











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkIntroduction
Food security is strongly influenced by the threat of climate change (CC), not only through global warming but also by altered rain patterns, which generate more extreme weather events (IPCC, 2019). The effects on fruit trees will probably intensify for countries in the tropics and subtropics with higher initial temperatures, which will affect more marginal or degraded lands and underdeveloped regions with a low adaptation potential (Yohannes, 2016). Thus, global warming will affect small farmers more, who, for the most part, depend solely on rain (Sthapit et al., 2012). In addition, the Andean tropics will be greatly affected by CC, not only because it would increase precipitation by 20% to 25% but also because warming would increase more at altitude than in valleys (Marengo et al., 2011).
Zandalinas et al. (2021) highlighted that multifactorial stress, generated by CC, severely affects the growth and survival of crops, for which Dubey et al. (2021) pointed out that the intensity and frequency of abiotic stress are constantly increasing. Chmielewski et al. (2008) stated that adaptations of fruit plantations to CC require time and long-term research.
One of the most important limitations for fruit growth is the presence of physiological disorders, especially fruit cracking (Fig. 1), which limits production and quality (Ramteke et al., 2017) both at harvest and post-harvest, mainly in berries and drupes. This disorder affects the appearance of fruits, increases water loss and susceptibility to infection by pathogens, and decreases storage and shelf-life, resulting in substantial commercial losses for growers (Ginzberg & Stern, 2016; Yu et al., 2020). The peel plays a crucial role in resistance to cracking, portability, storability, and quality during storage (shelf-life quality) (Wang et al., 2021).

FIGURE 1 Fruit cracking in different fruit species. A) cape gooseberry; B) sweet granadilla; C) feijoa; D) pear; E) banana passion fruit; F) lulo.
Cracked fruits lose their commercial value for the fresh market and can only be sold locally or for the processing and transformation, as reported by Rojas Alfonso et al. (2012) for cape gooseberry producers in Cienaga (Boyaca, Colombia), where the exporting company returns at least 20% of the fruits because of cracking. Likewise, in times of heavy and prolonged rainfall, exporters can reject up to 50% of cape gooseberry fruits because of cracking (Fischer, 2005). Interestingly, cracked pomegranate fruits developed a higher oil content than intact fruits (Zaouay et al., 2020).
For the main causes of this physiological disorder, Yilmaz and Özgüven (2019) mentioned genetic factors, very ripe fruits, and fluctuating soil moisture, apart from nutritional deficiencies, sunburn, injuries, and pathogens on the skin of the fruits. Generally, fruit cracking occurs because of a physical failure of the cuticle or skin as the result of tensions (stresses) and heavy rains (Ramteke et al., 2017). Because of fruit cracking, water management and nutrition in crops have become a concern for many countries (Fischer & Orduz-Rodríguez, 2012).
In nature, fruit cracking manifests at the end of fruit development, that is, just after ripening and before seed dispersal (Lichter et al., 2002). It is a physiological disorder that affects the exocarp and the mesocarp (Yu et al., 2020) and can be distinguished from epidermis cracking, which is more superficial, including the cuticle and epidermal tissue. Deeper cracking can be characterized by an opening to the interior of the pulp, known as splitting (Opara et al., 1997; Fischer, 2005). In the cape gooseberry, Gordillo et al. (2004) found two types of cracking: superficial (cracking) and deep (splitting) (Fig. 1A). However, in most cases, cracking and splitting are used synonymously (Lopez-Zaplana et al., 2020).
In the case of cherries, microcracking results in deterioration of the barrier, involving only the cuticle, whereas macrocracking involves both the barrier and structure, deteriorating the cuticle and the internal cell layers (Knoche & Lang, 2017).
The objective of this literature review was to report on advances in environmental, cultural, and physiological aspects of fruit cracking, which are important for making management decisions, and plant breeding programs to reduce and avoid this disorder in many fruit crops, especially in the context of climate change and variability.
Factors causing fruit cracking
Figure 2 shows these factors for the cape gooseberry (Physalis peruviana L.), a fruit that shows high percentages of cracking and various causes. Fruit cracking is generated by external factors, including agronomic and environmental, and internal factors (Yu et al., 2020). It is a physiological disorder that involves the exocarp and mesocarp of the fruit and affects its appearance, intensifies the loss of water and predisposition to infection by pathogens, and decreases the postharvest shelf-life, generating considerable losses for fruit growers (Ginzberg & Stern, 2016; Ramteke et al., 2017). Wang et al. (2021) characterized fruit cracking as a common physiological disorder, in which the surface cracks because of uncoordinated internal growth and an external environment with high climatic variability.
The incidence of cracking varies widely according to the climatic characteristics during fruit development, the different fruit species and varieties, growth sites, and crop management (Fischer & Orduz-Rodríguez, 2012). As these authors report, fruit cracking can occur occasionally in any season, garden plant, branch, or cultivar.
Environmental factors
Water
Cracking because of rain, especially in fleshy fruit, (Fig. 1) imposes a severe problem in production and is related to huge commercial losses worldwide (Grimm et al., 2019). This situation greatly affects the cultivation of fruit trees in the Andes, where rainfall will increase as a result of climate change (Marengo et al., 2011).
High amounts of rain for several days or short heavy rains after a dry season can cause fruit cracking, such as in the cape gooseberry (Fischer & Melgarejo, 2020; Fischer et al., 2021). There is a disproportion between the amount of water that enters a fruit in a given time and the extensibility of its epidermis (Fischer & Orduz-Rodríguez, 2012). However, this turgor model (the critical turgor hypothesis), initially developed for grapes (Vitis vinifera L.) by Considine and Kriedemann (1972), was questioned by Winkler et al. (2016) in sweet cherries (Prunus avium L.) because its results indicated that rain cracking is a localized phenomenon, a local exposure of a fruit's skin to water that is not related to the net water balance. According to Grimm et al. (2019), rain and fruit cracking have a causal relationship, and cracking is related to a change in the water relations of fruits, associated with the humidity of its surface. Also, abundant rains can cause very watery fruits, as in guava (Psidium guajava L.), with a reduced content of sugars, ascorbic acid, and titratable acidity, making them more susceptible to cracking (Fischer & Melgarejo, 2021).
The water balance of fruits is associated with several factors that can generate cracks (Saei et al., 2014), where the water potential is the force that produces cracking, while the cell wall and other structures must withstand this pressure (Lichter et al., 2002). Thus, the biomechanical characteristics of the epidermis are crucial in maintaining internal pressure and resistance to fruit cracking (Saei et al., 2014). In the case of cherries, Knoche (2015) pointed out that an important cause of cracking is the rapid absorption of water by a fruit, which can be generated by direct absorption of the skin of the fruit or absorption by the vascular system of the plant.

In berries, such as the tomato and cape gooseberry, the high-water content and the high concentration of solutes exert high pressure on the epidermis of fruits, which cannot resist and cracks (Peet, 2009; Fischer & Melgarejo, 2020). This situation is accentuated if there are few fruits on the plant, such as the first fruits when the cape gooseberry production cycle begins (Gordillo et al., 2004). In postharvest, this problem is also accentuated when cape gooseberry fruits are subjected to immersion treatments in water, which limits the use of technologies such as pre-cooling with water and disinfection with products in liquid form.
In the case of high-water availability in the soil, cracking is very frequent in combination with two other factors, high relative humidity (RH) and low air temperature, conditions that reduce the transpiration of fruits (Fischer, 2005). For the pomegranate, Ikram et al. (2020) mentioned environmental factors, such as the imbalance of soil and air humidity, that contribute to intense cracking, that is, factors that have a high influence on fruit growth and development.
Relative humidity and temperature
In several studies in apple, Opara et al. (1997) reported greater cracking as the result of a lower formation potential or change in the composition of the cuticle, losing protective capacity, and a high RH that decreases the loss of water through the fruit. Deep splitting occurred when there was noticeable and depressed transpiration for 6 hours or more (Opara et al., 1997). On the other hand, Fischer (2005) reported a propitious effect of fruit cracking from low night temperatures in combination with a high RH.
A high RH prevents transpiration, which generates a high pressure inside the fruit, so that the epidermis can crack (Fischer & Orduz-Rodríguez, 2012), especially during prolonged periods of a RH between 99% and 100%, alone or in combination with rain (Fischer, 2005).
In tropical species such as passion fruit (Passiflora ligularis Juss, Fig. 1B), temperatures below 12-15°C are conducive not only to floral abortion and lower fertilization but also to fruit cracking (Fischer & Miranda, 2021). In addition, this species cracks more when sudden changes occur between day and night temperatures (Miranda Lasprilla, 2020). Cape gooseberries are susceptible to cracking from high amounts of water (Figs. 1A and 2) when the fruit grows completely inside the closed calyx, which dampens extreme temperatures by up to 5°C, especially with variations in day and night temperatures (Fischer et al., 2021).
In the cherry, apple, and peach, Bohlmann (1962) observed a tendency to crack when the temperature of the water on the epidermis increased. Reddy et al. (2017) also pointed out that physiological disorders are accentuated by global warming, such as the cracking of lychee fruits. Likewise, in hotter and more humid weather that follows a cold and dry period, there is danger of cracking, especially when young fruits begin to fill (Opara et al., 1997).
For the pomegranate, an excessive increase in temperature, hot dry winds, a downpour after a dry season, and large differences between day-night temperatures with temperatures greater than 38°C combined with a RH of 60%, favor fruit cracking in this species (Ikram et al., 2020).
Light
Regarding light, the side of the fruit exposed to direct sun can crack more because the epidermal cells on this exposed side become comparatively thicker and inelastic and do not adapt to the rapid increase of the tissues below (Opara et al., 1997). Ikram et al. (2020) reported that direct sun increases the temperature and evapotranspiration of the fruit surface, which results in a high moisture loss and a greater susceptibility to cracking.
Opara et al. (1997) pointed out that, in some apple varieties, cracking occurs preferably on the shady side of the apples, i.e., the phenomenon is exclusively varietal. Fischer (2000) observed greater crack at 2,700 m a.s.l. in the department of Cundinamarca (Colombia) in 'Jona-gold' apples exposed directly to the sun and affected by Venturia inaequalis.
Ulinnuha et al. (2020) found a lower incidence of cracking in tomatoes under shade and proposed further investigation of this phenomenon with associated crops that produce shade. Suzuki et al. (2007) harvested more fruits without cracking in this species when the foliage and fruits intercepted a lower amount of light than with other types of conduction (training).
Mineral nutrition
The mineral nutrition of fruit is highly related to cracking (Yu et al., 2020). Ca deficiency has been reported as one of the main causes of cracking, such as in mandarin (Chabbal et al., 2020), lemon (Devi et al., 2018), grape (Yu et al., 2020), fig (Aydin & Kaptan, 2015), sweet cherry (Erogul, 2014), sour cherry (Simon et al., 2007), apricot (Nie et al., 2017), pomegranate (Davarpanah et al., 2018), litchi (Martínez Bolaños et al., 2017), loquat (Yilmaz, 2020), sweet granadilla (Fischer & Miranda, 2021; Fig. 1B), and cape gooseberry (Cooman et al., 2005; Fig. 1A).
As a macroelement and the most important element for fruit quality (Marschner, 2012), Ca is essential for multiple important functions in plant physiology, mainly because of its role in the structural integrity and stability of cell walls and the middle lamella, not only serving as a bridge between pectin molecules but also improving the integrity of cell membranes through the bonds with phospholipids. Additionally, it participates in stress signaling as a secondary messenger (Ranty et al., 2016; Yu et al., 2020).
Gordillo et al. (2004) observed that a deficient dose of potassium (K), as well as a lack of boron (B) in fertilization, induced greater cracking in cape gooseberry fruits. Cooman et al. (2005) also found an increase between 5.5% and 13.0% in the number of cracked fruits in this species when Ca or B was lacking in fertilization (Fig. 2). Fischer et al. (2020) reported cracking in banana passion fruit, known in the high tropics of Colombia as "the witch's laugh" (risa de bruja in Spanish) (Fig. 1E) because of the visibility of all seeds, attributed to Ca and/or B deficiency. Marschner (2012) underlined the role of B in the structure of cell walls.
On the other hand, Garzón-Acosta et al. (2014) recorded the highest cracking in mature and harvested cape gooseberry fruits in a greenhouse in plants that suffered from a magnesium (Mg) deficiency (Fig. 2), compared to plants deficient in Ca (11% and 1% cracked fruits, respectively). Problems with fruit cracking in feijoa (Acca sellowiana;Fig. 1C) have also been reported in Colombia with a Ca/Mg ratio >10 in the soil, where fruits crack and develop brown pigmentation (Fischer et al., 2021). Marschner (2012) stated that part of Mg is firmly bound to pectin in cell walls; therefore, deficiency of this element would contribute to cracking.
On the contrary, high concentrations of N in the soil from chemical or organic overfertilization, especially when the content of organic matter (OM) in the soil exceeded 20%, increased cracking in the cape gooseberry (Fig. 2) (Gordillo et al., 2004). Fischer and Orduz-Rodríguez (2012) suggested that the increase in the epidermis is not enough to keep up with the expansion of the fruit. Similarly, Yilmaz (2020) found an overconcentration of N in cracked loquat fruits.
Phytosanitary problems
Superficially or deeply cracked fruits not only exhibit open lesions that facilitate a loss of moisture and wrinkling but also are susceptible to infection by pathogens (Opara et al., 1997), which Fischer (2005) described in cracked cape gooseberries affected by Botrytis.
In apples cultivated in the high tropics, the epidermis of the fruit can be burned and cracked by high insolation (sun burning), especially by high amounts of UV light, which facilitates infection by Venturia inaequalis (Fischer, 2000). Casierra-Posada (2012) pointed out that B deficiencies can weaken the epidermis of young apples and pears, facilitating infection by V. inaequalis and V. pyrina, respectively, that later show lesions similar to cracking.
Ikram et al. (2020) and Singh et al. (2020) reported that attacks by insects, pathogens, and birds especially affect ripe, cracked fruits, which greatly reduces their commercial quality.
Another cause of cracking or aggravation in existing lesions may be the application of agrochemicals, especially mixtures with surfactants since they increase the penetration of water through the cuticle of the fruit (Opara et al., 1997).
Fruit factors
The relationships between fruit characteristics and susceptibility to cracking have also been evidenced (Fig. 2). Khadivi-Khub (2015) have reported a series of correlations between cracking and different aspects of the fruit, such as size, shape, and firmness, and the epidermis (resistance, stomata, and cuticular properties), in addition to the osmotic concentration, water capacity of the pulp, and the state of development of the fruit.
In the 'Terigas' mandarin, Hardiyanto and Nirmala (2019) found that fruit cracking increased with decreases in the thickness of the epidermis and, surprisingly, thinning of the peel of the mandarins was not influenced by nutrients (K, Ca, Mg).
In most species, the fruit suffers constant stress during development, because the volume is constantly increasing and, therefore, the surface of the fruit increases as well (Knoche & Lang, 2017). These authors characterized the skin of fruits as a primary structure, consisting of a polymeric cuticle whose function is similar to that of a barrier that covers the epidermis and hypodermis, cell layers that are responsible for the skin's load-bearing functions.
Mishra et al. (2016) in pomegranate (Punica granatum L.) and Fischer (2005) in cape gooseberry reported that a prolonged drought hardens the skin (epidermis) prematurely, and the next heavy rain causes the pulp to expand. The same occurs during phases of rapid fruit growth, in which the skin cracks, especially in late harvests of mature fruits (Fig. 2) (Mishra et al., 2016). The parts of the fruit that are most susceptible to cracking are its weak parts, as is the case with lenticels (Fischer, 2005).
In the pomegranate, during water stress, the growth of the peel is limited, as in the aril; however, when the water enters the fruit again, the aril expands more than the peel, which results in fruit cracking (Ikram et al., 2020). This desynchronization when the pulp turgor exceeds a critical threshold causes the fruit skin to stretch beyond its extensibility limit and crack (Considine & Kriedemann, 1972).
In most studies, an increase in the incidence of cracking was found as fruit maturation advanced (Fig. 2). Jiang et al. (2019) and Wang et al. (2021) stated that, when the fruit is in an advanced state of maturity and a downpour occurs, the risk of cracking is much higher. In addition, fruits in a state of overmaturity are more susceptible to cracking because of the senescence of the epidermis (Fischer & Orduz-Rodríguez, 2012).
Gordillo et al. (2004) found in the cape gooseberry that this physiological disorder occurs especially in fruits with a large volume and weight and at the beginning of the harvest season (Figs. 1A and 2), considering that this species has indeterminate growth (Fischer et al., 2021), in which vegetative and reproductive development occur at the same time. This situation was described by Torres Azuero et al. (2016) as an imbalance between the source-demand relationship because of an excess of assimilates in the first stage of the harvest, which supposedly generates pressure from assimilates, propitiating an average of 15% of cracked fruits in the cultivation of the cape gooseberry in the Cundiboyacense highlands. Also, Alvarez-Herrera et al. (2012) found the highest cracking (81%) in large cape gooseberry fruits (Fig. 2) but only 33% in small fruits in the same greenhouse study; the highest percentage of cracked fruits was observed in the upper stratum (38%) (Fig. 2), as compared to only 25% in the lower stratum of the plant.
In different citrus cultivars, most of the cracking occurs during cell elongation or in the period of fruit maturation. However, in some cultivars, cracking can occur throughout the development of the fruit (Li & Chen, 2017).
In tomato studies, Jiang et al. (2019) concluded that ripe fruit is more resistant to cracking as it contains more intact pectins as a result of a good degree of binding between the pectins by Ca2+ and other cross-linkages. Additionally, the correlation analysis by these authors verified that tomato cracking is significantly more related to the level of proto-pectin and cellulose than to that of Ca2+.
On the other hand, a transcriptome analysis of atemoya (Annona cherimola x A. squamosa) by Chen et al. (2019) found that the decomposition of starch into soluble sugars and the metabolism of polysaccharides of the cell wall are closely related to fruit cracking. The hydrolytic enzyme of the cell wall, polygalacturonase (PG), breaks the α-(1-4)-galacturonan bond in the pectin chain, so an increase in PG activity generates fruit softening (Lu & Lin, 2011). On the other hand, research on the non-enzymatic protein expansin (EXP), which is involved in the extension of cell walls, is essential for knowledge on causes of cracking (Balbontín et al, 2013) since varieties that are more resistant to cherry and lychee splitting have a level of EXP expression that was noticeably higher than in those most susceptible to this physiological disorder (Balbontín et al, 2014).
Brüggenwirth and Knoche (2017) and Jiang et al. (2019) noted that factors such as disassembly, modification, and composition of the cell wall can affect the mechanical characteristics of the pericarp; therefore, they become determining elements of susceptibility to fruit cracking.
Varietal and genetic factors
As described above, the causes of cracking can be diverse, and the reasons vary between different species (Lu & Lin, 2011) and, in many cases, also between varieties of the same species (Opara et al., 1997). Khadivi-Khub (2015) pointed out that fruit cracking is a quantitative trait that is controlled by several genes. Additionally, this author indicated that the cultivars most resistant to this problem and that show adequate fruit quality can be chosen for subsequent cultivation. Therefore, cultivars that exert greater skin break force and elasticity are more resistant to fruit cracking (Brüggenwirth & Knoche, 2016).
When evaluating 54 cape gooseberry materials from northeastern Colombia, Herrera et al. (2011) found large differences in the occurrence of cracking, which was higher in feral cultivars with an average of 23.9%. However, within this group, there was an accession (06Uch0001) without broken fruits, as compared to accession 06Uch0073 with 50% cracked fruits. This result confirmed that cracking depends to a high degree on the genetics of the material (Fischer, 2005), as in other nightshade berries, such as the tomato (Peet, 2009). Criollo et al. (2014) compared cracking in three genotypes of cape gooseberry and found that the highest percentages of this disorder occurred in the materials 'Silvania' and 'Kenya' (8.9% and 8.1%, respectively), compared to 'Regional Nariño' with only 3.1%. These results confirmed that, under the same climatic conditions, fruits of different cultivars of the same species show differences in susceptibility to cracking (Khadivi-Khub, 2015). On the other hand, Lagos-Burbano et al. (2021) characterized the fruit of 36 hybrids obtained from double haploid lines, with cracking being one of the three parameters that explained the total variability through principal component analysis. They also reported that two of the six clusters registered a high seed content, a low maturity index, and a high percentage of cracking.
Berries are particularly susceptible to splitting (Fischer et al., 2021). However, Medina et al. (2009) reported that improved material from 'Lulo de la Selva' was used to find materials more resistant to cracking in the cultivation of lulo (Solanum quitoense Lam.) (Fig. 1F), and, after a generation of open pollination, the Jalisco clone showed reduced cracking and large fruits. Fischer and Melgarejo (2020) pointed out that plant breeding programs should also include resistance to cracking, and not only the production of large fruits, because size is highly correlated with the number of seeds (Trevisani et al., 2017; Balaguera-López et al., 2020). This should always be considered in these programs because cracking is an attribute whose gene expression may also depend on environmental conditions.
It is worth mentioning that the development of high-quality varieties resistant to cracking and the identification of genes involved in this resistance should be an important objective in plant breeding programs in fruit trees (Balbontín et al, 2013).
Examples for cracking handling
The best way to reduce fruit cracking today would be proper orchard management that tries to minimize water stress and considers nutrition and physiological factors that contribute to fruit cracking (Khadivi-Khub, 2015).
Table 1 shows examples of Ca applications in cultivation, alone or in combination with B or K, which resulted in a decrease in the incidence of cracking in grapes, citrus, and cape gooseberries. Cronjé et al. (2013) suggested an interaction of Ca with B, forming a stabilizing complex in the middle lamella of cells. For these effects, Yu et al. (2020) carried out applications of Ca during the maturation phase to prevent a localized deficiency of this element since it is necessary for fruit quality. Also, in the cape gooseberry, B or K alone (Tab. 1) reduced the percentage of fruit cracking (Sabino-López et al., 2018). In the pomegranate fruits highly susceptible to cracking with losses of up to 40-60% (Ikram et al., 2020), 0.2% B reduced the presence of this physiological disorder (Sharma & Belsare, 2011).
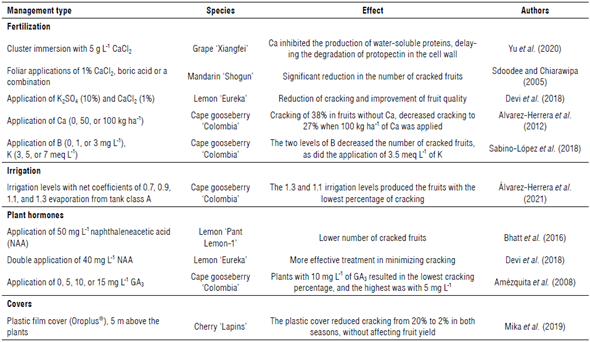
Applications of naphthaleneacetic acid have shown good results for controlling cracking in lemons (Tab. 1), which Devi et al. (2018) attributed to the significant role of this plant hormone in increasing resistance and plasticity of the peel, which determine the intensity of cracking. Furthermore, Amiri et al. (2012) specified that synthetic auxins decrease cracking in citrus fruits because of the increased thickness of the rind. In the pomegranate, an application of 5 mg L-1 of forchlorfenuron (CPPU), a cytokinin, significantly reduced cracking (Sahu & Sharma, 2019), as did applications with paclobutrazol (an anti-gibberellic compound) (Khalil & Aly, 2013). The results of the application of gibberellins in cultivation are not consistent (Amézquita et al., 2008), possibly because this regulator can increase the size of fruits. However, Amézquita et al. (2008) found that applications of 10 mg L-1 of GA3 generated the lowest percentage of cracking of cape gooseberry fruits (Tab. 1; Fig. 2).
Besides proper management with nutrition, irrigation, and growth regulators, Ikram et al. (2020) reported other practices to reduce cracking for the pomegranate, such as protecting the fruits from direct solar radiation by bagging or putting a shade net, as well as the application of antitranspirants based on kaolin.
Additionally, in different crops, specific management methods should be sought, such as the elimination of the first flowers at the beginning of the productive cycle in the cape gooseberry (Fischer & Miranda, 2012). Also, in the cape gooseberry 'Silvania', cracking was reduced by pruning (training) with an increase in primary branches, from three to four (Criollo et al., 2014). By increasing the number of branches, the plants had a greater transpiratory volume, an increase in cell mass, and a better water distribution (Marsal et al., 2006). Criollo et al. (2014) assumed that they reduced the risk of breaking the fruit surface because of a decrease in pressure exerted on the cell walls at the pericarp level.
Conclusions
Fruit cracking is a physiological disorder that seriously affects the quality and marketability of many fruit species.
Because of climate change, this physiological disorder is aggravated in areas where the intensity of rains is higher, but also in regions where there is an increase in temperatures and solar radiation and in the dry seasons that will be followed by rains, which is the main cause of this disorder. In general, fruit cracking is due to the stated climatic conditions, but also due to prolonged rains in combination with a high RH that suppress the transpiration of fruits.
High temperatures and/or sunburn age the epidermis of fruits, which cannot withstand the pressure when the fruit fills up with more turgor again. Likewise, sudden changes between daytime and nighttime temperatures or hot seasons after cold ones promote cracking in several species.
The deficiency of nutrients that are important for the stability of the cell wall, such as Ca, B, and Mg, is another important cause of this physiological disorder. Additionally, the lack of K that regulates the water rate in plants, influences cracking. Likewise, excess N can increase the pressure of the pulp on the epidermis of the fruit, causing it to crack.
Measures are proposed to control this disorder, such as the selection of more resistant varieties, adequate nutrition to strengthen the resistance of the cell wall, the avoidance of water stress in plantations, and the application of growth regulators such as NAA or GA3, depending on the species. Additionally, plastic covers should be placed on species, such as cherries, that easily crack as a result of water absorption on the skin of the fruit. Specific tasks are important according to the species, such as eliminating the first flowers or increasing the number of primary branches in the cape gooseberry to reduce the pressure of water and solutes on a small number of fruits when the plant is in full growth and just beginning production.
Finally, since many fruit species lack successful research on fruit cracking, this literature review provides initiatives to lessen this problem with proper crop management and breeding programs for the affected species and varieties that consider the aforementioned traits.

