











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
El género Kalanchoe pertenece a la familia crasulácea (crasa o suculenta), tiene alrededor de 125 especies descritas. Se encuentran principalmente en África, Madagascar, América tropical y Australia (Abdel-Raouf, 2012). Actualmente está distribuido por todo el mundo (Mora-Pérez y Hernández-Medel, 2016). Son comúnmente usadas en la medicina natural y farmacéutica, por producir metabolitos secundarios ricos en alcaloides, triterpenos, glucósidos, flavonoides, esteroides (bufadienolides) y lípidos (Majaz et al., 2011), que ayudan a tratar problemas de salud como la bronquitis, quemaduras, dolores de oído, picaduras de insectos, infecciones respiratorias, tuberculosis, problemas de la piel, infecciones bacterianas y fúngicas, problemas intestinales, cáncer (linfoma), epilepsia, varicela, tratamientos clínicos del corazón, entre otros (Okwu y Nnamdi, 2011; Tkalec et al., 2012; Salahdeen y Yemitan, 2006; Kolodziejczyk-Czepas et al., 2017).
Kalanchoe daigremontiana Raym.-Hamet & H. Perrier (espinazo del diablo) es una hierba suculenta, nativa de zonas semiáridas de Madagascar (Herrera, 2011). Su característica principal es producir pequeñas plántulas o hijuelos, que nacen entre los dientes del margen aserrado del foliolo (Garcês et al., 2007; Puertas et al., 2014). Produce sustancias bioactivas con propiedades antioxidantes, antitumorales, antihistamínicas, antiinflamatorias e inmunomoduladora (Tkalec et al., 2012). Además, se ha utilizado tradicionalmente para la cura de heridas y como relajante o calmante, se ha estudiado su eficacia para controlar células cancerígenas de pulmón, cuello de útero y en carcinoma hepatocelular, debido a la presencia de compuestos bufadienolides en hojas y raíces (Saz-Peiró y Tejero-Lainez, 2016).
El uso de K. daigremontiana como planta medicinal se ha incrementado específicamente en los tratamientos del cáncer, en donde se ve como una alternativa viable, a las moléculas químicas que produce (Puertas et al., 2014; Majaz et al., 2011). Sin embargo, la extracción y purificación de dichos compuestos, requiere una gran cantidad de material vegetal para obtener cantidades significativas del compuesto de interés (Foley y Moore, 2005). Por lo tanto, es necesario desarrollar un sistema o metodología de cultivo, que garantice una producción sostenible de estos metabolitos (Arias et al., 2009). Esto ha desarrollado alternativas para la obtención de mayores volúmenes de producción, a través de programas de selección y mejoramiento genético (Quintero et al., 2009; Sattler et al., 2016). La utilización de la colchicina ha sido exitosa, demostrando un mayor crecimiento y aumento en la producción de biomasa en diferentes plantas (Urwin, 2014), ya que permite un incremento de sus partes vegetativas ocasionadas por la poliploidía (Sánchez y Matos, 2012; Matos, 2014; Sadat et al., 2017). La colchicina es probablemente, el producto químico más ampliamente utilizado para la inducción de poliploidía en plantas (Urwin, 2014), debido a que es un agente inhibidor eficaz de la mitosis, que impide la polimerización de la tubulina durante la formación del huso mitótico, lo que causa el bloqueo de la metafase y dispersión de los cromosomas en la célula (Planchais et al., 2000). La ventaja más importante de la poliploidía, es que las plantas suelen tener mejor rendimiento y características morfológicas, como la altura y el tamaño de los órganos vegetales (Hannweg et al., 2016), y el aumento de la biomasa en general (Urwin, 2014).
Para identificar plantas poliploides, se realizan métodos indirectos como caracteres morfológicos (Sánchez y Matos, 2012), métodos citológicos midiendo el tamaño y densidad estomática, el número de cloroplastos en las células guardas de los estomas (Standring et al., 1990; Choque et al., 2007; Orrillo y Bonierbale, 2009; Sadat et al., 2017) y mediante el conteo de cromosomas (Poggio et al., 2008; Matos, 2014). Por lo anterior, teniendo en cuenta la importancia medicinal, y que no se han realizado investigaciones sobre la utilización de la colchicina en K. daigremontiana, el siguiente estudio tiene como objetivo evaluar el efecto de diferentes concentraciones, y tiempo de exposición de colchicina sobre la morfología foliar y los estomas de K. daigremontiana.
MATERIALES Y MÉTODOS
Material vegetal y condiciones del cultivo
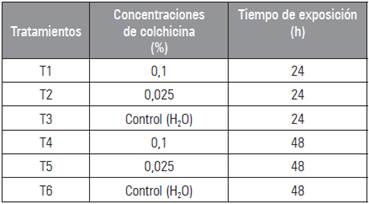
Las plántulas (hijuelos) de Kalanchoe daigremontiana, se recolectaron de una planta madre, adquirida en el municipio de Villa del Rosario de Norte de Santander (Colombia). Se tomaron 60 hijuelos, con una altura promedio de 1 cm y un sistema radicular desarrollado (Imery y Cequea, 2001). Posteriormente, en cámara oscura las plántulas fueron sometidas a diferentes concentraciones (0,025 y 0,1%: p/v) y tiempos de exposición (24 y 48 h) con colchicina, tomando como control el agua destilada (Tab. 1). Seguidamente, fueron sembrados en bolsas de polietileno de 1 kg en un sustrato compuesto de lombrinaza, cascarilla de arroz y suelo de perfil arcilloso en proporción 1:1:1, bajo condiciones climáticas de la zona, caracterizada por ser un bosque seco tropical a 320 msnm, temperatura promedio de 28,3 °C, humedad relativa del 60% y precipitación promedio anual de 450 mm año-1 (Ramírez, 2007). El estudio se realizó en el laboratorio de fitomejoramiento de la Universidad Francisco de Paula Santander.
Estudio morfológico
Para el análisis de la morfología se tuvieron en cuenta las plantas tratadas y los hijuelos que se formaron en los tratamientos. Se calculó el volumen foliar (V) empleando la fórmula empleada por Matos (2014):
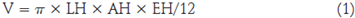
consideraron aspectos como altura de la planta (AP), largo de las hojas (LH), ancho de las hojas (AH), número de hojas (NH), espesor (EH). Las anteriores variables fueron tomadas con una regla milimetrada a excepción del EH, para este se utilizó un calibrador pie de rey. Las mediciones se realizaron cada 15 d, durante 16 semanas después de la siembra. Los hijuelos formados se evaluaron a las 8 semanas.
Características de los estomas
Para las características estomáticas como el ancho, el largo y el número de cloroplasto, se implementó y se modificó la metodología realizada por Ganem et al. (2014), donde se utilizó cinta adhesiva transparente, para retirar la epidermis de la zona abaxial de la parte central de la hoja (1 cm2 aproximadamente). La cinta se fijó en un porta objeto, posteriormente se colocó la muestra al microscopio compuesto LEICA modelo DME 500 (Leica Microsystems, Buffalo, NY, USA). Al elegir los estomas, se enfocó con el objetivo 100X, esto se repitió 10 veces por accesión. Las fotos obtenidas se analizaron con el software Infinity Analyze 6.5 (Release 5.0.3, Lumenera, Ottawa, Canada).
En la determinación del índice estomático (IE), se contó el número de estomas por célula epidérmica donde el campo de observación contaba con un aumento total de 400X. El IE se calculó utilizando la fórmula implementada por Wilkinson (1979):
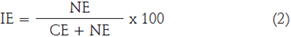
donde NE es el número de estomas por campo de observación y CE número de células epidérmicas en el campo visual. Se hicieron 10 réplicas por tratamiento.
Diseño experimental y análisis estadístico
Consistió en un diseño factorial 3x2 completamente al azar (tres concentraciones y dos tiempos de exposición), con 10 repeticiones por tratamiento, para un total de 60 unidades experimentales. Los datos obtenidos en el análisis morfológico y estomático, se evaluaron mediante un análisis de varianza (ANOVA). Posteriormente, las medias se compararon utilizando la prueba de rangos múltiples de HSD (Diferencia Significativa Honesta) de Tukey, para determinar las diferencias significativas a un nivel de P≤0,05 (Tukey, 1953). Para el análisis estadístico se utilizó el software Statgraphic Centurión v. 16.0 (Statpoint, Herndon, VA) y para graficar se implementó el software KyploT v. 2.0 (Kyence Lab. Tokyo).
RESULTADOS Y DISCUSIÓN
Estudio morfológico
De acuerdo a la evaluación de las características morfológicas en K. daigremontiana, se encontró variación en los tratamientos implementados (Figs. 1 y 2). En la valoración final (semana 16: Tab. 2), se observó diferencias estadísticamente significativas en el tratamiento con colchicina 0,025% por 24 h (T2) causando un mayor incremento en el largo de la hoja (LH), ancho de la hoja (AH) y volumen foliar (VF), con respecto a los tratamientos control y las demás plantas tratadas, de igual manera el tratamiento 0,025% por 48 h (T5) mostró diferencias significativas en el espesor de la hoja (EH), en la altura de la planta (AP) y número de hijuelos (NHT; Tab. 2; Fig. 2). Del mismo modo, no se observaron diferencias estadísticamente significativas entre los tratamientos de control (T3, 24 h y T6, 48 h), con medias muy por debajo del resto de los tratamientos en el análisis morfológico (Tab. 2). Es de destacar que solo en los tratamientos 2, 4 y 5 formaron hijuelos en el transcurso de las 16 semanas (Tab. 3 y Fig. 2B). Según lo anterior, los tratamientos T2 y T5 (colchicina 0,025% a 24 y 48 h, respectivamente de exposición) fueron más efectivos, ya que el incremento en la morfología foliar fue más rápido y mayor que los demás tratamientos (Tab. 2 y Figs. 2A, B, C y D). Sin embargo, en los hijuelos del tratamiento 0,025% por 48 h (T5) se encontró mejor desarrollo morfológico y volumen foliar (Tab. 3). Estos resultados difirieren de los encontrados por Sánchez y Matos (2012) y Matos (2014), en donde encontraron que el tratamiento con colchicina 0,10% por 48 h en Aloe vera causó incrementos significativos en altura de las plantas (AP), longitud de las hojas (LH) y volumen foliar de los hijuelos. En una investigación realizada por Sadat et al. (2017) el tratamiento en el cual se halló mayor altura de la planta en Trachyspermum ammi aplicando colchicina, fue al 0,1% durante 6 h. En cuanto a la longitud de la hoja el mejor tratamiento fue de 0,5% por 48 h de colchicina. Además, la aplicación de colchicina generó diferentes tamaños y hojas en las plantas medicinales de Trachyspermum ammi y Tetradenia riparia, algo similar ocurrió en las hojas de K. daigremontiana (Fig. 2D). Según Hannweg et al. (2016), la ventaja más importante de la poliploidía inducida es que las plantas obtenidas suelen tener mejor rendimiento y características morfológicas tales como altura, mayor tamaño de hojas, rizoma o raíz. Asimismo, incrementa la biomasa, la capacidad foto-sintética, el tamaño de los frutos y semillas (Urwin, 2014). En este estudio, se registraron diferencias altamente significativas en plantas tratadas con colchicina en cuanto a la morfología foliar se pudo apreciar un cambio en las proporciones de las hojas de acuerdo a cada tratamiento esto se puede constatar comparando las medias de variables como largo de hoja (LH) y ancho de la hoja (AH) entre los tratamientos T2 y T5, tomando estos como referencias puesto que presentaron mayor desarrollo (Tabs. 2 y 3; Figs. 1 y 2). Lo anterior representa un indicio muy fuerte de poliploidía (Sadat et al., 2017; Hannweg et al., 2016; Urwin, 2014). Sánchez y Matos (2012), señalaron que en plantas de A. vera tratadas con colchicina al 0,05% y 0,10% durante 48 h se logró inducir cambios a nivel morfológico, citogenético y anatómicos. Según Matos (2014), si se expone a mayor tiempo las plantas de A. vera con colchicina hasta 48 h, estas inducen cambios morfológicos en las plantas. Debido a que aumenta el número de células meristemáticas en contacto con colchicina, favoreciendo la penetración del alcaloide a las capas celulares durante la actividad mitótica de la misma.
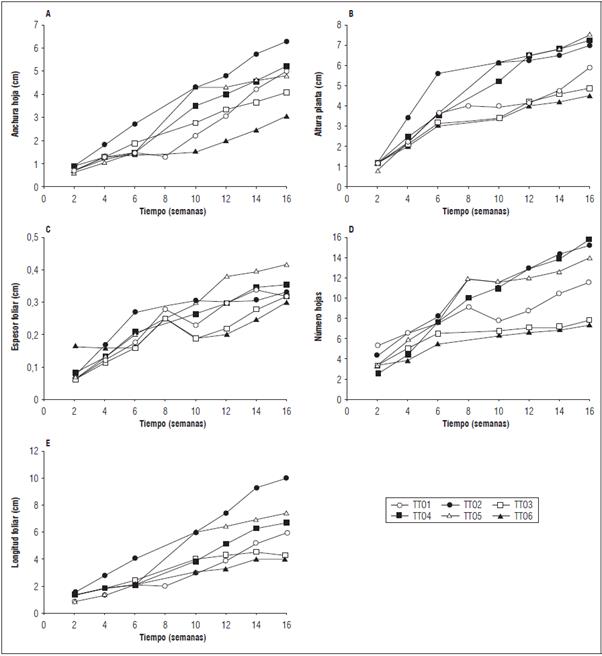
Figura 1 Estudio morfológico de Kalanchoe daigremontiana. A. Anchura de la hoja (AH); B. Altura de la planta (AP); C. Espesor foliar (EH); D. Numero de hojas (NH); E. Longitud foliar (LH). T1: 0,1% de colchicina por 24 h; T2: 0,025% de colchicina por 24 h; T3: Control (H2O) por 24 h; T4: 0,1% de colchicina por 48 h; T5: 0,025% de colchicina por 48; T6: Control (H2O) por 48 h.

Figura 2 Efecto de la colchicina sobre la morfología foliar de Kalanchoe daigremontiana. A. Comparación del T2 con T3 a las 8 semanas; B. Hijuelos formados; C. Comparación de los tratamientos en la semana 16; D. Hojas a las 16 semanas. T1: 0,1% de colchicina por 24 h; T2: 0,025% de colchicina por 24 h; T3: Control (H2O) por 24 h; T4: 0,1% de colchicina por _48 h; T5: 0,025% de colchicina por 48; T6: Control (H2O) por 48 h.
Tabla 2 Efecto de la colchicina sobre la morfología foliar de Kalanchoe daigremontiana. Evaluación realizada a las 16 semanas después de la siembra.
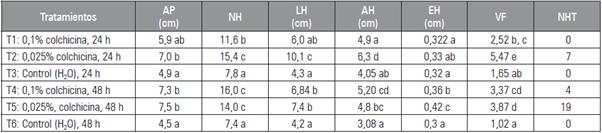
AP: altura de la planta; NH: número de hojas; LH: longitud foliar; AH: anchura de la hoja; EH: espesor foliar; VF: volumen foliar; NHT: número de hijuelos por tratamiento.
Promedios con letras distintas, en cada columna, indican diferencia significativa según la prueba de Tukey HSD (P≤0,05).
Tabla 3 Efecto de la colchicina sobre la morfología foliar de los hijuelos producidos por Kalanchoe daigremontiana. Evaluación realizada a las 8 semanas.
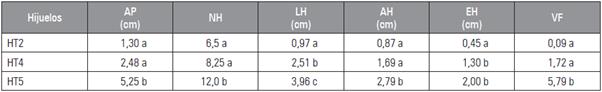
HT2: hijuelos del tratamiento con 0,025% de colchicina por 24 h; HT4: hijuelos del tratamiento con 0,1% de colchicina por 48 h; HT5: hijuelos del tratamiento 0,025% de colchicina por 48 h.
Abreviaturas de las variables como en tabla 2.
Promedios con letras distintas, en cada columna, indican diferencia significativa según la prueba de Tukey HSD (P≤0,05).
Características de los estomas
En las características estomáticas la concentración de colchicina de 0,025% y 0,10%, en los tiempos de exposición (24 y 48 h), tuvieron un aumento en el tamaño de los estomas (largo y ancho), la cantidad de cloroplastos y en las células guardas de los estomas (Figs. 3 y 4; Tab. 4).
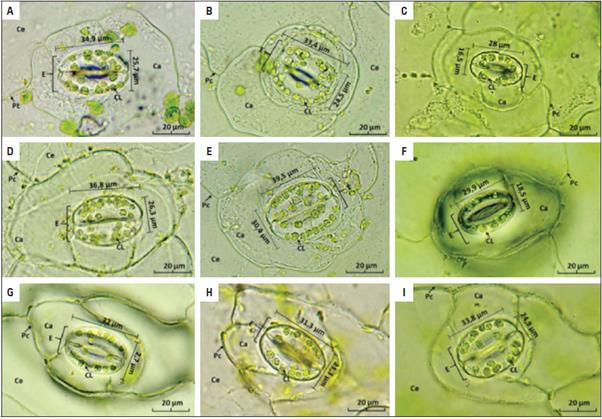
Figura 3 Características de los estomas anisocíticos de la epidermis abaxial de la lámina foliar de Kalanchoe daigremontiana. Los tratamientos correspondieron a T1: 0,1% de colchicina por 24 h; T2: 0,025% de colchicina por 24 h; T3: Control (H2O) por 24 h; T4: 0,1% de colchicina por 48 h; T5: 0,025% de colchicina por 48; T6: Control (H2O) por 48 h. A. Estoma del T1 con 20 cloroplastos (CL); B. Estoma del T2 con 30 CL; C. Estoma del T3 con 10 CL; D. Estoma del T4 con 25 CL; E. Estoma del T5 con 27 CL; F. Estoma del T6 con 11 CL; G. Estoma del hijuelo del tratamiento 2 (HT2) con 18 CL; H. Estoma del HT4 con 15 CL; I. Estoma del HT5 con 17 CL. Ca: célula acompañante; Ce: célula epidérmica; CL: cloroplasto; E: estoma; Pc: pared celular. Aumento total 1.000x. Escala de la barra = 20 µm.
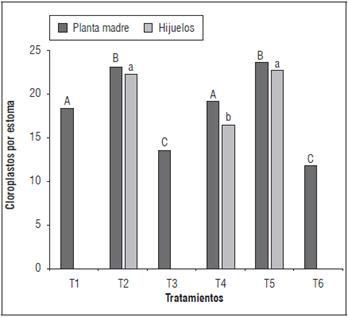
Figura 4 Número de cloroplastos por estoma en Kalanchoe daigremontiana tratadas con colchicina a diferentes concentraciones y tiempos de exposición. T1: 0,1% de colchicina por 24 h; T2: 0,025% de colchicina por 24 h; T3: Control (H2O) por 24 h; T4: 0,1% de colchicina por 48 h; T5: 0,025% de colchicina por 48; T6: Control (H2O) por 48 h. Promedios con letras distintas, para cada planta madre e hijuelos independientes, indican diferencia significativa según la prueba de Tukey HSD (P≤0,05).
Tabla 4 Características estomáticas de Kalanchoe daigremontiana tratadas con colchicina.
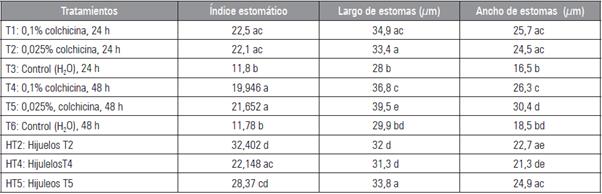
Promedios con letras distintas, en cada columna, indican diferencia significativa según la prueba de Tukey HSD (P≤0,05).
Estos resultados coinciden con los observados por Sadat et al. (2017) y Tambong et al. (1998) en donde mostraron coeficientes de correlación positivos y significativos, entre el largo y ancho de los estomas y el número de cloroplastos en las células guardas de los estomas. La morfología de los estomas es determinante para controlar la pérdida de agua, lo que posibilita la adaptación de muchas especies vegetales a condiciones ambientales muy diversas (Hetherington y Woodward, 2003). La modificación de las características de los estomas podría ejercer una tarea sustancial en el proceso de aclimatación de una especie al ambiente. Investigaciones anteriores en plantas tetraploides inducidas con colchicina, se ha observado repetidamente que el largo y ancho, como el número de estomas y el número de cloroplastos existentes dentro de las células guardas, cambian significativamente en caso de duplicación del cromosoma, en comparación con el estado diploide (Beck et al., 2003; Urwin et al., 2007). Los cloroplastos son organelos donde se realiza el proceso de la fotosíntesis, por tanto, a mayor número de estos es probable que haya una mayor actividad foto-sintética en la planta, sintetizando mayor contenido de materia orgánica (Gordillo et al., 2008). Sadat et al. (2017) en una comparación de las características de los estomas en plantas tetraploides y diploides de Trachyspermum ammi, mostraron el aumento del largo y ancho de los estomas en plantas tetraploides inducidas con colchicina, en contraste con sus pariente diploides. Igualmente, Zhang et al. (2008), encontraron un mayor número de estomas en plantas tetraploides que en diploides en Phlox subulata. En otro estudio, Aryavand et al. (2003) concluyeron que el tamaño y el número de los estomas pueden utilizarse para distinguir las plantas tetraploides de las hexaploides en Aegilops neglecta. En otras especies, como Vitis vinifera, Triticum monococcum, Triticum durum y Triticum aestivum se obtuvieron diferencias significativas en las características estomáticas en plantas poliploides y diploides (Yang et al., 2006; Khazaei et al., 2010).
En el T5 y T2 se hallaron la mayor cantidad de cloroplastos, con una media de 23,6 y 23,0 cloroplastos, respectivamente (Figs. 3 y 4). A diferencia, en los tratamientos control, el cual se observó la menor cantidad de cloroplastos (T3: 13,5 y T6: 11,5). Respecto a los hijuelos de los tratamientos T2 y T5, no presentaron diferencias significativas (HT2: 22,3 y HT5: 22,8). Standring (1990), planteó que el uso de variaciones en el número de cloroplastos de las células oclusivas del estoma, es una manera segura de predecir el nivel de ploidía en algunos o diferentes géneros. Se debe agregar que las modificaciones de los niveles de ploidía presentan una relación con los cloroplastos; si se modifica el número de juegos de cromosomas, estos pueden modificar la cantidad de cloroplastos que presentan las células guardas de los estomas (Orrillo y Bonierbale, 2009).
Con respecto al índice estomático, se observó en el campo visual que los tratamientos T1 y T2 (Figs. 5A y B), una menor cantidad de células epidérmicas dándose un aumento en el número de estomas, posiblemente al aumento en el largo y ancho del estoma. A diferencia del T3 (Fig. 5C) en la cual existe una mayor cantidad de células epidérmicas, con menor número de estomas. Estos hallazgos difirieren de los encontrados por Sadat et al. (2017), en cuanto a la densidad estomática, ya que los investigadores demostraron una disminución en el número de estomas por campo visual para plantas tetraploides con respecto a las plantas dipolides de Trachyspermum ammi. Según Foschi et al. (2013) es posible determinar el nivel de ploidía, a través del tamaño y densidad de los estomas, siendo la medición estomática un aspecto muy importante para determinar los diferentes niveles de ploidía en la mayoría de las especies.

Figura 5 Epidermis abaxial de la lámina foliar de Kalanchoe daigremontiana tratada con colchicina. A. Índice estomático a 0,1% de colchicina por 24 h (T1); B. Índice estomático a 0,025% de colchicina por 24 h (T2); C. Índice estomático del control (H2O) por 24 h (T3). Aumento total 400x. Escala de la barra = 40 µm. Ce: célula epidérmica; Es: Estomas; Pc: pared celular.
CONCLUSIONES
La aplicación de colchicina a Kalenchoe daigremontiana al 0,025% por 24 y 48 h (tratamientos T2 y T5), mejora los rasgos morfológicos como altura de la planta, largo, ancho, y número hojas, espesor y volumen foliar y las características estomáticas. Lo que demuestra que el uso de la colchicina favorece el crecimiento en poco tiempo y aumenta la biomasa en la planta K. daigremontiana. Es importante comparar el análisis morfológico y estomático con el conteo de cromosomas y el análisis de citometría de flujo, para determinar el nivel de ploidía (Sadat et al., 2017). Por tal motivo, se plantea la necesidad de realizar estudios adicionales en donde se pueda determinar específicamente el nivel de ploidía en la planta medicinal de K. daigremontiana.


