











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkIntroduction
Yellow passion fruit (Passiflora edulis f. flavicarpa) is one of the leading fruit species of the Neotropics due to its high nutritional content and economic potential as fresh or processed fruit (Ocampo Pérez et al., 2021). This species originated in Brazil and is currently cultivated in tropical areas on four continents from sea level to mountain areas up to 1,500 m a.s.l. (Faleiro et al., 2020). The flower is hermaphrodite with a high degree of self-incompatibility (93%) and is cross-pollinated (allogamy) mainly by insects of the genus Xylocopa spp. (González et al., 2009; Arias-Suárez et al., 2014).
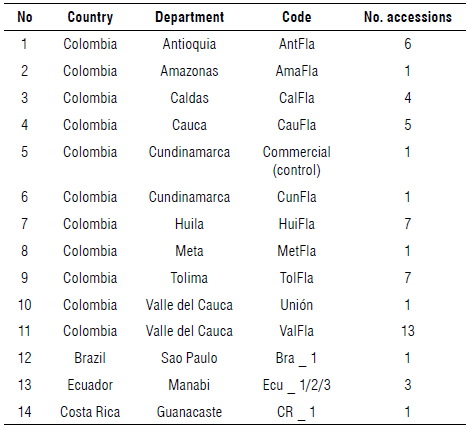
Yellow passion fruit was introduced to Colombia from Hawaii (USA) through Venezuela at the experimental station of the Colombian Agricultural Institute (ICA) in Valle del Cauca in the early 1960s (Morton, 1967). Colombian growers have contributed considerably to its domestication process by implementing new agronomic practices for better yield, such as pruning, fertilizer application, and the use of irrigation systems (Ocampo, Urrea, et al., 2013). There are currently about 8,165 ha cultivated with yellow passion fruit, with an average annual production of 137,622 t (Agronet, 2019). The departments of Antioquia, Huila, Meta, and Valle del Cauca concentrate the highest production with about 7,115 ha and an average yield that varies between 15 and 21 t ha-1 per year. Most of the production (60%) is destined for export, and national consumption is essentially supplied as fresh fruit, ice cream, desserts, and homemade juices. This production system suffers from multiple phytosanitary problems that prevent better crop development, such as collar rot [Fusarium solani f. sp. passiflorae (Mart.) Appel & Wollenweber], anthracnose [Col-letotrichumgloeosporioides (Penz.) Penz. & Sacc.], Soybean mosaic virus (SMV), Bean dwarf mosaic virus (BDMV), thrips (Neohydatothrips signifier Priesner), and lance fly (Dasiops inedulis Steyskal) (Hernández et al., 2011; Osorio Cardona et al., 2020), forcing the application of chemical treatments for crop protection. Additionally, the absence of certified nurseries and the lack of breeding programs have led to a decrease in the useful life of plantations from 36 to 18 months and a reduction in annual average yields to just 17.1 t ha-1 in the last decades (Agronet, 2019).
Genetic variability for agronomic traits is the key component of breeding programs for broadening the gene pool of crops to face the risk of epidemic plant diseases, obtain greater adaptability to different ecological zones, and ensure sustainable agriculture (Meletti et al., 2005; Freitas et al., 2011). In Colombia, some studies have been carried out on the genetic variability in some species of the genus Passiflora L. at the intra and interspecific levels using morphological descriptors (Primot et al., 2005; Ocampo Pérez et al., 2009; Viana et al., 2010; Checa Coral et al., 2011), molecular markers such as amplified fragment length polymorphism (AFLP) (Segura et al., 2002; Ocampo et al., 2004), and microsatellites (Ortiz et al., 2012; Bernal-Parra et al., 2014). These studies have established the distances between cultivated species and their wild relatives as a strategy for the exploration and conservation of genetic resources. They have also allowed the development of further research on interspecific hybridization between the main economically important species of Passiflora and their closest wild species (Ocampo et al., 2016).
Most of research efforts in yellow passion fruit have been focused on its characterization, selection, and breeding in Colombia (Restrepo & Kogson, 1998; Espitia et al., 2008; Galeano Mendoza et al., 2018). However, the sample size, composition, and origin of these studies limit their interpretation and do not allow a broader exploration of the intraspecific variability of this species. Ocampo, Urrea, et al. (2013) reported the collection and characterization of 44 yellow passion fruit accessions in different producing areas of Colombia, identifying eight elite materials with high fruit quality (weight >200 g, pulp percentage >50%, and °Brix >14.5) as promising for a breeding program. This same sample of accessions was characterized using microsatellite markers (Ocampo et al., 2017), finding relative genetic variability with certain geographic relationships between accessions of the same origin based on fruit-associated characters. While in Colombia variability is currently being explored, in Brazil, several yellow passion fruit cultivars have been released since the year 2000 for fresh consumption and the agroindustry (Meletti et al., 2000; Nascimento et al., 2003; Cerqueira-Silva et al., 2014; Viana et al., 2016) with significant impact in the last decade by improving fruit quality in this country (Cerqueira-Silva et al., 2016; Faleiro et al., 2020).
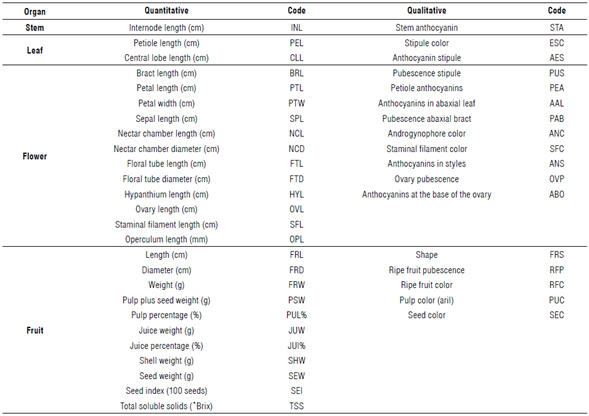
Characterization of agro-morphological descriptors has been used in several fruit cultivated species, such as Carica papaya L. (Ocampo et al., 2006), Theobroma cacao L. (Santos et al., 2012), and Annona muricata L. (Moreira-Macías et al., 2020), to establish the levels of intra and interspecific variability (Ocampo Pérez & Coppens d'Eeckenbrugge, 2017). These levels are easily accessible, and their information is essential for breeding that cannot be based primarily on DNA sequences. A descriptor is a characteristic or attribute whose expression is easy to measure, register, or evaluate and refers to the shape, structure, or behavior of an accession (Hidalgo, 2003). These descriptors are based on the measurement of quantitative and qualitative characters of different plant organs, such as stem, leaf, flower, and fruit. However, these characters must be measured at the same locality to avoid environmental influence (Hidalgo, 2003). In Brazil, Crochemore et al. (2003) established high differentiation in 56 accessions of Passiflora species, but with small divergences within yellow passion fruit accessions. Studies carried out by Ocampo Pérez et al. (2009) and Ocampo Pérez and Coppens d'Eeckenbrugge (2017) demonstrated that 23 quantitative and 18 qualitative descriptors could detect variability between and within 61 Passiflora species, including accessions of P. edulis f. fla-vicarpa.Castro et al. (2012) defined 16 quantitative and six qualitative descriptors for passion fruit with discriminating capacity by analyzing 24 genotypes grown in Brazil. These studies highlight that the agro-morphological descriptors that contribute most to the total variance are related to flower and fruit traits.
Collection and characterization studies carried out in Colombia by Ocampo, Urrea, et al. (2013), Ocampo et al. (2017), and Galeano Mendoza et al. (2018) are the basis for further research in the search for new planting materials for different ecological niches with greater hardiness and productivity. Therefore, the aim of this study was to establish the genetic relationships between individuals and accessions of different geographical origins through morphological characterization based on desirable agronomic traits, allowing the selection of elite yellow passion fruit accessions.
Materials and methods
Study area
This research was carried out at the experimental station of the Luker farm (5°04'25.95" N; 75°41'4.71" W) located in the municipality of Palestina, department of Caldas (Colombia) at 1023 m a.s.l. The edaphoclimatic conditions registered in this locality were as follows: soil with a loamy texture, pH from 5.5 to 6.0, 9% organic matter, an average environmental temperature of 23°C, heliophany of 190 h per year, average relative humidity of 75.6%, and total annual average precipitation of 2200 mm.
Plant material and field trials
The germplasm included 52 commercial accessions collected in previous studies by Ocampo, Urrea, et al. (2013) in different geographical areas of Colombia and other producing countries (Tab. 1). In total, the sample comprised 104 individuals, and each genotype was obtained through open pollination, considering half-siblings within each accession. Seeds of each accession were germinated in 12x17 cm plastic bags (1.9 kg) using substrate composed of soil and rice husk at a 3:1 ratio. Plants were established in the field 40 d after germination. Agronomical practices such as fertilization, weed control, pruning, and pest and disease control were adopted according to the recommendations of Jaramillo Vásquez et al. (2009) with emphasis on the use of biological products (Trichoderma harzianum and Pachycrepoideus vindemiae). Seedlings were sown 3 m between rows and 3 m between plants in a simple trellis system and under a completely randomized block experimental design with 52 treatments (accessions) and five replicates per experimental unit.
Morphological characterization
In each block, the most vigorous plant per accession was selected, and this plant was characterized with a list of 23 quantitative and 18 qualitative descriptors associated with vegetative, floral and fruit characters selected by Ocampo Pérez et al. (2009), Castro et al. (2012), and Ocampo Pérez and Coppens d'Eeckenbrugge (2017) (Tab. 2). For the quantitative descriptors, five measurements were made per descriptor in each plant organ using a digital caliper, and weighing was done directly using a digital scale. The traits related to fruit quality were analyzed at the Quality Laboratory at the experimental station of the Luker farm, and total soluble solids (°Brix) per fruit were measured with a manual refractometer on a scale of 0 to 32. All qualitative characteristics were recorded as binary numbers, i.e., presence (1) or absence (0), and the colors of each organ were compared with the Royal Horticultural Society color chart (R.H.S., 2001). The shape of the fruit was recorded and divided into three categories as oval (1), spherical (2) and oblate (3) based on the index generated between diameter and length (fruit diameter (FRD)/fruit length (FRL).
Agronomic evaluation
The incidence of pests and diseases (percentage of plants affected) was registered in 10 plants per accession (five replicates) every 15 d for 12 months after sowing the plants of each accession in the field, for a total of 24 phytosanitary evaluations. The number of days after the first flower (DAF; anthesis) was recorded when at least 50% of the plants of each accession bloomed. Fruit yield (kg ha-1, YLD) was determined by the sum of the total of harvests carried out during 8 months after the first harvest. Yield was calculated in kilograms, collecting the fruits of the five plants in each accession per block. The identification of pest insects and diseases was carried out in the Plant health laboratories of Universidad de Caldas and the International Center for Tropical Agriculture (CIAT) - Bioversity International, following the methodologies proposed by Morales et al. (2006), Fischer and Rezende (2008), and Hernández et al. (2011).
Selection of superior genotypes
A selection index (SI) based on a linear combination of economic weight for the traits yield, precocity (DAF), and fruit quality for market requirements was calculated according to Baker (1986). The values of each variable were standardized (E) using the following equation:

where X ij is the individual observation, m is the general average of each accession per variable, and sd is the standard deviation.
Once the data were standardized, the SI was established according to the following equation:
Ocampo, Marín, and Urrea: Agro-morphological characterization of yellow passion fruit (Passiflora edulis f. flavicarpa Degener) reveals elite genotypes for a breeding program in Colombia
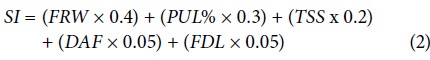
where FRW is the fruit weight estimated in t ha-1, PUL% is the percentage of pulp plus the seeds, TSS is the content of total soluble solids (°Brix), DAF is the days after the first flower or precocity, and FDL is the ratio between fruit diameter and length.
Statistical analysis
The main traits of fruit quality (weight, pulp content, and total soluble solids), and yield were subjected to an analysis of variance (ANOVA; P≤0.05) and the Tukey's mean difference test. The quantitative descriptors were subjected to univariate and multivariate principal component analyses (PCA) using varimax normalized function. The qualitative descriptors underwent a multiple correspondence factor analysis (MCFA). The analysis of classification by the neighbor-joining method (Saitou & Nei, 1987) was used with the quantitative descriptors utilizing Euclidean distances, and the qualitative descriptors employed the distances of Sokal and Michener (1958). Data were analyzed with the software STATISTICA® v.8.1 and DarWin® v.5.01.
Results
Agronomic evaluation
Precocity
The 52 passion fruit accessions showed an average of 119 DAF with a coefficient of variation of 14.2% (Tab. 3). Likewise, 51% of the accessions reached their first bloom below the average value, and only seven of them between 86 and 105 d (CauFla04, ValFla13, HuiFla04, CalFla04, Ecu_2, Commercial, and ValFla07). In contrast, TolFla01, HuiFla02/3, Ecu_1, and CauFla02/03, with more than 130 DAF, come from regions distant from the area where this study was carried out.
TABLE 3 Average of the trait's precocity, yield, and fruit quality for market requirements in the yellow passion fruit germplasm.
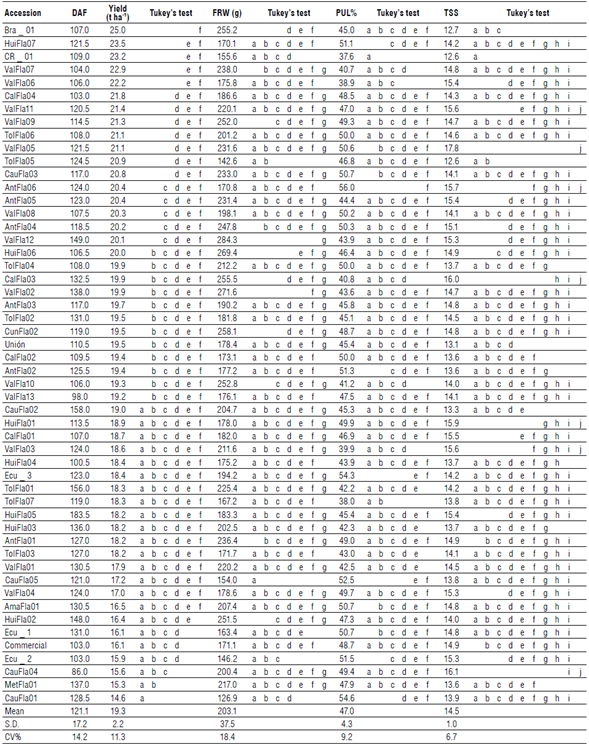
DAF - days after the first flower; FRW - fruit fresh weight (g); PUL% - percentage of pulp; TSS - total soluble solids (°Brix). Lowercase letters indicate the differences between the treatments. S.D. - standard deviation; CV - coefficient of variation.
Yield
The ANOVA showed significant differences between the means of each treatment (52 accessions), with a P<0.05. The yield during the first eight months after the first fruit harvest registered an average of 19.3 t ha-1, ranging between 14.6 and 25 t ha-1 (Tab. 3). Thirty percent of the accessions reached yields of more than 20 t ha-1 and were associated with the departments of Valle del Cauca (Val-Fla05/06/07/08/09/11/12), Tolima (TolFla05/06), Antioquia (AntFla04/05/06), Huila (HuiFla07), Cauca (CauFla03), and Caldas (CalFla04), and the countries Brazil (Bra_1), and Costa Rica (CR_1). In contrast, accessions from Cauca (CauFla04/5), Meta (MetFla01), Ecuador (Ecu_2), and the commercial control showed yields below 16.2 t ha-1. A point to highlight is the discreet yield of the commercial control accession (16.1 t ha-1), with a value lower than 90% of the evaluated accessions. The divergence found in yield between all accessions can be explained by the degree of adaptability of the accessions to the environmental conditions of the study area.
Incidence of pests and diseases
The phytosanitary evaluation carried out for 12 months recorded the incidence of five pest insects during the 24 evaluations carried out (Fig. 1A). Thrips (Neohydatothrips signifier) were the most limiting insects in the collection, with a 98% incidence in all accessions, affecting terminal shoots and causing growth retardation. Lance fly (Dasiops inedulis) ranked second in importance with 77%, followed by the defoliation worm [Dione juno (Cramer)] with 22%, affecting mostly the leaves. Mites (Tetranychus ludeni Zacher) and aphids (Aphis gossypii) were also detected attacking young and adult leaves with 10% and 21% incidence, respectively.
Accessions with a low pest attack (21% of incidence) were identified. Thrips were the main pest of economic importance affecting yellow passion fruit, particularly the accessions from Valle del Cauca (ValFla01/03/06/08), Huila (HuiFla01), Antioquia (AntFla03), and Tolima (TolFla01). The accession from Ecuador (Ecu_02) showed an incidence of less than 2%, indicating a high potential for the selection of genotypes with pest tolerance.
Four diseases were registered affecting the development of the 52 accessions (Fig. 1B). Scab [Cladosporium clado-sporioides (Fresen.) G.A.de Vries] was the disease with the highest incidence (100%), affecting branches and fruits at all stages of the crop cycle. Viral symptoms (Soybean mosaic virus - SMV) in the leaves were also detected in 96% of the accessions after the first harvest until the end of the evaluation. Bacteriosis [Xanthomonas axonopodis pv. passiflorae (Pereira) Gonçalves & Rosato] and anthracnose [Colletotrichum gloeosporioides (Penz.) Penz. and Sacc.] showed severe symptoms in the branches, leaves, and fruits with an incidence of 48% and 83%, respectively. The latter was characterized by partial defoliation and loss of plant vigor, and such symptoms were more affected by increased rainfall and high relative humidity.
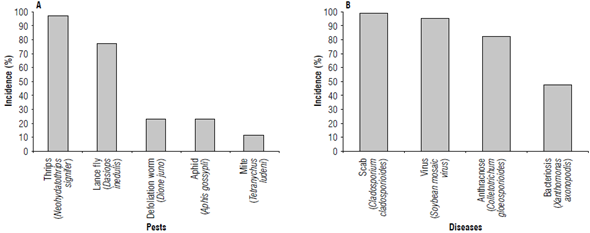
FIGURE 1 Phytosanitary evaluation carried out during 12 months in 52 yellow passion fruit accessions.
The incidence of diseases between the accessions was more variable than the incidence of pests. Seven accessions (TolFla03/06, HuiFlaOÓ, CauFla04, CunFla02, Bra_01, and CR_01) showed tolerance to diseases (C. cladosporioides) that mainly attack fruit shells lowering their marketing quality as fresh fruit. A positive reaction to the attack of bacteriosis and anthracnose was observed in 57% of the accessions with less than 2% incidence; VaLFla04 and the commercial control showed no visible symptoms. Regarding viral attacks, mainly SMV, accessions AmaFlaOl, Cau-Fla04, and HuiFla02 did not show symptoms of the disease, and the remaining accessions did not exceed 13% incidence. The results of this phytosanitary evaluation suggest a second agronomic evaluation to confirm these results.
Selection of elite accessions
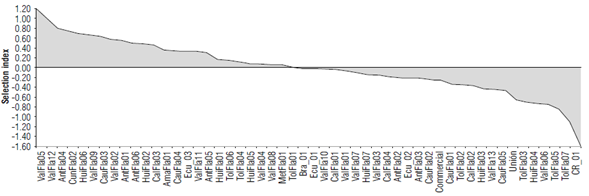
The SI analysis identified 48% of the accessions with desirable agronomic traits (Fig. 2). These accessions combine the best fruit characteristics, earliness or precocity and yield. Thus, when selection pressure of 10% was applied, six elite accessions were identified: Valle del Cauca (ValFla05/09/12), Antioquia (AntFla04), Cundinamarca (CunFla02), and Huila (HuiFlaOÓ) with a SI higher than 0.66 (Tab. 4). Additionally, accession ValFla05 showed the highest values of pulp percentage (50.6%) and total soluble solids (17.8 °Brix), while ValFla09 and HuiFla06 showed high yield (21.3 t ha-1) and a low number of days after the first flower (106.5). The average fruit fresh weight varied between 231.6 to 284.3 g, and the accessions ValFlal2 and HuiFla06 were characterized by fruits of more than 269 g. In contrast, 27% of the accessions recorded a SI lower than -0.25, showing lower values of fruit quality, precocity, and yield.
The commercial control showed yield of less than 16.1 tha-1 compared to the general average of 19.21 ha-1 among the 52 accessions evaluated. The Brazilian commercial accession Bra_l showed high qualities regarding average yield (25 t ha-1), precocity (107 DAF), and fruit fresh weight (255.2 g), but pulp percentage (PUL%) of only 45% and low content of total soluble solids (12.7 °Brix), which is supported by its low SI (-0.02). The commercial accessions from Ecuador (Ecu_1/2/3) obtained a selection index rate between -0.02 and 0.33, showing yield of less than 18.5 t ha-1 and fruit weight <163.5 g, but average PUL% and TSS higher than 50% and 14.5 °Brix.
Quantitative morphological variability
Univariate analysis
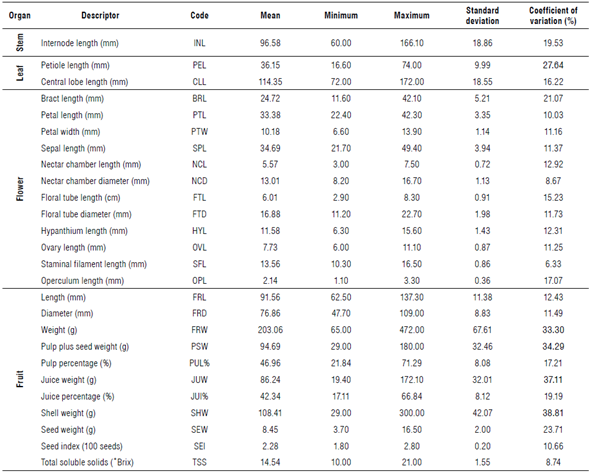
The estimated genetic variability was calculated for the 26 descriptors analyzed using descriptive statistics (Tab. 5). The average coefficient of variation (CV) was 17.67% and only the descriptors related to petiole length (PEL), and the weights of the fruit (FRW), pulp (PSW), juice (JUW), and shell (SHW) exceeded 25% of the variation. Descriptors associated with the diameter of the nectar chamber (NCD), the length of the stamen filaments (SFL), and the seed index (SEI) showed a CV below 10%, suggesting that these do not contribute to the total variation. The main descriptors related to fruit quality, such as TSS and PUL% showed relative variation (10.66 and 17.21%) that can be considered for the selection of elite individuals. This pomological variation is the consequence of an unsystematic selection of plants with uniform fruit characteristics by the producers in each harvest cycle.
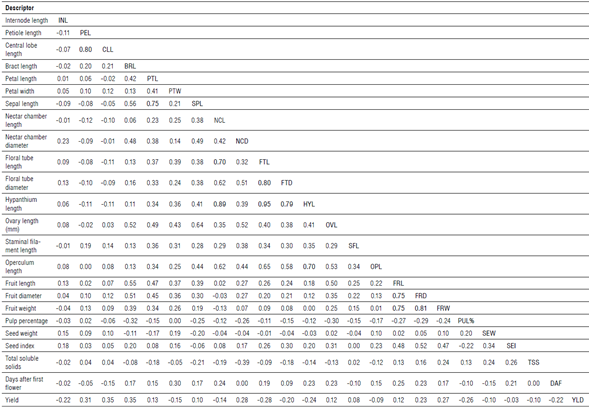
Phenotypic correlations
The average correlation analysis between the morphological and agronomic descriptors was r = 0.14 (Tab. 6). The most significant correlations showed values higher than r≥0.70 and were represented by the most morphologically related descriptors. The most related were floral tube length (FTL) and hypanthium length (HYL) vs. floral tube diameter (FTD), petiole length (PEL) vs. central lobe length (CLL), and sepal length (SPL) vs. petal length (PTL) with an r>0.74. Regarding the descriptors of fruit quality, only FRW and dimensions (FRL and FRD) showed an average correlation coefficient of r = 0.77. Total soluble solids did not show significant correlations, although it is negatively related to the NCD with r = -0.39. The yield (t ha1) exhibited moderate correlations with PEL, CLL, and bract length (BRL), with values between 0.31 and 0.35.
Multivariate analysis
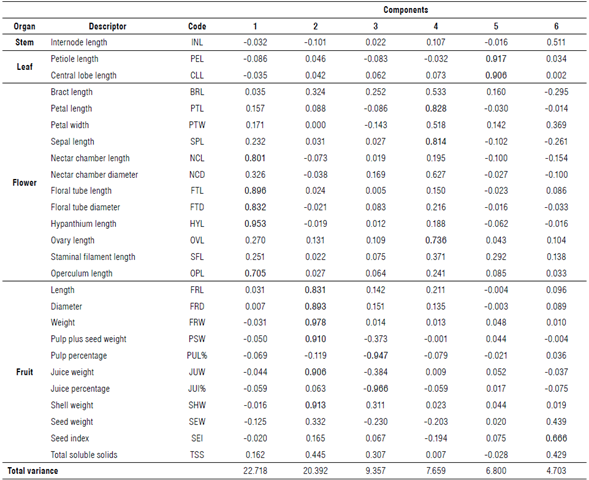
The quantitative descriptors were subjected to a principal component analysis (PCA), and only six were retained with an eigenvalue higher than 1, explaining 71.6% of the total variance (Tab. 7). The first component explains 22.7% of the variance related to flower length descriptors (nectar chamber length (NCL), FTL, FTD, HYL, and operculum length (OPL)). The second component assumed 20.4% of the variance and includes fruit descriptors, such as dimension (FRL and FRD) and weight (FRW, PSW, JUW, and SHW). The third (9.4%) was associated with PUL% and JUW (%). The last three components (4, 5, and 6) explained 19.2% of the variance and were associated with the PTL, SPL, ovary length (OVL), PEL, CLL, as well as seed weight (SEW). The PCA identified 42% of the descriptors proposed in this study and mainly those that were retained in the first two components, with 43.1% of the variance explained.
TABLE 7 Principal component analysis (PCA) performed on the quantitative descriptors used in the yellow passion fruit collection evaluated.
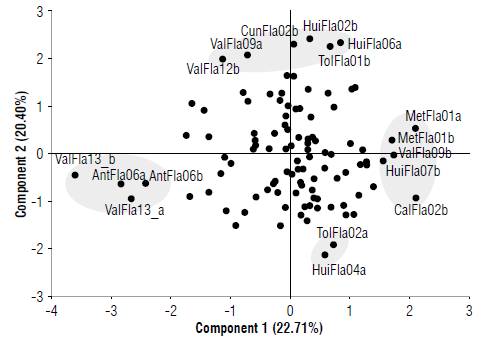
Plants of each accession were projected in the main plane (Fig. 3). In the first component on the left side, accessions ValFla13ab and AntFla06ab appear characterized by having small flowers, while in the other extreme, MetFla01a,b, CalFla02b, and ValFla09b showed the largest flowers. In the upper part of the second component, the accessions with the largest fruit size and weight such as CunFla02b, Tol-Fla01b, HuiFla02b, HuiFla02a, ValFla09b, are distributed. In the lower part, two plants of the accessions TolFla02a and HuiFla04a are differentiated with small and light fruits. In general, the main plane is dominated by a central cloud, where only 30% of the total accessions move away from the point of origin (0). These results suggest that the descriptors retained in the first two components do not allow grouping or clear differentiating of the accessions studied according to their geographical origin. However, descriptors related to flower and fruit size can contribute to the selection of elite individuals.
Tree classification
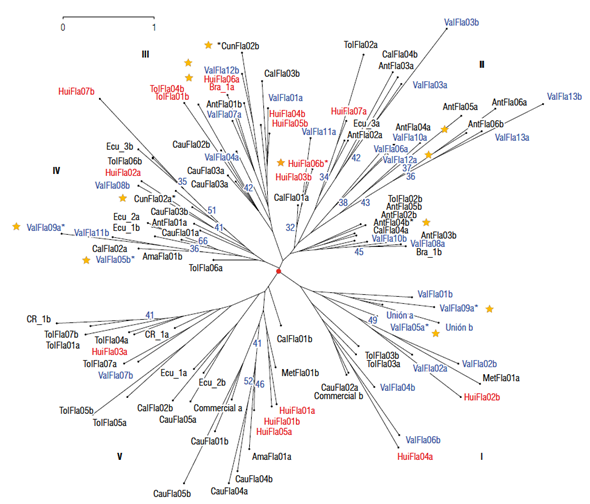
Classification analysis of the accessions utilizing the neighbor-joining method was carried out based on the results of the three principal components (PCA) that explained 52.45% of the total variance (Fig. 4). The dendrogram shows five main groups with little differentiation (bootstrap<30) and with high heterogeneity among the individuals of the same accession. The distances among the plants of each accession are higher than among the main groups. This is clearly observed in the accessions from Valle del Cauca (ValFla), Huila (HuiFla), and Tolima (TolFla) that are distributed in all groups. The accessions from Ecuador (Ecu_1/2/3) and Brazil (Bra_1) do not show a considerable differentiation in relation to the Colombian accessions; additionally, plants within the same accession are placed in different groups. In general, grouping of individuals is unsystematic, with some exceptions such as the accessions Union and TolFla03 (group I), ValFlal3, AntFla06, and HuiFl06 (group II), and TolFla05 and CauFla04 (group V). The six elite accessions (ValFla05, ValFla09, ValFlal2, AntFla04, CunFla02, and HuiFla06) are distributed in four groups and their individuals are well differentiated.
Multiple correspondence factor analysis
The characterization showed that descriptors related to the color of the androgynophore (ANC) and staminal filaments (SFC), the pubescence of the ovary (OVP), and the yellow color of ripe fruit (RCF) did not show variability in the evaluated accessions. These descriptors were excluded, and a MCFA was performed with 13 qualitative polymorphic descriptors. The MCFA was represented by four factors that explain 44.4% of the total variability (inertia). The first factor retained 13.4% of the variability and was related to the absence of pubescence in the upper part of the bract (PAB01), anthocyanins in the petiole (PEA01), anthocyanins in the abaxial surface of the leaf (AAL01), and the pale-yellow color in the fruit (RFC03). In the second factor (11.5%), the descriptors related to the presence of anthocyanins at the base of the ovary (ABO02) and in the androgynophore (ANA02), absence of anthocyanins in the stem (STA01), pubescence in the mature fruit (RFP01) and dark green color in the stipule (EST02) stood out. The presence of pubescence (PUS02), absence of anthocyanins (AES01) in the stipule, and fruit shape (FRS) represented 19.5% of the total variability (factors 3 and 4). The FRS showed three categories, ovate, spherical and oblate, across the total accessions with an index of shape (ISH = FRD/ FRL) that varied between 0.65 and 1.17. Thus, 96% of the harvested fruits were ovate (ISH<1), 3% spherical (ValFla05, TolFla05, AntFla02, Bra_l, and Union), and some plants of the accessions HuiFla03, CauFla03 and AntFla03 were oblate (ISH>1).
The accessions were projected in the principal plane generated by the MCFA and showed a similar distribution compared to the PCA with the quantitative descriptors. In this main plane, the accessions were not clearly grouped, and in most cases, they did not follow a pattern according to the geographical origin (Fig. 5). In the first factor, the Cauca accessions (CauFla03b/04a) stood out with the absence of pubescence in the bract (PAB01). On the opposite side of factor 1, some accessions from Valle del Cauca (ValFla07a/08a), Antioquia (AntFla01b/06b), Huila (HuiFla04b), Tolima (TolFla04b/07a), and Caldas (CalFla04a) were characterized by the absence of antho-cyanin on the petiole (PEA01) and on the abaxial surface of the leaf (AAL01), and by a pale-yellow color of the fruit (RFC03). For the second factor, accessions MetFla01a,b (Meta), AntFla02a,b (Antioquia), ValFla02a/09a,b (Valle del Cauca) and Ecu_2a/b (Ecuador) showed anthocyanin at the base of the ovary (ABO02) and on the androgynophore (ANA02), and its absence in the stem (STA01).
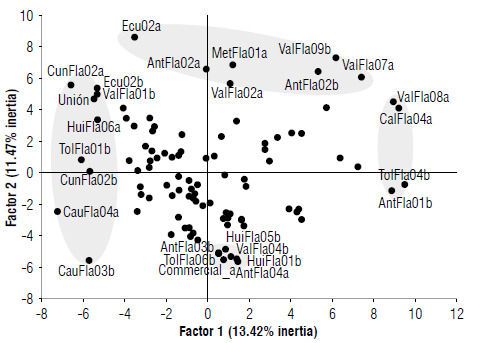
FIGURE 5 Yellow passion fruit accessions projected in the main plane generated by the multiple correspondence factor analysis (MCFA) using qualitative descriptors.
Tree classification
The maximum variability determined by the MCFA with the four factors (44.4% of inertia) was retained to construct the classification analysis of the qualitative descriptors (Fig. 6). Thus, the dendrogram showed five poorly supported main groups (bootstrap<30) with a similar structure to that obtained with the quantitative descriptors and shorter distances between the individuals of each accession. The accessions from Valle del Cauca (ValFla), Huila (HuiFla), and Tolima (TolFla) are distributed in the five branches, with no association among the plants that comprise them. Despite the high heritability of the qualitative traits, there is a high divergence between individuals of the same accession, with some exceptions such as ValFlaOl, ValFla03, TolFla07, ValFlal3, and Union. These two accessions are selections from Valle del Cauca and show some uniformity compared to both types of descriptors.
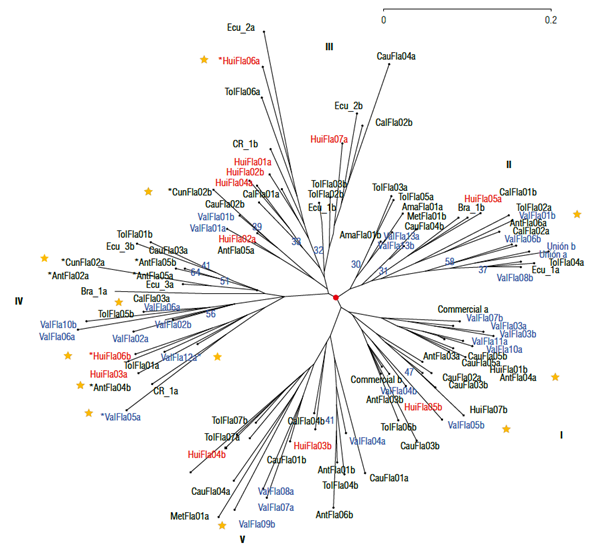
FIGURE 6 Classification analysis of yellow passion fruit accessions using the neighbor-joining (NJ) method employing qualitative descriptors. Yellow stars indicate elite accessions.
The plants of the six elite accessions are scattered in the five groups and especially in group IV, where the accessions from Brazil (Bra_01), Costa Rica (CR_01), and Ecuador (Ecu_03) are also grouped. Despite the high heritability of qualitative descriptors, the plants of each accession appear separated into different groups (ValFla05 - groups I and IV; ValFlal2 - groups II and IV; AntFla04 - groups I and IV; CunFla02 - groups III and IV; ValFla09 - groups IV and V, and HuiFlaOÓ - groups III and IV). Overall, the qualitative descriptors show similar structure variability regarding the quantitative descriptors in the yellow passion fruit collection assessed, reflected by the differences between the accessions and their plants. The low geographical structure of the individuals makes it challenging to identify duplicates within the collection for conservation if the origin criterion of the accessions is used.
Discussion
Pomological variability
In this study, 11 characters related to the fruit register high heterogeneity as reported by Ocampo, Urrea, et al. (2013), with a CV higher than 33% in FRW, PSW, JUW, and SH W. However, in this study, TSS showed values between 10 to 21 °Brix with little variation (CV = 11%). The differences between studies can be explained by the origin of the germplasm collected by Ocampo, Urrea et al. (2013) which was characterized in situ in different producing areas. This germplasm was influenced by edaphoclimatic offer and determined by the genotype x environment interaction.
The high degree of genetic exchange in the species also contributes to the heterogeneity in the quality of the harvested fruits and the progressive genetic degeneration of the cultivated genotypes. However, the variability found in the 52 characterized accessions allows selecting plants with large to small fruits depending on the market's demand for fresh or processed consumption (Tab. 4). The results showed that several accessions with yield higher than 20.0 t ha-1 have a PUL% lower than 45%. This suggests that the shell represents more than half the weight of the fruit and is an agronomic parameter not desirable for the yellow passion fruit market (Viera et al., 2020) that should be considered in the selection process.
Precocity and yield
Yield is a function of the production obtained in each harvest and depends largely on the days after the first flower (DAF) and the environmental conditions where the orchards are established (Wallace & Enriquez, 1980; Viera et al., 2020). The precocity of the harvest is beneficial as it brings advantages to geneticists for breeding processes and is excellent in economic terms for producers. In this way, 46% of the accessions have an average of less than 121 DAF; accessions CauFla04 and ValFla13 were particularly more precocious with 86 and 98 d, respectively. This agronomic trait that estimates the precocity in flower production showed that most of the accessions can adapt to the ecological conditions of the area where they were evaluated. However, the yield of most of these precocious accessions was lower than the average (19.3 t ha-1). The correlation between precocity and yield was r = -0.22 (Tab. 6), indicating that these traits are not always related, and in some cases, the effect is inverse. The commercial control accession showed a high precocity with 103 DAF; however, its yield reached only 16.1 t ha-1. Although precocity is a highly heritable trait (Acquaah, 2012), the results shown in this study suggest that it would not be effective in the selection processes in yellow passion fruit. Precocity must complement yield and fruit quality; thus, breeders should pay more attention to these precocious genotypes.
The national annual average yield in Colombia is 17.1 t ha-1 (Agronet, 2019), with maximum values between 19.9 and 20.6 t ha-1 in the departments of Meta and Antioquia, respectively. In this research, 83% of the evaluated accessions exceed this value and ten exceed 21 t ha-1 (Val-Fla05/06/07/09/11, TolFla06, HuiFla07, CalFla04, Bra_01, and CR_01). The low yield of the commercial control (16.1 t ha-1) questions the crop quality of this material that is currently offered to Colombian growers without any agronomic and scientific validation. The period between the fertilization of the flower and fruit ripening for harvest in yellow passion fruit varies between 50 to 60 d (Arias-Suárez et al., 2014), and if this is considered, the precocious accessions (DAF<121) with good yield (>21 t ha-1) constitute an important promising genetic material since their first harvest could start before six months. The good yield reached by most of the accessions in Valle del Cauca is explained by the selection processes of the best genotypes carried out by growers since the 1950s when passion fruit was introduced to Colombia (Morton, 1967; Ocampo, Urrea, et al., 2013). This department is a reference for the cultivation of yellow passion fruit and many growers from other regions have established new orchards with seed from this area. However, in recent years, phytosanitary problems, climate variability, and the absence of cultivars have affected the crop, preventing its expansion to new areas (Ocampo, Arias, et al., 2013; Galeano Mendoza et al, 2018). The differences in precocity and yield of the accessions were due to the interaction of the genotype x environment that affects the agronomic behavior of the different passion fruit materials that come from different regions in Colombia and other countries (Viera et al., 2020).
Reaction to phytosanitary problems
Plant pests and diseases are a complex problem that generates significant losses in yield and income for farmers. These effects are immediate and impose chemical treatments for the protection of orchards, reducing productivity and the cycle duration and leading to the loss of competitiveness of agricultural systems (Fischer & Rezende, 2008; Faleiro et al., 2019; 2020). Phytosanitary evaluations show that thrips (Neohydatothrips signifer), lance fly (Dasiops inedulis), scab (Cladosporium cladosporioides), viruses (SMV), and anthracnose (Colletotrichum gloeosporioides) are the insects and pathogens with the highest incidence (>75%). These phytosanitary problems have been reported throughout the producing areas of the country (Morales et al., 2006; Wyckhuys et al., 2010; Monje et al., 2012), with a higher prevalence in orchards located between 500 and 1,000 m a.s.l. The accessions collected by Ocampo, Urrea et al. (2013) and evaluated in this study were also severely affected by these phytosanitary problems. Pest insects are favored (>30%) during summer, while diseases increase (>40) their frequency during rainy periods. Thrips, viruses and collar rot (Fusarium solani) are the major limitations for passion fruit production in Colombia, since they cause considerable economic losses (Ocampo, Arias, et al., 2013; Osorio Cardona et al., 2020). In the evaluated germplasm, several accessions such as ValFla05/12/09, AntFla04 and HuiFla06 responded positively to the pressure of these agents during the 24 phytosanitary evaluations carried out, showing high fruit quality and yield higher than 20 t ha-1. This study shows that accession ValFla09, identified by Ocampo, Urrea et al. (2013) as promising, does indeed show good agronomic traits that can respond to the needs of producers.
Selection of elite accessions
The genetic potential in crops determines growth, yield, and quality traits (Acquaah, 2012). However, identifying genotypes with yield potential and good fruit quality traits is a job of extreme precision for geneticists (Moraes et al., 2005; Chavarría-Perez et al., 2020). Selection indices are a strategy widely used by breeders in various crops for plant selection and the establishment of working collections with the best genotypes (Bos & Caligari, 2008; Lagos et al, 2015). The six elite accessions identified in the study with a selection index higher than 0.66 (ValFla05, ValFla09, ValFla12, AntFla04, CunFla02, and HuiFla06) constitute significant progress for future work on the breeding of yellow passion fruit in Colombia. Nevertheless, this germplasm must be evaluated in other regions of the country under a recurrent selection scheme to determine its genetic potential in other agroecological zones. Thus, this strategy will enable the increase in the frequency of favorable alleles that control the quantitative traits, such as total soluble solids (°Brix), pulp weight, and yield. This methodology, combined with SI, has been used successfully in Brazil as an alternative to accumulate intrapopulation genetic gain (Silva et al., 2016) and selection of superior progeny through recurrent selection in passion fruit (Silva et al., 2009; Rodrigues et al., 2020). Recurrent selection has also been used in Brazil to develop the yellow passion fruit cultivar 'UENF Rio Dourado' (Viana et al., 2016) with yields of up to 25 t ha1. In Colombia, using an SI, 30 promising passion fruit accessions were identified by Galeano Mendoza et al. (2018) as possible parents for a first cycle of recurrent selection. Another important genetic resource is the elite passion fruit accessions identified by Ocampo, Urrea et al. (2013) in Colombia, whose desirable agronomic traits are evidenced in this research. The results confirm the genetic potential of each accession and suggest that productivity does not only depend on agronomic management and environmental factors, as reported in other studies on passion fruit (Araújo et al., 2006). This germplasm should also be used to obtain cultivars that respond better to water stress and tolerate saline-sodium soils, which might be key in the near future.
On the other hand, the commercial accessions from Ecuador (Ecu1/2/3) obtained a selection index rate between -0.02 and 0.33, showing yields below 18.5 t ha-1 (16.1 to 18.4) and fruit weight <163.5 g, although the average PUL% and TSS were higher than 50% and 14.5 °Brix. These accessions show agronomic characteristics similar to the Colombian commercial control and superior to those reported by Viera et al. (2020) in the Ecuadorian Pacific coast, where the orchards do not exceed average yield of 6 t ha-1 and 14 °Brix.
Morphological relationships
The use of descriptors in the morphological characterization of germplasm is essential for structuring breeding programs (Bioversity International, 2007). Phenotypic descriptors allow direct access to variability and must be evaluated in the same location due to environmental influence (Bioversity International, 2007). The multivariate analysis grouped the 23 selected quantitative descriptors into six principal components explaining 71.6% of the variance, of which 11 are those that contribute the most to the variability. These descriptors were related to flower length (NCL, FTL, FTD, HYL, and OPL), dimension (FRL and FRD), and fruit fresh weight (FRW, PSW, JUW, and SHW). Ocampo Pérez et al. (2009) report similar values, with 77.9% of the variance explained by the same set of descriptors in P. edulis f. flavicarpa.Castro et al. (2012) mention that the descriptors related to the fruit are essential to estimate the genetic variability in yellow passion fruit. At the interspecific level, the descriptors associated with the flower are those that contribute the most to the variability and allow ranking the species of the genus Passiflora (Villacis et al., 1998; Ocampo Pérez & Coppens d'Eeckenbrugge, 2017). However, Crochemore et al. (2003) mention that vegetative descriptors, such as leaf dimensions, contribute to interspecific variability in Passiflora. The quantitative descriptors identified in this study are confirmed as a powerful tool to explore variability in yellow passion fruit accessions. The neighbor-joining classification analysis showed five main groups with low geographical structure. This may be a consequence of the inter-cross pollination present in P. edulis f. flavicarpa since the plants of each accession originate from seeds of half-siblings and constitute different genotypes.
The five groups identified are higher than the groups reported by Ocampo et al. (2017) with microsatellite markers, with the same germplasm size and constitution. In the molecular study, these 52 accessions were distributed into three main groups with high heterogeneity among the materials, but with a higher association among the individuals of the same accession. In general, in both morphological and molecular characterizations, the six elite accessions appeared distributed in different groups, as well as the two plants that compose each accession.
Regarding the qualitative descriptors, the presence or absence of pubescence and anthocyanins in the stem, petiole, abaxial surface of the leaf, bract, androgynophore, base of the ovary, and the mature fruit are the most influential descriptors and separate accessions. The accessions in the dendrogram showed a similar distribution to the results of the PCA, indicating that these characteristics are not systematically associated with groups of accessions according to their geographical origin. In general, 53% of the accessions were discriminated by the qualitative descriptors with the highest contribution to variability in the MCFA; some accessions that possess these qualitative characteristics of high heritability showed tolerance to thrips and viruses (VaLFla07/09, TolFla07, CauFla04, AntFla02/06, Ecu_2, and MetFla01). This germplasm should also be considered for future studies of selection of genotypes with tolerance to biotic factors.
The genetic variability estimated in this study is higher than the molecular variability found by Ocampo et al. (2017); this high heterogeneity in the fruit variables is the result of cross-pollination between different genotypes in the same orchard. The dispersion of the accessions from Valle del Cauca (ValFla) in all dendrogram groups confirms that this department has been the source of seed dispersal for all the producing areas of the country since the 1950s, when yellow passion fruit was introduced to Colombia (Morton, 1967). The accessions of Ecuador (Ecu_1/2/3) and Brazil (Bra_1) did not show considerable differentiation, which is supported by the recent introductions of Brazilian germplasm in the last decade to Ecuador (Viera et al., 2020; personal observations). However, these new introductions do not show the desirable quality characteristics of the fruit for the concentrated juice export market, which demands fruits with >45% pulp and °Brix>14.0.
Current status and future prospects for breeding
In Colombia, yellow passion fruit was introduced to Valle del Cauca in the early 1950s from Hawaii through Venezuela. For nearly 70 years, producers have contributed to its domestication process with new agronomic practices. Despite this, there are no selected cultivars in Colombia available for the producers that meet the demands of the different fresh and processed consumption markets.
Colombian growers, for many generations, have carried out the massal selection of fruits from the best yellow passion fruit plants (Ocampo, Urrea, et al., 2013). However, growers do not distinguish different local varieties based on the morphology of fruit characters due to the high heterogeneity in fruit shape, size, and color within the orchards. This pomological variability is also supported by outcrossing pollination (Arias-Suárez et al., 2014) and by the exchange of seeds between producers in different areas (Ocampo et al., 2017).
Yellow passion fruit breeding programs in Colombia can benefit from the results obtained from the germplasm evaluation and characterization in this study, together with information reported by Ocampo et al. (2017) employing molecular markers. These studies establish considerable variability in the cultivated yellow passion fruit materials present in Colombia, structured in four and five clusters by the neighbor-joining analysis. Pre-breeding efforts can improve fruit quality and yield, by emphasizing the elite accessions identified. Elite selections with specific quality characteristics (TSS or PUL%) are of utmost importance for breeders establishing the current genetic structure and variability of cultivated yellow passion fruit for Colombia, in the search for more productive cultivars with resistance to specific phytosanitary problems, such as thrips, collar rot, and viruses that are spread across the country. These accessions must be evaluated in the field in other localities at different selection cycles to establish interesting traits with controlled pollinations to avoid gene flow with unwanted genotypes, through a multivariate analysis. An alternative for a yellow passion fruit breeding program in Colombia is employing the residual or restricted maximum likelihood/ best linear unbiased prediction (REML/BLUP) methodology aimed at the selection of progeny or individuals from recurrent selection cycles. This allows accumulating genetic gains in the traits of interest while maximizing heterosis to develop new cultivars (Santos et al., 2015; Viana et al., 2016; Silva et al, 2017).
Advances in phenotype and molecular genetics may overcome some of the limitations of the traditional breeding methods based on sexual recombination by increasing selection efficiency using high-throughput phenotyping (Moreira et al., 2020), molecular markers (genotyping by sequencing - GBS), and transgene technology. In these methods, individual genes, or the recent emergence of genome-editing technologies such as CRISPR/Cas9 (Limera et al., 2017; Fernie & Yan, 2019), have been a significant milestone in crop breeding. Breeding programs could implement all these strategies in Colombia. Grafting with relative species of P. edulis f. flavicarpa is another technology that must be considered to solve soil-borne diseases such as Fusarium solani (Ambrósio et al., 2018), one of the leading causes of economic losses in yellow passion fruit crops in Colombia (Osorio Cardona et al., 2020).
The selection of genotypes with abiotic stress tolerance used as rootstock could also be an effective strategy to overcome/mitigate constraints for the adequate development of yellow passion fruit orchards (Hurtado-Salazar et al., 2017; 2020). Additionally, all elite accessions identified in this study must be included to enrich the primary gene pool of the cultivated germplasm in Colombia as an ex situ conservation strategy in gene banks and to improve the genetic resources of this species for future research studies (Ocampo Pérez et al., 2021).
Conclusions
The agro-morphological characterization of 52 yellow passion fruit accessions was highly informative, showing important levels of variability in 11 qualitative and 19 quantitative descriptors. The genetic variability was structured in five main groups, with low genetic differentiation among different geographic origins of the accessions due to several factors, such as outcross pollination and seed exchange between growers in different regions. The results of the agronomic evaluation identified six elite accessions (ValFla05, ValFla09, ValFla12, AntFla04, CunFla02, and HuiFla06) that constitute a genetic reservoir for establishing a working collection for future selection and recombination cycles, with well-differentiated and agronomically outstanding parents for passion fruit breeding programs with a commercial focus in the country.

