











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkLa enfermedad de Chagas es una infección parasitaria crónica inicialmente considerada endémica de América, en donde actualmente se estima que entre 9 y 11 millones de personas la padecen, y que entre 60 y 120 millones se encuentran en riesgo de adquirirla (1). Debido a los cambios sociodemográficos, actualmente se la categoriza como una enfermedad global (2).
Causada por el protozoo Trypanosoma cruzi, la enfermedad de Chagas involucra a numerosos reservorios animales domésticos y selváticos. Se transmite al hombre principalmente porlas heces de insectos hematófagos infectados pertenecientes a la subfamilia Triatominae (Heteroptera: Reduviidae), y de madres a hijos, así como por transfusiones y trasplante de tejidos no controlados y por la ingestión de alimentos contaminados, principalmente jugos de frutas (3). Si a ello se añaden los recientes flujos migratorios, las rutas comerciales de los países emergentes y la modernización de los métodos de transporte, es fácil comprender su difícil control y el efecto de la globalización en su distribución (2). No obstante, la enfermedadde Chagas es una de las 17 enfermedades infecciosas desatendidas (4).
Una de las principales estrategias para reducir el número de nuevos enfermos es eliminar de las zonas endémicas a los vectores responsablesde transmitir el parásito (5). En Argentina, el principal vector transmisor de T. cruzi es Triatoma infestans. Su capacidad de adaptarse al ambiente humano y colonizar densamente el domicilio y el peridomicilio, y ciertos patrones fisiológicos como la cantidad de sangre que ingiere y la capacidad de excretar parte del contenido del tubo digestivo mientras se está alimentando, entre otros, hacen de esta especie un vector muy eficaz en la transmisión del parásito (6).
Además de la voluntad política de eliminar los principales vectores deT. cruzi de las regiones endémicas mediante iniciativas como la del Cono Sur (Incosur), la del Pacto Andino (IPA), la centroamericana (IPCA) y la de la Amazonia (AMCHA), el reemplazo del lindano (gammahexaclorociclohexano) por piretroides sintéticos a mediados de los 80 marcó un hito en el control del vector (1). Los piretroides demostraron ser moléculas de gran eficacia en T. infestans, pues poseen un gran poder residual, son bien aceptados por los habitantes de los hogares fumigados y no sufren bioacumulación, como ocurre con los organoclorados utilizados previamente (7).
En 1997, Uruguay fue el primer país en el cual se pudo certificar la interrupción vectorial de la enfermedad de Chagas (8). Chile y Brasil consiguieron dicha certificación en 1999 y 2006, respectivamente (9), en tanto que hasta el día de hoy, Argentina, Perú y Paraguay solo han logrado certificar la interrupción de la transmisión vectorial en ciertas zonas de su territorio.
Si bien en un comienzo los piretroides utilizados en las campañas de fumigación tuvieron gran eficacia en la eliminación deT. infestans en los domicilios, dos situaciones condicionaron su uso masivo en distintas poblaciones del vector: la poca eficacia detectada frente a poblaciones de triatominos habitantes del peridomicilio (10), y la aparición de resistencia y tolerancia registradas en diferentes localidades de la región (11).
Ante estas limitaciones, surgió la urgente necesidad de implementar estrategias alternativas y complementarias de control. Una de ellas postula la administración de diversos principios químicos con actividad frente a los triatominos en animales quehabitan el domicilio y el peridomicilio. Muchos de estos animales (perros, gallinas, cabras, etc.), además de representar una fuente de alimentación para los triatominos, se comportan como reservorios de T. cruzi (12).
Aunque el objetivo principal de los estudios es hallar moléculas que puedan provocar la muerte de los triatominos al ser incorporadas en estos, no es el único, ya que también se estudian moléculas con capacidad de interferir en sus patrones de alimentación y defecación como buenos blancos a la hora de encontrar alternativas para su control.
En un estudio previo, nuestro equipo demostró mediante pruebas in vitro la actividad tóxica de tres antiparasitarios muy utilizadosen medicina veterinaria, la doramectina, la eprinomectinay laivermectina, en ninfas de quinto estadio de T. infestans (13). En dicho estudio, se encontró que la ingestión de las tres moléculas provocaba diferentes grados de intoxicación en las ninfas.
La doramectina exhibió una mayor potencia contra los insectos y mantuvo su actividad anti-T. infestans,incluso en las concentraciones más bajas evaluadas, del orden de los 0,4 ng/ml.
Los tres compuestos estudiados pertenecen al grupo de las avermectinas, las cuales, conjuntamente con las milbemicinas (moxidectina), se conocen como endectocidas, término que hace referencia a su espectro de acción contra endoparásitos y ecto-parásitos. Lo interesante de estas moléculas es que ejercen su acción sobre los parásitos de manera diferente a la de los piretroides.
Cuando se usa un endectocida en un parásito sensible, se produce un aumento en el flujo de iones cloruro hacia el interior de las células neuronales, evento que se atribuye a los canales de cloro regulados por el glutamato, lo cual desencadena una hiperpolarización de la neurona (14). Este mecanismo da lugar a una disminución de la motilidad del parásito, pues afecta su musculatura somática y le impide mantenerse en su hábitat, en tanto que el efecto sobre la musculatura de la bomba faríngea interfiere profundamente en la alimentación. La combinación de estos dos mecanismos produce la muerte del parásito (15). Los piretroides, en cambio, aumentan el tiempo de apertura de los canales de sodio y, por lo tanto, desencadenan descargas repetitivas en las neuronas, dando lugar a una parálisis espástica del insecto (15,16). El hecho de que la diana molecular de ambas familias de agentes sea distinta no es un dato deleznable, pues son varios los casos en los que los endectocidas se han convertido en una alternativa para eliminar a insectos vectores que desarrollaron resistencia a los piretroides (17).
En diversos trabajos se ha comprobado la estrecha relación entre distintos roedores y diferentes triatominos en todo el continente americano (18-20), ya que ciertos roedores, además de representar una buena fuente de alimento para los triatominos, también pueden actuar como huéspedes deT. cruzi (20), lo cual resalta su importancia en la epidemiología de la enfermedad de Chagas.
El objetivo del presente estudio fue evaluar si el efecto tóxico, demostrado in vitro de la ivermectina, la doramectina y la eprinomectina en T. infestans persistía en un modeloin vivo de roedor (ratas Wistar). El efecto tóxico de lasmoléculas se evaluó con base en tres variables: la cantidad de sangre ingerida, el volumen de excreciones y el porcentaje de mortalidad en los insectos.
Materiales y métodos
Animales experimentales
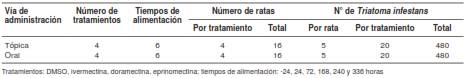
Se utilizaron 32 ratas Wistar de ambos sexos y de dos meses de edad, aproximadamente. Los roedores se dividieron al azar en dos grandes grupos según la vía de administración de los endectocidas, y cada uno de estos grupos se sub-dividió en cuatro grupos de cuatro individuos (dos machos y dos hembras) cada uno (cuadro 1).
Cuadro 1 Distribución del material biológico utilizado (ninfas de Triatoma infestans y ratas Wistar) según los diferentes tratamientos evaluados
Cada uno de los grupos de roedores se alojó en una jaula debidamente identificada. Los animales se mantuvieron en condiciones ambientales controladas y constantes (22 ± 2 oC, humedad relativa, HR, de 70 a 80 % y fotoperiodo de 12 horas), y se les suministraron agua y alimento ad libitum.
Formulaciones experimentales
Se prepararon soluciones de ivermectina, doramectina y eprinomectina al 0,5 % en dimetilsulfóxido (DMSO), todascon los ingredientes activos de grado técnico: ivermectina (90 %), eprinomectina (90,7 %) y doramectina (94,8 %) (Romikin, S.A., Argentina). Para el cálculo de la concentración se consideró la pureza de cada sustancia, con el fin de ajustar la masa del ingrediente activo que debía pesarse e igualar así las concentraciones de los tres endectocidas.
Administración de las formulaciones experimentales
Cada grupo de roedores recibió ivermectina, doramectina, eprinomectina o DMSO por vía oral mediante sonda buco-esofágica, o por vía tópica mediante su aplicación puntual a nivel dorsal (spot-on). Los endectocidas se administraron a razón de 600 µg/kg de peso vivo. Se administró DMSO (excipiente de control) en igual volumen conjuntamente con el resto de los tratamientos. El momento de administración de las moléculas se consideró como el tiempo cero.
Prueba con Triatoma infestans
Se utilizaron 960 ninfas de quinto estadio de T. infestans criadas en laboratorio en condiciones ambientales constantes: 25 a 30 °C, 50 a 70 % de humedad relativa y fotoperiodo de 12 horas.
Para la alimentación de los insectos, estos se depositaron individualmente en tubos Eppendorf de 1,5 ml (figura 1). Se colocó tela adhesiva en la mitad superiorde la pared interior de cada tubo para permitir a los insectos trepar y llegar a la piel de los roedores. Para mantener lostubos Eppendorf en contacto con las ratas, se utilizaron planchuelas de poliestireno de 20 x 20 cm, en las que se incrustaron cinco tubos Eppendorf (un insecto por recipiente), de manera que la parte superior del tubo, cubierta por un tul, quedase al ras de la planchuela. Así, sobre cada planchuela se ubicó una rata en posición de decúbito ventral, previamente anestesiadacon ketamina (75 mg/kg de peso vivo) y acepromacina (2,5 mg/kg de peso vivo) por vía intraperitoneal, para garantizar su inmovilidad.
Los insectos fueron alimentados durante 45 minutos en los siguientes intervalos: 24 horas antes de la administración de las moléculas endectocidas y del DMSO a las ratas (-24 h: control basal), y 24, 72, 168, 240 y 336 horas después de la administración de cada compuesto. Por cada tratamiento y tiempo de observación, se evaluaron 20 insectos (cinco ninfas por rata por cada cuatro ratas y por tratamiento), según se detalla en el cuadro 1.
Mediante micropipetas de 100 y 10 µl, se midió el volumen de las excreciones emitidas por los insectos dentro de los tubos Eppenforf (figura 1), durante las primeras dos horas a partir del momento de alimentación.
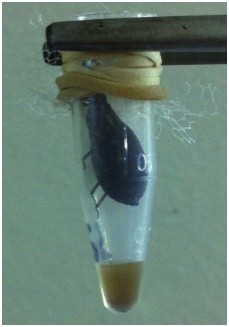
Figura 1 Ejemplar de quinto estadio de Triatoma infestans contenido en un tubo Eppendorf en cuyo fondo se pueden observar las excreciones emitidas por el insecto después de haber estado dos horas en contacto con una rata Wistar
La cantidad de sangre ingerida (g) se calculó aplicando la siguiente ecuación:
CSI = PETPA - PET,
donde CSI correspondía a la cantidad de sangre ingerida, PETPA, al peso de los tubos Eppendorf con el respectivo triatomino después de suministrado el alimento, y PET, al peso del respectivo tubo Eppendorf con el triatomino antes de ser alimentado sobre los roedores. Para este procedimiento, se utilizó una balanza de precisión Denver Instrument (USA).
Después de alimentarlos, se observó a los insectos cada 24 horas durante una semana. La mortalidad se definió como la incapacidad del triatomino para trasladarse sobre un papel de filtro, ya fuera de manera espontánea o incentivada por el contacto con una pinza metálica. Para calcular el porcentaje de mortalidad de las ninfas por efecto de los endectocidas, no se consideró el control basal, ya que los triatominos alimentados en ese momento no los habían recibido, como tampoco el DMSO. Con este control se estableció el nivel basal de la cantidad de sangre ingerida, el volumen de excreciones y la mortalidad de los insectos antes de la administración de los endectocidas y del excipiente a los roedores.
Análisis estadístico
Las variables de cantidad de sangre ingerida y volumen de las deyecciones de los insectos tras su alimentación sobre los roedores tratados con los diferentes endectocidas o con DMSO, se compararon mediante análisis de varianza de una vía (ANOVA). Las diferencias iguales o menores de 0,05 se consideraron significativas. Cuando se encontró significación en el ANOVA, se utilizó la prueba de Tukey de comparaciones múltiples de medias. En los casos necesarios, se usó laprueba de ji al cuadrado con corrección de Yates, para determinar la diferencia en los porcentajes de mortalidad de las ninfas de T. infestans.
Consideraciones éticas
El protocolo para el manejo de los animales en los distintos ensayos se basó en la Guide for the Care and Use of Laboratory Animals del Institute for Laboratory Animal Research (21) y fue aprobado por el Comité de Ética de la Facultad de Ciencias Médicas de la Universidad Nacional de La Plata, Argentina.
Resultados
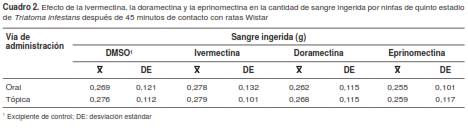
La cantidad de sangre ingerida por los insectos durante la alimentación se mantuvo constante a lo largo del ensayo y no se vio afectada por los tiempos posteriores a la administración del fármaco en que los triatominos entraron en contacto con las ratas, como tampoco sufrió variación alguna relacionada con el tratamiento previo recibido por el roedor (distribución F de Fisher-Snedecor, F=2,36; grados de libertad, gl=92; p=0,061) (cuadro 2). El promedio total de consumo de sangre fue de 0,268±0,115 g por insecto.
Cuadro 2 Efecto de la ivermectina, la doramectina y la eprinomectina en la cantidad de sangre ingerida por ninfas de quinto estadio de Triatoma infestans después de 45 minutos de contacto con ratas Wistar
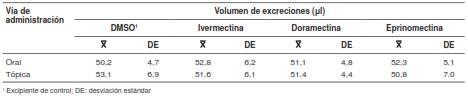
La variable del volumen de excreciones de los insectos tampoco sufrió cambios durante el experimento (F=0,56; gl=92; p=0,63) (cuadro 3). El volumen total de excreciones de los insectos durante las primeras dos horas a partir del momento de su contacto con los roedores fue de 51,7 ± 5,7 µl.
Cuadro 3 Efecto de la ivermectina, la doramectina y la eprinomectina en el volumen de excreciones de ninfas de quinto estadio de Triatoma infestans durante las dos primeras horas después de la alimentación
Por el contrario, los porcentajes de mortalidad de las ninfas variaron según la vía de administración, el tipo de endectocida administrado y elmomento de alimentación de las ninfas después de la administración de cada molécula a los roedores. Los porcentajesde mortalidad total entre los insectos del control basal (-24 h) y los insectos de control con el excipiente (DMSO) fueron idénticos (2,5 %).
En cuanto a la vía de administración, se encontró que la mortalidad de los triatominos fue significativamente más alta (23 %; 69/300 insectos) cuando los endectocidas se administraron de forma tópica a las ratas en los cinco momentos de alimentación después del tratamiento, que cuando se administraron por vía oral (9,3 %; 28/300 insectos; χ2=24,69, gl=1, p<0,001).
El endectocida que causó mayor mortalidad al aplicarlo por vía tópica fue la doramectina (28 %), seguido por laivermectina (21 %), la eprinomectina (20 %) y el DMSO (2 %), diferencias que fueron estadísticamente significativas (χ2= 25,26; gl=3; p<0,0001).
Con respecto a los momentos de alimentación después de la administración de las sustancias, la actividad de los tres endectocidas sobre las ninfas comenzó a las 24 horas de la alimentación. En este momento la mayor mortalidad se presentó con la ivermectina (40 %), comparada con la doramectina (35 %), la eprinomectina (25 %) y el DMSO (5 %), aunque las diferencias no fueron estadísticamente significativas (χ2=7,43; gl=3; p=0,059). A las72 horas, los tres compuestos endectocidas presentaron su máxima actividad: 80 % con la doramectina, 70 % con laeprinomectina y 65 % con la ivermectina, en tanto que el excipiente de control no ocasionó mortalidad alguna (0 %), y las diferencias fueron estadísticamente significativas (χ2=31,93; gl=3; p<0,001). Solamente la doramectina mantuvo su actividadhasta las 168 horas de la administración (25 %). A las 240 y a las 336 horas de la alimentación, no se presentó mortalidad con ninguno de los compuestos ensayados.
En cuanto a la administración oral, solo demostraron actividad insecticida la ivermectina (11 % ) y la doramectina (15 %), con diferencias estadísticamente significativas al compararlas con el excipiente de control (3 %) (χ2=8,55; gl=2; p=0,014).
Con respecto a los momentos de alimentación después de la administración, a las 24 horas la mortalidad observadafue de 20 % con doramectina y de 15 % con ivermectina, mientras que con el excipiente de control no se observó mortalidad alguna (χ2=4,20; gl=2; p=0,14).
La doramectina y la ivermectina demostraron su mayor eficacia en las ninfas a las 72 horas después de la administración, con 55 y 35 %, respectivamente, siendo estas diferencias estadísticamente significativascon respecto al excipiente de control (10 %;χ2=9,19, gl=2; p=0,011). A partir de las 168 horas de alimentación, no provocaronmortalidad.
El hecho de poder alojar a las ninfas en recipientes individuales permitió conocer la cantidad de sangre consumida por los triatominos que luego murieron, con promedios de 0,294 ± 0,063 g, 0,341 ± 0,048 g y 0,332 ± 0,056 g para la doramectina, la ivermectina y la eprinomectina, respectivamente; estas diferencias fueron estadísticamente significativas (F=6,26; g =92; p=0,002).
El análisis realizado con la prueba de Tukey demostró que los triatominos del tratamiento con doramectina tomaron una cantidad de sangre significativamente menor, comparados con aquellos de los tratamientos con ivermectina y eprinomectina. La diferencia mínima significativa entre medias (DMS) fue de 0,035 g, valor menor que la diferencia observada entre la doramectina y la ivermectina (0,047 g; p<0,05) y que la diferencia entre la doramectina y la eprinomectina (0,038 g; p<0,05). Por otra parte, la diferencia observada entre la ivermectina y la eprinomectina (0,009 g) no fue estadísticamente significativa (p>0,05).
Discusión
El presente estudio se llevó a cabo en el marco de la estrategia de complementación del uso masivo de piretroides contra T. infestans con moléculas que pueden controlar las poblaciones en el peridomicilio y el domicilio. Anteriormente, nuestro equipo había demostrado en ensayos in vitro la actividad de estos tres endectocidas en T. infestans, lo cual motivó la continuación del estudio aplicando distintos modelos animales. La primera prueba in vivo se llevó a cabo en gallinas ponedoras (22), y se pudo demostrar la actividad insecticida de la ivermectina en ninfas de quinto estadio de T. infestans (23,25 % de mortalidad), administrada únicamente en forma puntual (spot-on) y con una dosis igual a la empleada en el presente ensayo (600 µg/ml de una formulación de ivermectina al 0,5 % en DMSO). El efecto insecticida de la ivermectina en T. infestans se extendió hasta las 336 horas. Cada especie animal tiene características propias que inciden en el comportamiento farmacocinético de la molécula administrada por diferentes vías y, por ende, los procesos de absorción, distribución, metabolismo y excreción difieren. El patrón metabólico puede ser significativamente diferente entre aves y mamíferos, por lo cual la extrapolación entre especies no se recomienda.
En el presente estudio, el hecho de haber colocado a los triatominos en recipientes individuales y mantener anestesiadas a las ratas durante el tiempo de alimentación de los insectos permitió evaluar, además de la mortalidad de las ninfas, dos variables muy importantes que tienen relación directa con la efectividad de T. infestans para transmitir T. cruzi: la cantidad de sangre ingerida y el volumen de excreciones durante o tras la alimentación (23). Bajo las condiciones experimentales propuestas durante la alimentación, ninguno de los tres endectocidas tuvo incidencia en los dos procesos fisiológicos estudiados. Debe recordarse que, debido a su mecanismo de acción, las avermectinas interfieren en la alimentación de los ectoparásitos y los endoparásitos alterando la musculatura de la bomba faríngea (15). Según los resultados obtenidos, en T. infestans, la musculatura de la bomba faríngea aparentemente no se vio afectada, ya que no hubo interferencia en la alimentación de los insectos por acción de los endectocidas, comportamiento también observado en el estudio in vitro realizado por nuestro grupo de investigación (13). Esta hipótesis debería investigarse mediante estudios moleculares.
Sin embargo, sí se encontraron diferencias en los porcentajes de mortalidad de las ninfas según el endectocida administrado al roedor y la vía de administración empleada. Los tres endectocidas demostraron tener mayor eficacia sobre las ninfas cuando se administraron de forma puntual (spot-on), lo cual podría explicarse por la gran liposolubilidad de las moléculas, característica que les permite atravesar la barrera epitelial fácilmente, posibilitando una mayor biodisponibilidad, es decir, los insectos disponen de una mayor cantidad demoléculas en el momento de alimentarse. Por otra parte, el DMSO es un disolvente ideal, que se comporta como un transportador de moléculas y facilita la permeabilidad de las membranas. Si a estas características se añade la gran facilidad con que todas aquellas sustancias con propiedades simultáneamente hidrosolubles y liposolubles, como las avermectinas, pasan a través de la piel, se explicaría por qué se logró un mayor efecto en T. infestans tras la administración tópica.
También debe tenerse en cuenta que cuando se utiliza la vía tópica, se evitan las reacciones del primer paso metabólico en el hígado, fenómeno que sí ocurre cuando la vía de administración es la oral. La ivermectina, por ejemplo, sufre distintas reacciones químicas por acción del complejo enzimático citocromo P450, siendo las isoformas 1A1 y A3 las involucradas en el metabolismo del endectocida en ratas (24). Por lo tanto, con esta formulación experimental la vía tópica parece ser la más apropiada para lograr concentraciones suficientes en la sangre que ocasionen la muerte del triatomino.
Cabe destacar que, tanto en los estudios in vitrocomo en el presente ensayo, la doramectina fue el endectocida que produjo la mayor mortalidad en ninfas de quinto estadio de T. infestans. In vitro también se pudo determinar que la doramectina fue más potente que la ivermectina y la eprinomectina. La concentración mínima para obtener 100 % de mortalidad entre las ninfas fue de 1,6 ng/ml con la doramectina, mientras que con la eprinomectina se necesitó el doble de concentración (3,15 ng/ml) y, con la ivermectina, cuatro veces más (6,25 ng/ml) (13).
Esta mayor potencia de la doramectina podría estar relacionada con lo observado en el presente trabajo, en el cual los insectos que murieron por acción de la doramectina fueron los que, en promedio, ingirieron menor cantidad de sangre, hallándose una diferencia estadísticamente significativa con respecto a la ivermectina y la eprinomectina. En consecuencia, se podría aseverar que, al ser más potente, se necesitó una menor concentración de moléculas de doramectina para matar al insecto.
Otra explicación podría relacionarse con la mayor biodisponibilidad de la doramectina con respecto a la ivermectina y la eprinomectina, lo que determinaría una mayor cantidad de moléculas de doramectina en el plasma de las ratas disponibles para los triatominos durante la alimentación. La disposición plasmática de las avermectinas se caracteriza por su prolongada persistencia en el organismo y su gran volumen de distribución, así como por un efecto significativo de la formulación y la ruta de administración en la biodisponibilidad, y una gran variación entre especies e individuos.
Si bien los porcentajes de mortalidad total entre las ninfas no fueron elevados durante el ensayo, debe destacarse la gran actividad tóxica demostrada por los tres endectocidas en las ninfas alimentadas a las 72 horas (80, 70 y 65 % para la doramectina, la eprinomectina y la ivermectina, respectivamente) a partir de la administración tópica de las moléculas en los roedores, lo cual probablemente coincidió con la concentración plasmática máxima alcanzada tras la administración.
La pérdida de actividad de los tres endectocidas después de las 168 horas de la administración estaría relacionada con la caída de las concentraciones plasmáticas. Lógicamente, esta breve persistencia respondería a la formulaciónexperimental ensayada (solución en DMSO). En el estudioin vivo con gallinas (22), se encontró que el porcentaje de mortalidad observado en las ninfas se relacionaba directamente con la concentración plasmática de ivermectina en el momento de la alimentación de las ninfas sobre las aves, es decir, la actividad de estas moléculas se relacionó directamente con la concentración plasmática lograda y con su mayor persistencia.
Cuando se utilizó la vía oral para administrar los endectocidas en ratas, nuevamente se observó una mayor actividad de las moléculas a las 72 horas de la administración, 55 y 35 % para doramectinae ivermectina, respectivamente, en tanto que la eprinomectina fue el único de los endectocidas ensayados que no demostró actividad al ser administrado de manera oral, lo cual coincide con lo reportado por Zeng, et al., quienes, mediante estudios sobre el metabolismo de la eprinomectina en ratas Sprague-Dawley, determinaron que la mayorproporción de las moléculas de eprinomectina administradas por vía oral se eliminaban intactas en las heces, y solo un pequeño porcentaje de ellas sufría una N-desacetilación como reacción metabólica, de manera que su biodisponibilidad oral resultaba muy baja (25,26).
En el presente estudio, nos propusimos evaluar la actividad de tres endectocidas en ninfas de quinto estadio deT. infestans en un modelo in vivode roedor. De las tres variables estudiadas, la mortalidad fue la única que, bajo las condiciones experimentales, demostró una diferencia estadísticamente significativa entre triatominos alimentados a partir de ratas tratadas y aquellos que se alimentaron de ratas tratadas con el excipiente de control (DMSO). Los altos porcentajes de mortalidad en las ninfas obtenidos a las 72 horas de la administración representan un hallazgoalentador, sobre todo porque estos compuestos antiparasitarios podrían emplearse como complemento de los piretroides en el domicilio y en el peridomicilio.
Dada la breve persistencia obtenida con esta formulación experimental, uno de los objetivos de futuros trabajos sería ensayar distintas formulaciones de acción prolongada y constatar si, al prolongar la permanencia de las moléculas, se aumenta la eficacia frente a las ninfas.

