











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Inbreeding is defined as the mating of individuals whose relatedness between them is greater than the average degree of relationship existing in the population (Lush, 1945). Inbreeding seems to be a useful tool to improve the population by increasing the frequency of desirable genes, but the initial consequence of inbreeding is inbreeding depression. Inbreeding depression implies the reduction of additive genetic variance and phenotypic values for efficiency traits in inbred animals. Nowadays, this is a serious concern in animal breeding in which selection responses are maximized using animal model and Best Linear Unbiased Predictors (BLUP) of breeding value. The inbreeding level needs to be limited to maintain diversity at an acceptable level so as to improve the livestock programs (Van Wyk et al., 2009). Despite some positive inbreeding effect in selecting schemes, breeders are aware of its deleterious effects and try to avoid it. Inbreeding response is not the same for all animals. There is a wide range of variation in the literature regarding the estimation of inbreeding depression of growth traits (Mirza et al., 1999; Norberg and Sørensen, 2007; Van Wyk et al., 2009; Pedrosa et al., 2010; Ghavi Hossein-Zadeh, 2012; Eteqadi et al., 2014; Mokhtari et al., 2014).
Lori sheep is an important mutton fat-tailed Iranian breed, numbering approximately 2.5 million heads, mainly distributed in the Zagros Mountains of Lorestan province. It can also be found in some areas of Khuzestan, Kermansheh and Ilam provinces (Mohammadi et al., 2015). This breed has an average litter size of 1.25 lambs and its daily gain rate is higher compared to other Iranian sheep breeds. Average mature live weight in this breed is 62 Kg for ewes and 67 Kg for rams; milk yield ranges from 80 to 85 Kg in a 3-month period, approximatly, with annual greasy fleece weight between 1.5 to 2 Kg and 2 to 3 Kg in ewes and rams, respectively (Jalali Zonour, 2010).
There are no reports, to our knowledge, evaluating the inbreeding effects on growth traits in Lori lambs. This is the first study evaluating inbreeding depression and its trend on growth traits, to plan optimal designations to select programs. The aim of this study was to estimate and compare the inbreeding coefficient in Lori sheep and its effect on growth traits during a 10-year period.
Material and methods
Data and management of animals
Data and pedigree information were collected at the Lorestan Agricultural and Natural Resources Research Center, Khorramabad, Iran, from 2001 to 2010. The traits were birth weight (BW), weaning weight (WW), 6-month weight (6MW) and 9-month weight (9MW). The number of available records, mean, standard deviation, coefficient of variation and number of sires and dams are shown in Table 1.
Table 1 Descriptive statistics for growth traits of Iranian Lori sheep.
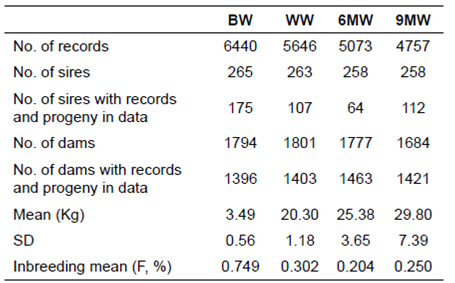
BW: birth weight; WW: weaning weight; 6MW: 6-month weight; 9MW:9-month weight; SD: standard deviation.
Iranian Lori sheep is a meat purpose breed. Maiden ewes were randomly mated to the rams for the first time at approximate 18 months of age. Ewes were usually kept in the flock for six parities (8 years of age) and culling was based on health and low performance. Selected rams were typically 3 to 4 years old and separately kept from ewes. Mating season initiated in early October until mid-November during the whole study. Consequently, lambing was restricted to one season, from mid-February until late March. Singlesire pens were used to allocate 20-25 ewes per ram. All lambs were weighed and ear tagged within 24 h of birth and pedigree and birth information were recorded. After lambing, lambs were separately kept in stalls with their dams for three days. Lambs were fed their mother milk together with good quality alfalfa hay ad libitum from 3 weeks of age until weaning, i.e. approximately 90 days. They were raised separately from older animals until 1 year of age. During this period, lambs were sold after weaning, generally due to poor growth or health issues. Weaned lambs were reared by sex group on natural pasture during spring and summer. In the fall, both lambs and ewes grazed on stubble fields of wheat, and barley, then kept indoors during the winter months with a ration composed of wheat barley straw and dry alfalfa, three times per day. The flock was fed a concentrate mixture containing alfalfa, barley grain, and wheat straw during the mating season as a flushing ration.
Statistical analysis
Data and pedigree file were statistically analyzed to assess the inbreeding depression of growth traits in Lori lamb and inbreeding trend. All of the phenotypic records exceeding 3.5 standard deviations above or below the mean were excluded. The GLM procedure of SAS (Version 9.2, SAS Institute Inc., Cary, NC, USA; 2009) was used to determine the fixed effects, which substantially affect the studied traits. Least square mean was estimated for each trait via the algorithm of Average Information Restricted Maximum Likelihood (AIREML) of the Wombat software (Meyer, University of New England Armidale, NSW 2351, Australia; 2007) by fitting six single trait animal models ignoring or including the additive direct, maternal genetic, and permanent environmental effects. The models included the fixed effects of sex (2 levels, male or female), litter size (2 levels, single or twin), age of ewe at lambing (6 levels, 2 to 7-year-old) and year of birth (10 levels, from 2001 to 2010) and also inbreeding categories and animal random effects. Age of lambs at 3, 6 and 9 months (in days) was considered as a linear covariate for WW, 6MW, and 9MW. Interactions amongst the mentioned fixed effects were not significant, so they were not considered in the final model.
The CFC (contribution, inbreeding and coancestry) software (Sargolzaei et al., Niigata University, Niigata 950-2181, Japan; 2006) was used to calculate the pedigree statistics and analysis of genetic structure of the population.
All the animals were classified in four categories based on the inbreeding coefficient of individual: the first category included non-inbred animals (F = 0); second, third, and fourth categories included inbred animals as 0<F ≤ 0.10, 0.10<F ≤ 0.20 and F>0.20, respectively. Inbreeding depression was estimated as the regression of growth traits (corrected for fixed effects) on the inbreeding coefficients of individual lamb using an animal model. Inbreeding trend was also estimated using the regression of the linear individual inbreeding on the year of birth via the Reg procedure of SAS (Version 9.2 SAS Institute Inc., Cary, NC, USA; 2009).
Results
The number of animals (in total), inbred animals, sires, dams, founders, and non-founders in the pedigree were 7,097, 2,126, 273, 1,955, 657, and 6,440, respectively. There were also 942 full sib groups in total, with average family size of 2.06 in the pedigree file. Table 2 illustrates the inbreeding categories. The maximum and minimum number of animals was in the first (F = 0) and fourth category (F>0.20). Inbreeding categories were only significant for BW and 6MW for all the traits.
Table 2 Inbreeding category distribution for growth traits in Lori lambs.
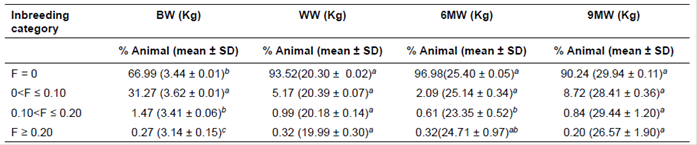
BW: birth weight; WW: weaning weight; 6MW: 6-month weight; 9MW: 9-month weight; SD: standard deviation. Means with the same superscript letter in each subclass within a column do not differ. Significant effect at p<0.05.
The different inbreeding categories for growth traits by lamb sex and litter size are presented in Tables 3 and 4, respectively. The BW of male and female lambs in different inbreeding categories were significant (p<0.05). Inbreeding categories for BW by litter size, for both male and female and 6MW were significantly different (p<0.05).
As it is shown in Table 3, considerable differences were observed in neither male lambs nor female lambs in the WW. Notwithstanding, non-significant difference amongst the four categories for 6MW female lambs; the males in the third category were lighter than others (p<0.05). On the other hand, the 9MW male lamb was non-significant amongst the four inbreeding categories. However, the 9MW female lamb within the fourth category was lighter than the others (p<0.05).
Although BW of single-born lambs within the second category was heavier than that of others, twins pertaining to the fourth category were lighter than those of others (p<0.01; Table 4). However, no significant differences were observed in WW of single and twin-born lambs. The 6MW of single born lambs in the third category was lighter than that of others, but twins within the fourth category were heavier than those of others (p<0.05). Although 9MW of single born lambs indicated no significant differences in the categories, the twins in the fourth category were heavier than others (p<0.05).
Regression coefficients of growth traits on inbreeding of Lori lambs for a change of 1% in inbreeding are presented in Table 5. By increasing the inbreeding level, 6MW and 9MW significantly decreased to -76.3 g and -77.4 g, respectively (p<0.05).
Table 3 Inbreeding category distribution by sex of Lori lambs.
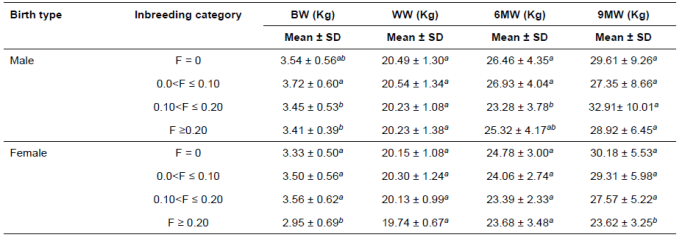
BW: birth weight; WW: weaning weight; 6MW: 6-month weight; 9MW: 9-month weight; SD: standard deviation.
Means with the same superscript letter in each subclass within a column do not differ. Significant effect at p<0.05.
Table 4 Inbreeding category distribution for growth traits by litter size of Lori lambs.
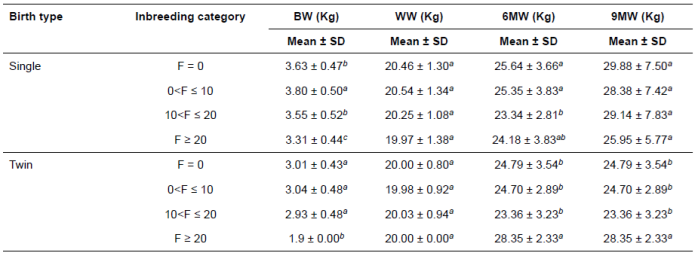
BW: birth weight; WW: weaning weight; 6MW: 6-month weight; 9MW: 9-month weight; SD: standard deviation.
Means with the same superscript letter in each subclass within a column do not differ. Significant effect at p <0.05.
Table 5 Regression coefficients for growth traits (in grams) on Lori lamb inbreeding for 1% change in inbreeding
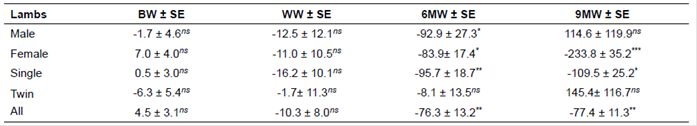
BW: birth weight; WW: weaning weight; 6MW: 6-month weight; 9MW: 9-month weight; SD: standard deviation. nsnon-significant, *Significant at p<0.05, **Significant at p<0.01, ***Significant at p<0.001.
Annual mean of inbreeding in whole and inbred populations of Lori lambs are illustrated in Figures 1 and 2, respectively.
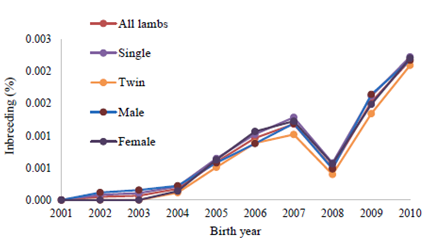
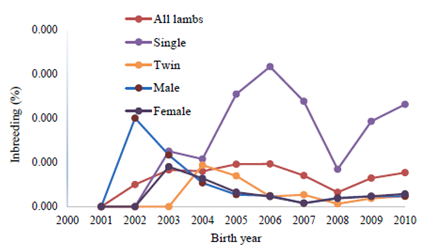
Figure 1 shows an overall increase in the inbreeding magnitude, irrespective of fluctuations in different years. In Figure 1, the inbreeding trend of growth traits shows an ascending change with mild fluctuations over a 10- year period. The mean of inbreeding decreased in year 2008 due to prevention of closed mating and selection of non-related animals to mate by the breeder. The maximum of inbreeding was observed in 2010 since inbred rams and ewes were mated with each other. The mean of inbreeding for twin lambs was 0 until 2003. In 2010, the mean of inbreeding was 2.19% for all of the lambs, showing an increased trend compared to the base year (2001). In total, there was an overall increase in the magnitude of inbreeding, irrespective of the fluctuations in different years (Figure 1).
Figure 2 shows the non-significant inbreeding trends in inbred population of Lori lambs. In Figure 2, considerable and irregular fluctuations were observed in annual inbreeding mean for male and single-born lambs; there were abrupt increases in the annual inbreeding mean for some years. Generally, relatively severe fluctuations were observed, especially for male and single-born lambs over a 10-year period, but inbreeding trends were insignificant.
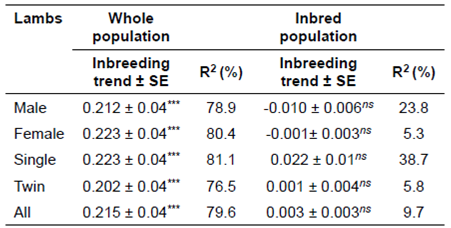
Inbreeding trends (% per year) in whole and inbred populations of Lori lambs are demonstrated in Table 6. In the current study, positive and significant inbreeding trends were only observed in whole population for male, female, single-born lambs, twinborn lambs, and all lambs. Inbreeding trend for all of the animals was 0.215 per year during the 10 years.
Discussion
According to Table 2, BW of lambs within the second category (0<F ≤ 0.10) was heavier than other categories (p<0.01); however, the differences were not remarkable between the first and third (0.10<F ≤ 0.20) categories. Consistent with the current results, Ghavi Hossein-Zadeh (2012) reported that BW of Moghani lambs pertaining to 0<F ≤ 0.10 category was heavier than that of lambs belonging to others. Nevertheless, there were no significant differences between seven inbreeding categories of BW in the study of Dorostkar et al. (2012). Concerning WW, it was not significant amongst four inbreeding categories. This finding was different from the results reported by Ghavi Hossein- Zadeh (2012), indicating that the WW of lambs within F>0.10 inbreeding was heavier than those of other categories (p<0.05). Moreover, heavier WW related to category F ≥ 0.25 reported by Dorostkar et al. (2012). The 6MW of lambs in the third category of inbreeding was lighter than those of the first and second categories (p<0.05); however, the difference was not significant between the third and fourth categories. Contrary to our findings, Dorostkar et al. (2012) reported heavier 6MW relating to F ≥ 0.25 in Moghani sheep. Also, there were no considerable differences amongst the four inbreeding categories on 9MW (p>0.05).
Inbreeding depression in males might be further reduced by sex chromosome hemizygosity, affecting the dominance of some mutations, as well as by the absence of sexual selection (Ebel and Phillips, 2016). The inbreeding coefficient is strongly determined by the two main factors, including depth and completeness of pedigree and intensity of selection. Intensity of selection is often increased by reproductive technologies focused on few superior animals, especially rams, and the application of advanced genetic evaluation methods.
Analyzing a total of 8,494 lamb records, Ghavi Hossein-Zadeh (2012) showed no significant differences in BW of single and twin born lambs, WW of single born lambs, 6MW, and 9MW of twins. The author found heavier WW of twin-born lambs in the F>0.10 compared to twins within the first and second categories (p<0.05). Moreover, the same researcher reported remarkable differences for 6MW and 9MW of single-born lambs in the F>0.10 with the lambs belonging to 0<F ≤ 0.10.
The regression coefficient of BW on inbreeding of lambs was positive and negligible (4.5 g, p>0.05; Table 5). Similarly, Dorostkar et al. (2012) found nonsignificant inbreeding depression per 1% increase in inbreeding for BW. Conversely, Van Wyk et al. (1993) observed a significant reduction of BW for Elsenburg Dormer lambs as 8 g per 1% increase in inbreeding. Khan et al. (1995) and Mirza et al. (1999) reported regression coefficients of -8 and -7 g, respectively. Norberg and Sørensen (2007) obtained significant reductions for BW, being 112, 82, and 88 g per 10% increase in Texel, Shropshire, and Oxford Down lamb inbreeding, respectively. Pedrosa et al. (2010) found a significant inbreeding depression for BW as 3.4 g per 10% increase in Santa Inês lamb inbreeding. In addition, the findings by Ghavi Hossein-Zadeh (2012), Mokhtari et al. (2014), and Eteqadi et al. (2014) were not similar to the findings of the present study. Ghavi Hossein-Zadeh (2012) reported that BW was affected by inbreeding of lambs being -9 g (p<0.001). The inbreeding depression of BW obtained as -7 g (p<0.05) using 6,082 Iran-Black sheep records analyzed by Mokhtari et al. (2014). Eteqadi et al. (2014) obtained inbreeding depression of BW as -5.5 g using 14,549 Guilan lamb records over an 18-year period.
The current results indicate that by increasing the inbreeding level, WW decreased, but not reached significance (-10.3 g, p>0.05). This finding was similar to the study by Mokhtari et al. (2014) and incompatible with the study by Dorostkar et al. (2012), being -37 and -291 g, respectively. Also, the WW of male, female, single and twin-born lambs was negative and non-significant decreased by 12.5 -11.0, -16.2, and -1.7 g owing to 1% increase in inbreeding, respectively. The studies by Khan et al. (1995), Mirza et al. (1999), and Akhtar et al. (2000) are in agreement with the present results. The WW of Rambouillet lamb was 21.11 ± 0.15 Kg; it decreased 10 g per 1% increase in inbreeding (Khan et al., 1995). A negligible reduction was found for WW of Lohi lambs as 6.9 g per 1% increase due to inbreeding (Mirza et al., 1999). Conversely, the inbreeding regression of WW was 99 g (p<0.01) in Elsenburg Dormer lambs (Van Wyk et al., 1993). In contrast, a non-significant decrease was observed as 6 g in WW of Horro lambs (Negussie et al., 2002). However, by increasing the inbreeding level, WW of Hissardale lambs insignificantly increased (Akhtar et al., 2000). Also, considering the sex of lamb and litter size, Eteqadi et al. (2014) reported a significant decline of 30.1, 28.9, 27.1, and 75.7 g in WW of male, female, single, and twin-born Guilan lambs per 1% increase in lamb inbreeding, respectively.
The reduction due to inbreeding for 6MW as 76.3 g found in the present study was higher than that observed by Eteqadi et al. (2014). Considering lamb sex, there were significant regression coefficients on male and female lamb inbreeding for change of 1% inbreeding by -92.9 and 83.9 g, respectively. Consequently, 6MW male and female lambs decreased to 92.9 and 83.9 g owing to 1% increase in inbreeding. Furthermore, a significant effect was obtained for litter size of 6MW; the regression coefficient of 6MW of single-born lambs on inbreeding was negative, being -95.7 (p<0.01). Hence, 6MW of single-born lambs decreased to 95.7 g due to 1% increase in inbreeding; on the other hand, 6MW of twins was non-significantly negative (-8.1 g, p>0.05). Inbreeding depression of 6MW was up to 79 g (Ghavi Hossein-Zadeh, 2012). Also, the coefficient of inbreeding regression was significantly different in terms of the lamb sex and litter size for 6MW (Ghavi Hossein-Zadeh, 2012). Inconsistent with the current findings, Pedrosa et al. (2010) and Eteqadi et al. (2014) obtained inbreeding depression of -204 and +20.1 g, respectively. Eteqadi et al. (2014) reported significant inbreeding reduction only for 6MW of twin lambs as -413.5 g, although inbreeding depressions were 20.9, 17.1, and 22.3 g for male, female, and single-born lambs, respectively. Moreover, Dorostkar et al. (2012) did not report any significant inbreeding effects on 6MW.
Inbreeding depression of 9MW was -77.4 g (p<0.01) per 1% increase in inbreeding coefficient. Also, 9MW of females and single-born lambs decreased up to 233.8 (p<0.001) and 109.5 g (p<0.05), respectively. Similar to the present findings, Eteqadi et al. (2014) reported regression coefficient as -45 g per 1% increase in 9MW of individual inbreeding; the non-significant inbreeding depression was -166, -71, and -25 g for twins, males, and females, respectively.
There are few published reports regarding the inbreeding effect on 9MW. There are several methodological and biological factors that determine the estimated inbreeding impact on performance traits. It is well known that both negative and positive effects exist. Hence, in a study undertaken on the population, «bad» and «good» inbreeding effects are mixed (Barczak et al., 2009).
Inbreeding significantly reduced post weaning growth traits, but it had no effect on the preweaning period. Thus, inbreeding showed no deleterious effects at early ages, but as the lambs grew older, the effect was remarkable.
Similar findings were reported by Dorostkar et al. (2012), as well. Notwithstanding, lower inbreeding trends were assessed by Norberg and Sørensen (2007), Van Wyk et al. (2009), Sheikhloo et al. (2012) and Rashedi Dehsahraei et al. (2013), for Danish, Elsenburg Dormer, Baluchi, and Lori-Bakhtiari sheep breeds, respectively. Concerning inbred population, inbreeding trends were only negative for male and female lambs being -0.010 and -0.001, respectively, which showed a non-significant level (p>0.05).
The difference between the present study and the others might be due to differences amongst breeds, variations of inbreeding level, genetic variation of the base population, population size, and ram to ewe ratio for mating, and different environmental and managing conditions. Also, responding to inbreeding banks on the genetic differences or differentials in levels of inbreeding density and differences between the animals such as age, sex, and litter size. Inbreeding is a special parameter in every population; being different in the various flocks is common. In general, these estimates of inbreeding trend provide a good prospective of the mating system for Lori sheep. Inbreeding level was low in the present study. The deleterious inbreeding effects were not obtained for BW and WW of this breed, but inbreeding depression was observed on 6MW and 9MW, especially for the 9MW females. Continuation of inbreeding trend rising in the whole population over the years warns that mating shall be planned more carefully in the future to avoid mating of close relatives. Furthermore, using the designed mating system could be an appropriate approach to maintain the inbreeding level under control in the present population.
In this study, positive and significant inbreeding trends were 0.212, 0.223, 0.223, and 0.202 in the whole population for male, female, single born, and twin born lambs, respectively (Table 6). A lower inbreeding trend was observed by Huby et al. (2003) and Van Wyk et al. (2009). However, a higher inbreeding trend (0.40) was reported by Norberg and Sørensen (2007). The mean of inbreeding for the whole population (0.69) was lower than that in previous reports (Norberg and Sørenson, 2007; Van Wyk et al., 2009; Tahmoorespur and Sheikhloo, 2011; Rashedi Dehsahraei et al., 2013).
Acknowledgements
We would like to thank Parviz Mohammadi Ghalaei, Pharm. D., for providing Spanish and Portuguese versions of the Abstract, and Mr Abouzar Mohammadi, Kashan University of Medical Sciences and Health Services, Kashan and Dr Hassan Khajavy, University of Bojnord, for proofreading our manuscript. Also, we acknowledge the staff of the Agricultural and Natural Resources Research Center of Lorestan province for providing the data for the study.

