 1797) combinados con alambre y cultivos transitorios como maíz (Zea mays L., 1753). En la periferia, el cultivo está rodeado de árboles de mandarina (Citrus reticulata Blanco, 1837) y aguacate (Persea americana Mill., 1768). 2) Predio Buena Vista (02º31’33,6”N, 076º05’06,1”O) de la vereda Guanacas, ubicado al lado oeste del Rio Ullucos, a 1.743 msnm, con cultivo de pitaya amarilla en pendiente de 15 % y tutorado vivo de maderables en asocio con yuca (Manihot esculenta Crantz, 1766). 3) Predio La Cabaña (02º31’16,6”N, 76º05’52,1”O) surcado al sureste por el río Ovejas, se encuentra localizado en la vereda Mesopotamia a 1.859 msnm. El cultivo de pitaya amarilla se estableció en terreno con pendiente de 25 % y con tutorado vivo de árboles maderables de flor amarillo y alambre, en asocio con plantas de café (Coffea arabica L., 1753), guayaba (Psidium guajava L., 1753) y guadua (Guadua angustifolia Kunth, 1822) (Fig. 1). La densidad de siembra para el cultivo de pitaya amarilla es de 1,5 m entre plantas por 3,0 m entre surcos en cada predio. El manejo agronómico se realiza experimentalmente, con control de arvenses manual y cosecha de frutos teniendo en cuenta el grado de madurez determinado por el color (Martínez et al. 2013). Durante el estudio, se observó permanente floración y fructificación en las plantas y no se realizaron medidas de control fitosanitario a los cultivos.
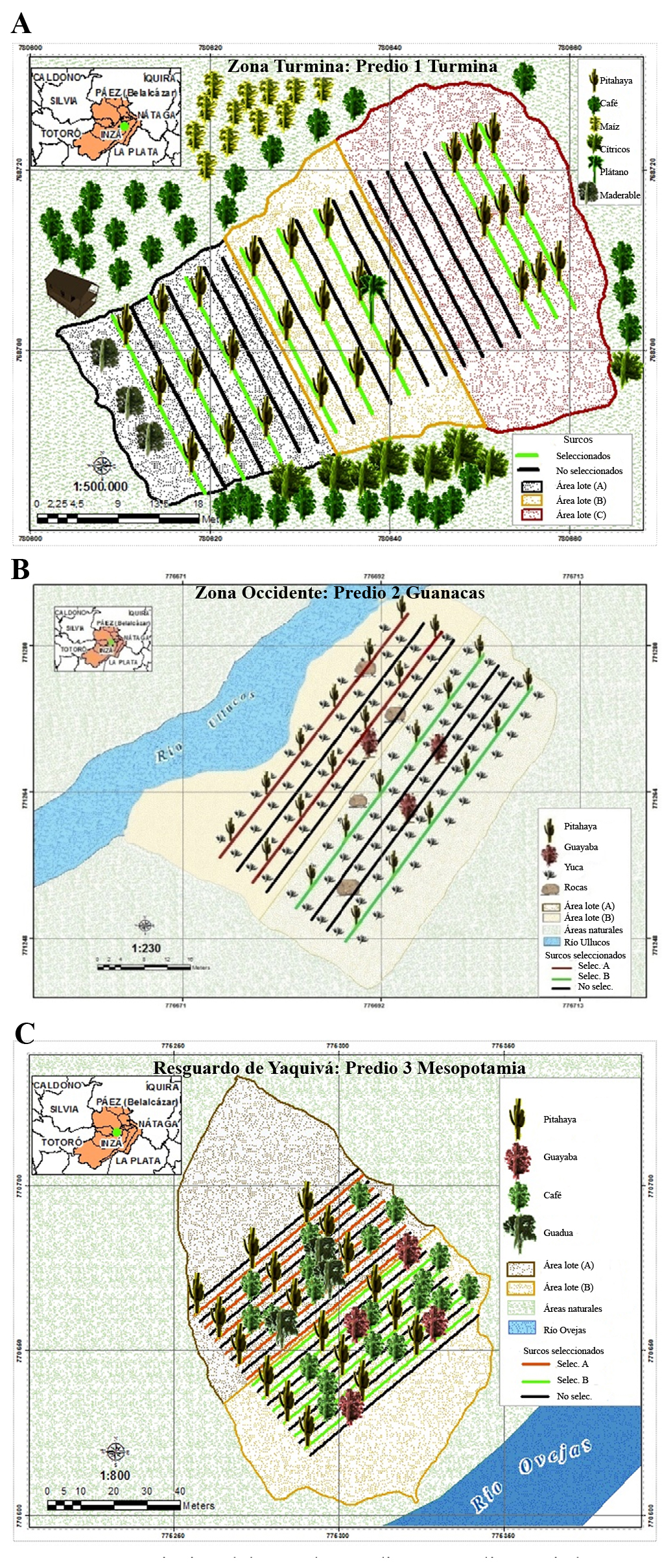
Figura 1 Descripción del área de estudio. A. Predio 1: Piedra Mesa, Vereda Turminá. B. Predio 2: Buena Vista, Vereda Guanacas. C. Predio 3: La Cabaña, Vereda Mesopotamia. Fuente: González et al. 2018.
Componente entomológico
Se realizó un estudio de tipo transversal con tres eventos de muestreo asociados al régimen de lluvias del municipio, así: junio 2016 (100-150 mm), septiembre 2016 (100-150 mm) y enero 2017 (50-100 mm) (rangos de precipitación obtenidas de estaciones climáticas del IDEAM, próximas a los predios evaluados). Los muestreos se realizaron por bloques definidos de acuerdo a la pendiente de cada predio, considerando un tamaño de muestra del 5 % por predio. El predio Piedra Mesa estuvo compuesto por 3 bloques y 27 unidades muestrales, el predio Buena Vista por 2 bloques con 24 unidades muestrales y el predio La Cabaña por 2 bloques con 36 unidades muestrales. Cada planta constituyó una unidad muestral y los muestreos se realizaron entre las 09:00 y 12:00 horas. Se consideraron variables fenológicas como longitud del cladodio más largo de la planta, número de cladodios, número de botones florales y número de frutos.
Los insectos se recolectaron empleando dos métodos realizados de forma simultánea por cultivo: el primero consistió en aspiración bucal y recolección manual directa de los insectos sobre órganos vegetales en todo el componente epigeo de la planta. En el segundo método se realizaron pases dobles con red entomológica a 0,50 m del tallo de la planta bajo el área foliar y 0,50 m del suelo. El material entomológico se preservó en alcohol al 70 %, en viales individuales etiquetados de acuerdo a la ubicación de la planta en el cultivo. La determinación taxonómica de artrópodos se realizó con base en caracteres morfológicos externos según claves taxonómicas (Sharp 1889; Linnavuori 1959; Korytkowski y Ojeda 1971; González y Carrejo 1992; Norrbom y McAlpine 1996; Delvare et al. 2002; Habeck 2002; Korytkowski 2003; Resh y Cardé 2003; Triplehorn y Johnson 2005; Dietrich 2005; Brown et al. 2009; Jones y Deitz 2009; Thyssen 2010; Hernández 2013; Fernandes et al. 2015). La determinación de individuos de la familia Lonchaeidae (Diptera) se realizó con base en caracteres morfológicos (Castro 2012) y confirmación con experto.
Para cada grupo insectil encontrado se determinó la abundancia relativa y su asociación con las variables fenológicas de la planta. Se calcularon los índices ecológicos de riqueza específica de especies (Margalef, DMg), de diversidad (Shannon-Wiener, H’) y de recambio de especies (Whittaker, βw). Los resultados obtenidos en campo se sometieron a un análisis descriptivo y de varianza con un nivel de significancia de P < 0,05 con comprobación de los supuestos de homogeneidad (prueba de Levene) y normalidad (prueba Kolmogorov-Smirnov). Las variables que no se ajustaron a la distribución normal se analizaron mediante pruebas no paramétricas (Test de Kruskal-Wallis y Kendall) con un nivel de significancia P < 0,05. La relación entre la composición de la entomofauna, variables climáticas y fenológicas de las plantas se estimaron mediante correlación de Kendall (Calderón y Alzamora 2011). Todos los análisis estadísticos se realizaron con SPSS 22© (IBM Corp. Released 2013) siguiendo los métodos propuestos por Guisandé et al. (2013).
Resultados
En el muestreo directo de insectos asociado a cada órgano y estado fenológico de las plantas, en todos los predios y eventos de muestreo, se recolectó un total de 170 individuos representados por 9 órdenes y 58 familias (Tabla 1). En ningún caso se observaron daños en las plantas atribuibles a especies registradas como limitantes al cultivo (Kondo et al. 2013a).
Tabla 1 Familias de insectos encontrados en predios productivos para pitaya amarilla, Inzá, Cauca 2016-2017.
| Orden | Familia | Piedra Mesa (n) | Buena Vista (n) | La Cabaña (n) |
|---|---|---|---|---|
| Diptera | Syrphidae | - | Cb, Cm, (4) | Cm, B (3) |
| Lonchaeidae | Ca, Cm, F (4) | F (1) | - | |
| Tachinidae | Cm (1) | - | Cm, B (2) | |
| Hemiptera | Reduviidae | - | Cb (1) | Cm (2) |
| Miridae | - | Cm (1) | Cm (1) | |
| Coreidae | Cm (10) | - | - | |
| Coleoptera | Chrysomelidae | Cm (2) | Ca, Cm (4) | |
| Coccinellidae | Ca (1) | Cm (3) | ||
| Hymenoptera | Formicidae | Ca, Cm, B (8) | Ca, Cm, F (7) | Pa, B (7) |
| Apidae | B, F (2) | - | B (1) | |
| Braconidae | Cm (3) | - | ||
| Ichneumonidae | - | Pa, F (2) | - |
Ca: Cladodio zona alta. Cm: Cladodio zona media. Cb: Cladodio zona baja. F: Fruto. B: Botón.
La familia Lonchaeidae (Diptera) estuvo representada por tres individuos del género Lonchaea y cuatro de Dasiops. Todos correspondieron a machos, lo que imposibilitó la determinación a especie, debido a que se requiere el análisis de la forma del ovipositor (Castro 2012). Lonchaea se reconoció por la presencia de lúnula pequeña claramente pilosa con cerdas en el margen superior, antenas casi juntas en la base con un tercer segmento antenal alargado y de forma ovalada que sobrepasa levemente el margen oral y una cerda esternopleural. Dasiops se reconoció por presentar una lúnula amplia pubescente, base de las antenas ampliamente separada y un tercer segmento antenal corto y redondeado (Korytkowski y Ojeda 1971). Las hormigas recolectadas por el método 1 se hallaron en los cladodios, frutos y botones en todos los predios. Se determinaron como individuos del género Crematogaster spp., al presentar lóbulos frontales bien desarrollados y expandidos, glándula metatibial ausente; espiráculos de los segmentos dos a cuatro del gáster no expuestos, escondidos bajo el tergo de los segmentos anteriores (Palacio y Fernández 2003; Fernández y Sharkey 2006). La familia Coreidae (Hemiptera) estuvo representada por estados inmaduros asociados siempre a cladodios de la zona media de las plantas. Su determinación taxonómica se basó en la ausencia de tricobotrias en la cabeza pero presentes ventralmente en el abdomen, primordios de las alas mesotorácicas cercanos entre sí, lóbulo escutelar sin porción libre, glándulas abdominales levemente separadas posteriormente, tergum V no constreñido en la línea media, antenas con el primer segmento antenal más corto que la longitud conjunta entre el tórax y la cabeza; y en vista lateral, la antena se inserta por encima de la línea entre el centro del ojo y el ápice del tylus (Herring y Ashlock 1971). Posiblemente los especímenes corresponden a la subfamilia Coreinae al presentar coxas cercanas entre si, ausencia de glándula metatorácica, presencia de estrías en el fémur y ausencia de espinas en la tibia (Fernandes et al. 2015).
No se encontraron diferencias estadísticamente significativas en la distribución espacial de los individuos entre los bloques o entre la abundancia relativa de insectos para todos los eventos de muestreo en los predios evaluados (Tabla 2). Sin embargo, para el predio Piedra Mesa la mayor abundancia relativa de insectos se obtuvo en cladodios medios y altos para todos los eventos de muestreo, mientras que solo hubo asociación de individuos con flores y frutos en los muestreos 2 y 3. En el predio Buena Vista, la mayor cantidad de individuos se encontró en cladodios altos, medios y bajos y no se encontraron individuos en botones y en flores. En el predio La Cabaña, la fauna insectil se encontró asociada a cladodios altos, medios y bajos de las plantas (Figs. 2A-2C). Sin embargo, se encontró correlación positiva entre plantas con mayor densidad de cladodios y el número de individuos la familia Lonchaeidae (τ = 1,0; P = 0,00) y estos con las variables ‘temperatura’ (τ = 1,0; P = 0,00) y ‘precipitación’ (τ = 1,0; P = 0,00). En el predio Buena Vista, las poblaciones de las familias Formicidae y Syrphidae presentaron una correlación positiva con la ‘humedad relativa’ (τ = 1,00; P = 0,007). El comportamiento de la familia Lonchaeidae en el predio La Cabaña presentó correlación positiva con la variable número de frutos (τ = 1,0; P = 0,00).
Tabla 2 Comparación de la distribución espacial y abundancia relativa de insectos entre muestreos para los predios evaluados P > 0,05. Test de Kruskal-Wallis y Kendall.
| Predio | Bloques | Eventos de muestreo | ||
|---|---|---|---|---|
| M1 | M2 | M3 | ||
| Piedra Mesa | P = 0,278 | P = 0,721 | P = 0,897 | P = 0,463 |
| Buena Vista | P = 0,023 | P = 0,889 | P = 0,340 | P = 0,028 |
| La Cabaña | p = 0,380 | P = 0,224 | P = 0,107 | P = 0,760 |
M1 = muestreo 1, M2 = muestreo 2, M3 = muestreo.
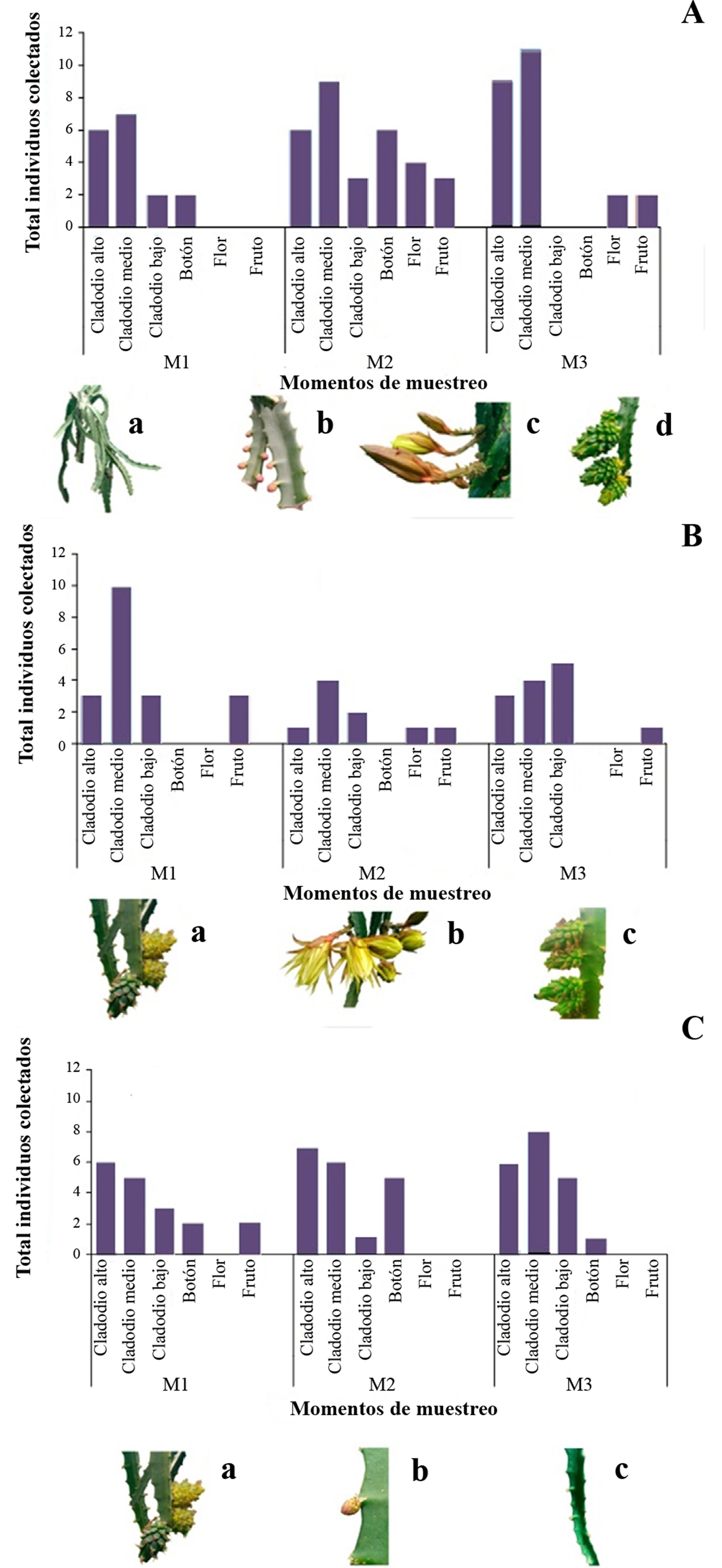
Figura 2 Población total (n = 170) total de insectos recolectados en los órganos vegetales de pitaya amarilla (Selenicereus megalanthus) por evento de muestreo (M1, M2 y M3) en el municipio de Inzá, Cauca. M1 (100-150 mm), M2 (100-150 mm) y M3 (50-100 mm). A. Predio 1: Piedra Mesa, Vereda Turminá. B. Predio 2: Buena Vista, Vereda Guanacas. C. Predio 3: La Cabaña, Mesopotamia. Fenología de la planta encontrada para cada evento de muestreo: a. Cladodios. b. Botones. c. Flores. d. Frutos. Fuente: González et al. 2018.
La biodiversidad basada en insectos en los predios evaluados reveló una riqueza equiparable con sistemas de baja intervención antrópica (Piedra Mesa DMg = 7,01; Buena vista DMg = 6,46; La Cabaña DMg = 7,42), siendo ligeramente menor en el predio Buena Vista, con menor diversidad de plantas y a menor altitud. Entre predios se encontró alta heterogeneidad con diversidad moderada de insectos, ligeramente mayor en el predio de La Cabaña con menor intensidad de prácticas agronómicas (Índices de Shannon: Piedra Mesa H’ = 3,089; Buena Vista H’ = 3,005; La Cabaña H’ = 3,217). En general, se encontró homogeneidad para la zona de estudio (βw = 1,0) con bajo solape de familias de insectos (βr = 0,25).
El muestreo realizado con el método 2 permitió registrar a Lepidoptera, no encontrado con el método 1 y en total, 56 familias de insectos asociadas a los cultivos de pitaya amarilla en los predios productivos evaluados: Buena Vista n = 69, La Cabaña n = 63, Piedra Mesa n = 58 (Tabla 3).
Tabla 3 Familias de insectos encontrados mediante el método 2 en predios productivos de pitaya amarilla, Inzá, Cauca 2016-2017. n = número de individuos.
| Orden / Familia /Subfamilia/ Género | Buena Vista (n) | La Cabaña (n) | Piedra Mesa (n) |
|---|---|---|---|
| COLEOPTERA | |||
| Carabidae | 2 | 2 | |
| Chrysomelidae | 1 | ||
| Curculionidae | 1 | 1 | 1 |
| Dytiscidae | 2 | ||
| Hybosoridae | 1 | ||
| Lycidae | 1 | ||
| Nitidulidae | |||
| Carpophius sp. | 1 | 1 | |
| Glischrochilus sp. | 10 | ||
| Staphylinidae | 1 | 2 | |
| DIPTERA | |||
| Agromyzidae | 5 | ||
| Anthomyzidae | 3 | 1 | |
| Asteiidae | 10 | ||
| Cecidomyiidae | 1 | ||
| Ceratopogonidae | 1 | 1 | |
| Chloropidae | 2 | 3 | |
| Cryptochetidae | 1 | ||
| Dixidae | 1 | ||
| Dolichopodidae | 1 | 4 | 2 |
| Drosophilidae | 6 | 2 | |
| Empididae | 1 | ||
| Heleomyzidae | 1 | ||
| Lonchaeidae | 2 | ||
| Milichiidae | 7 | ||
| Mycetophilidae | 1 | ||
| Phoridae | 8 | ||
| Sciaridae | 1 | 6 | |
| Somatiidae | 1 | ||
| Sphaeroceridae | 5 | 1 | |
| Syrphidae | 9 | 3 | |
| Tipulidae | 1 | 1 | |
| HEMIPTERA | |||
| Alydidae | 1 | ||
| Aphididae | 1 | ||
| Aradidae | 2 | ||
| Berytidae | 1 | ||
| Cercopidae | 1 | ||
| Cicadellidae | |||
| Cicadellinae | 1 | ||
| Deltocephalinae | 1 | ||
| Megophthalminae | 2 | ||
| Ledrinae | |||
| Xerophloea sp. | 1 | ||
| Coreidae | 1 | ||
| Cydnidae | 3 | 6 | |
| Geocoridae | 1 | ||
| Lygaeidae | 4 | 1 | |
| Miridae | 1 | ||
| Pachygrontidae | 1 | ||
| Pentatomidae | 1 | ||
| Reduviidae | 2 | 1 | |
| Rhopalidae | 2 | ||
| Rhyparochromidae | 1 | 4 | |
| Thyreocoridae | 3 | ||
| HYMENOPTERA | |||
| Apidae | 1 | ||
| Braconidae | |||
| Braconinae | 1 | ||
| Opiinae | 1 | ||
| Formicidae | |||
| Acromyrmex sp. | 6 | 2 | |
| Ichneumonidae | |||
| Cryptinae | 1 | ||
| Pteromalidae | 1 | ||
| Sphecidae | 1 | ||
| LEPIDOPTERA | |||
| Geometridae | 1 | ||
| ORTHOPTERA | |||
| Acrididae | 4 | 7 | |
| Gryllidae | 1 | ||
| Tettigoniidae | 1 | ||
| Total | 69 | 63 | 58 |
Discusión
La pitaya amarilla constituye un cultivo introducido en Inzá-Cauca, que por su carácter perenne puede modificar de forma importante la dinámica poblacional y las interacciones ecológicas de las poblaciones de insectos, favoreciendo aquellas que puedan obtener mayor beneficio del nuevo recurso (Mack et al. 2000). La producción en pequeñas parcelas a modo de policultivos, con labranza mínima y un manejo fitosanitario basado en podas y remoción manual de arvenses en calles y plato, genera menor impacto en la biodiversidad insectil permitiendo la representatividad de familias de insectos benéficos para el sistema productivo, lo que promueve relaciones tróficas con tendencia al equilibrio de las poblaciones y sus distintas funciones en el sistema (Harvey et al. 2004; Schoonhoven 2005; Weyland y Zaccagnini 2008; Altieri y Nicholls 2007; Gutiérrez et al. 2007). No obstante, debido a que se encontró una correlación positiva entre variables abióticas y algunos grupos de insectos asociados al número de cladodios y el número de frutos de pitaya amarilla en los predios evaluados, debe enfatizarse en la arquitectura de las plantas durante su establecimiento, con miras a la regulación del microclima al interior de los cultivos (Pérez 1999; Medina y Kondo 2012; García 2016; Torres y García 2014; Kolb et al. 2016) como estrategia de prevención y manejo fitosanitario, particularmente en épocas de alta precipitación.
Dasiops saltans puede llegar a ocasionar pérdidas entre 40 - 80 % del rendimiento del cultivo de pitaya amarilla en un ciclo de floración, debido a que las larvas consumen el interior del botón floral (Vergara y Pérez 1988; Delgado et al. 2010). Aunque se encontraron especímenes del género Dasiops spp. en los predios con pitaya amarilla en Inzá, se sabe que el género está representado por lo menos por 45 especies asociadas a múltiples hospedantes (MacGowan 2017), por lo que no puede concluirse la presencia de D. saltans para esta zona de estudio.
No se encontró clorosis o punturas por daño asociado con las chinches del género Leptoglossus, pero las poblaciones deben ser vigiladas porque formas inmaduras de la familia Coreidae se asociaron con cladodios de la zona media de las plantas, como ha sido registrado para L. zonatus en otras zonas productoras de pitaya amarilla del país (Medina y Kondo 2012). A pesar de los hábitos polífagos de esta especie (Kondo et al. 2013a; Medina y Kondo 2012) y a la heterogeneidad del arreglo de los cultivos de pitaya en Inzá, debe realizarse seguimiento a las poblaciones del insecto, no solo por los daños directos ocasionados por sus hábitos de alimentación sino por el oportunismo que genera para la colonización de patógenos en la planta (Kondo et al. 2013a).
El país cuenta con información que permite el reconocimiento temprano de daños asociados a insectos en pitaya amarilla (Kondo et al. 2013a) por lo que en aras de mantener el equilibrio trófico en las poblaciones de artrópodos que redunde en la producción limpia de pitaya amarilla en Inzá, se requiere capacitación a los productores para el reconocimiento de las principales plagas. Por ejemplo, además de D. saltans, plaga clave, se ha registrado a Lonchaea longicornis Willinston, 1896 y Neosilba batesi (Curran, 1932) (Medina y Kondo 2012), de las cuales no se tiene definida su asociación con el cultivo. Aunque los tres géneros, que pertenecen a la familia Lonchaeidae no se diferencian a simple vista, la asociación con el daño en frutos, permitiría un sistema de alerta temprana para la mosca del botón floral de pitaya amarilla. Existe un importante riesgo de introducción y establecimiento de D. saltans en Inzá, proceso que puede favorecerse a través de transporte pasivo por los corredores de comercialización con zonas productoras de pitaya amarilla cercanas a Inzá en los departamentos del Huila y Valle del Cauca, donde la especie constituye el principal problema fitosanitario (Kondo et al. 2009; Delgado et al. 2010) y por las características ecológicas de la zona que pueden constituir un nicho apropiado para esta especie limitante (Patiño-Tira et al. 2013).
Los índices ecológicos revelaron una alta riqueza y diversidad de grupos insectiles, muy posiblemente por la contribución de familias con funciones benéficas como parasitoides (Braconidae e Ichneumonidae), depredadores (Syrphidae y Reduviidae), posibles cleptoparásitos de la familia Milichiidae, que podrían estar relacionadas a la presencia de Leptoglossus zonatus (Dallas, 1852) (Kondo et al. 2010) y polinizadores (Syrphidae, Nitidulidae y Apidae), particularmente en los predios Buena Vista y La Cabaña, caracterizados por un manejo agronómico no intensivo y alta diversidad vegetal presente con distintos cultivos acompañantes como yuca y maderables en el primer predio y con café, guayaba y guadua en el segundo. La promoción de prácticas agronómicas compatibles con la preservación de esta biodiversidad debe ser vigilada para evitar el aumento en la dominancia y consecuente reducción de la equitabilidad de grupos y desbalance trófico.
A pesar de no contar con criterios técnicos para el manejo de los cultivos comerciales de pitaya amarilla en Inzá, los agricultores establecieron una asociación para su comercialización nacional e internacional, por lo que se espera un negocio de largo aliento en la zona. Durante el desarrollo del estudio, los frutos presentaron sanidad fitosanitaria, excelentes cualidades organolépticas y gran tamaño (con peso promedio de 433 g). Debido a los retrasos en el desarrollo fenológico de las plantas, definido por estar localizados en zona marginal para el cultivo (Alarcón et al. 1996; García y Straube 1998; Wintgens 2004; Alvarado-Gaona et al. 2015), la cosecha está desfasada con respecto a la producción nacional, lo que favorece su comercialización. Aunado al impacto familiar, económico y social que ha tenido el cultivo de pitaya amarilla en Inzá-Cauca, se espera la ampliación de hectáreas sembradas, por lo que la información que aporte a la vigilancia entomológica es fundamental para un manejo integrado de plagas basado en criterios técnicos y de evaluación.
Colombia cuenta con importante información sobre el cultivo de pitaya amarilla (Ayala et al. 2009; Jiménez et al. 2012; Kondo et al. 2013b), por lo que debe establecerse un sistema de acompañamiento a productores que permita la adopción de sistemas de vigilancia fitosanitaria y de estrategias de manejo agronómico con criterio técnico, amigables con el ambiente y con perspectiva a largo plazo. La evaluación permanente de los sistemas productivos en Inzá-Cauca cobra especial importancia para alimentar los sistemas de alerta temprana para enfermedades y plagas, ya que una vez establecidos pueden generar pérdidas muy importantes. El equilibrio de las relaciones planta-insecto que se describen en el presente estudio deberían mantenerse a largo plazo, para lo cual la vigilancia entomológica regular es esencial.
Conclusiones
Las características del sistema de siembra tipo policultivo a pequeña escala para pitaya amarilla en Inzá, Cauca, fomentan la riqueza y diversidad de insectos con amplia representación de benéficos, base fundamental de relaciones tróficas complejas en beneficio del equilibrio del ecosistema. Aunque los hallazgos del estudio no revelaron la existencia de D. saltans y L. zonatus, descritas como insectos limitantes para pitaya amarilla para el país, se deben implementar sistemas de alerta temprana para reducir las posibilidades de ingreso y establecimiento de poblaciones de estas especies en la zona. Esta estrategia es fundamental para fortalecer el posicionamiento del municipio de Inzá como productor y exportador de la fruta, particularmente en épocas de baja producción en otras zonas del país.

