











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkINTRODUCTION
Magnoliaceae Juss. comprises approximately 300 species distributed mainly in tropical and subtropical areas of Asia and America (Rivers et al. 2016). The family is characterized by trees and shrubs with an annular scar around the nodes and floral organs spirally arranged. Most species have bisexual flowers except for a few Asian species with unisexual flowers (Chen & Nooteboom 2003). Two monophyletic subfamilies have been well recognized: Liriodendroideae and Magnolioideae (Azuma et al. 2001). However, the classification of their members has been very controversial (Azuma et al. 1999, 2001, Dandy 1927, 1978, Chen & Nooteboom 1993, Figlar & Nooteboom 2004, Frodin & Govaerts 1996, Keng 1978, Kim et al. 2001, 2002, Law 1984, Li & Conran 2003, Lozano 1975, 1983, Nooteboom 1984, 1985, 1987, 1993, 2000, Vázquez-García 1994).
According to the most recent classification of neotropical species based on morphological and molecular studies, Magnolioideae comprises only the genus Magnolia L. and the other genera are currently considered sections and subsections of this genus (Figlar & Nooteboom 2004). Currently, all the Colombian species are included in Section Talauma (Figlar & Nooteboom 2004). However, these species belong to two different subsections based on morphological characters: Talauma Juss. and Dugandiodendron Lozano (Figlar & Nooteboom 2004, Lozano 1975, 1983, 1994).
Dugandiodendron, with 14 species restricted to Colombia, northern Ecuador, and eastern Venezuela, was initially described at generic rank by Lozano (1975) based mainly on two morphological characters: heliciconvolute prefoliaton and the pseudoaxillary position of the flowers. Although the characters used for generic circumscription have been questioned (Nooteboom 1985), the monophyly of Dugandiodendron has been suggested based on fruit dehiscence and stamen morphology (Serna 2005).
Several morphological characters still need to be explored in order to clarify the systematics of Colombian magnolias. However, these species are difficult to study because of their rarity, restricted geographical distribution and high level threats to their survival. For all these reasons, Magnoliaceae was chosen as a research and conservation priority in Colombia in its National Strategy for Plant Conservation (Samper & García 2001).
In light of the controversy about the taxonomy of Magnoliaceae and the paucity of phylogenetically informative characters (Nie et al. 2008), a study of pollen morphology was undertaken in order to contribute additional character data. Magnoliaceae pollen is stenopalynous in shape and aperture type. Some variations are common in the sporoderm structure, sculpture and size (Praglowsi 1974). Agababian (1972) described pollen grains with a fine granular exine and perforated tectum. Similarly, Praglowski (1974) considered that most pollen grains in this family have a perforated or slightly rugulate tectum,and occasionally a microreticulate exine. Other recent studies have described several ornamentation types in Asian species (Xu 1999, Xu & Kirchoff 2008). In Colombia, pollen grains of Magnoliaceae have been described as monocolpate, prolate and subprolate. Smooth pollen grains with few and scattered perforations have been described in Talauma species, while Dugandiodendron pollen grains have been described as microreticulate or reticulate (Lozano 1975).
Although Lozano (1975) provides some information fo r a limited sample of colombian species, the pollen morphology of neotropical magnolias is still largely unknown. The goal of this study was to describe the pollen grains of a broader sample of Magnolia species from Colombia and evaluate the contribution of pollen morphology to the classification and systematics of Colombian magnolias. To characterize the pollen grains, a description of size, shape and ornamentation was carried out on available material of fourteen species obtained from herbaria and field collections.
MATERIALS AND METHODS
Pollen grains of Magnoliaceae species from Colombia were collected from herbarium specimens deposited in COL and JAUM (Table 1). Material fixed in alcohol was also available for some species from Antioquia department. Pollen grains were subjected to standard acetolysis standard (Erdtman 1960) in the laboratory of Paleoecology at the National University of Colombia, Medellin. Light microscopic analyses were performed using an Olympus BMX 40 microscope, and SEM analyses were performed using a JEOL JSM 5910 LD Scanning Electron Microscope. The accelerating voltage was 15 kV.
Table 1 Voucher specimens deposited in the laboratory of Paleoecology (LP), Universidad Nacional de Colombia, Medellín.
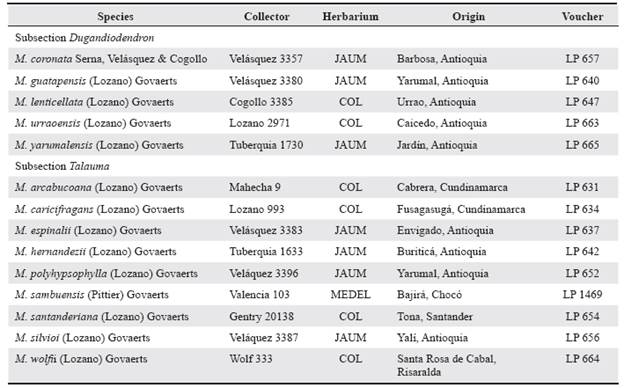
For each species, at least 25 pollen grains were measured under light microscopy at 100X with an Olympus OMS4 micrometer, except in Magnolia silvioi (Lozano) Govaerts and Magnolia sambuensis (Pittier) Govaerts, which were measured at 40X because of their size. Exine was always measured at 100X. After acetolysis, the pollen voucher specimens were mounted on slides with glycerine and deposited in the collection of the laboratory of paleoecology of the Universidad Nacional de Colombia, Medellín (Table 1). The terminology of Punt et al. (2007) and Zhang et al. (2014) has been adopted here to describe pollen morphological characters, except those related to pollen grain shape, for which terminology was adopted from Walker and Doyle (1975). The shape of pollen grains was described as the product of division of the longest axis (LA) by the shortest equatorial axis (SEA) (Zhang et al. 2014).
Pollen type was defined according the classification proposed by Praglowski (1974) based on tectum surface and grain size: large pollen grains, 45 [im or larger, smooth tectum surface (Type 1); small pollen grains, <45 μm, smooth tectum surface (Type 2); large pollen grains, sculpture slightly rugulate (Type 3); small pollen grains, sculpture slightly rugulate (Type 4), sculpture coarsely rugulate (Type 5); pollen grains semitectate, exine structure microreticulate, large (Type 6) and pollen grains semitectate, exine structure microreticulate, small (Type 7).
RESULTS
The species studied are listed in Table 1. All the pollen grains were anasulcate, bilateral symmetric, heteropolar, and folded in boat shape. In polar view, the pollen grains were ellipsoidal and in lateral view, biconvex or plane-convex. The size of the pollen grains varied from 68.20 ± 7.1 μm to 115.0 ± 7.7 μm in the longest axis (LA) and 41.4 ± 2.3 μm m to 69.3 ± 10.0 μm in the shortest equatorial axis (SEA) (n=25), and are considered large or medium in size. Three types of pollen shapes in the species analyzed were found: prolate, perprolate and subprolate. The ornamentation was also variable: psilate-perforate, slightly rugulate, rugulate and rugulate-perforate (Figs. 1-2). Measurements and ornamentation information for each species are shown in Table 2.
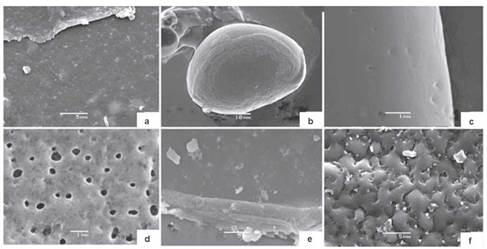
Figure 1 Scanning Electron Microscopy images of pollen grains from Subsection Dugandiodendron: a. Magnolia coronata, b. M. urraoensis. Subsection Talauma: c. M. arcabucoana, d. M. espinalii, e. M. hernandezii, and f. M. silvioi.
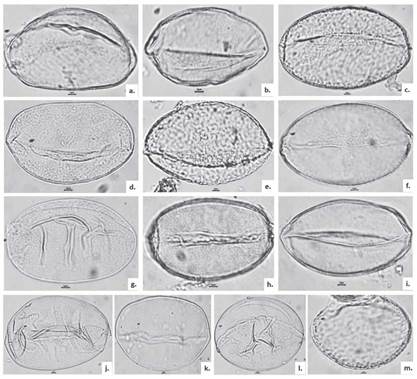
Figure 2 Light Microscopy images of pollen grains of the species studied from Subsections Dugandiodendron (D) and Talauma (T): a. M. arcabucoana (T), b. M. caricifragans (T), c. M. coronata (D), d. M. espinalii (T), e. M. guatapensis (D), f. M. hernandezii (T), g. M. lenticellata (D), h. M. polyhypsophylla (T), i. M. santanderiana (T), j. M. silvioi (T), k. M. urraoensis (D), l. M. wolfii, (T) and m. M. yarumalensis (D).
Table 2 Pollen morphological features of the species of Magnolia (Subsections Dugandiodendron and Talauma) examined from Colombia.
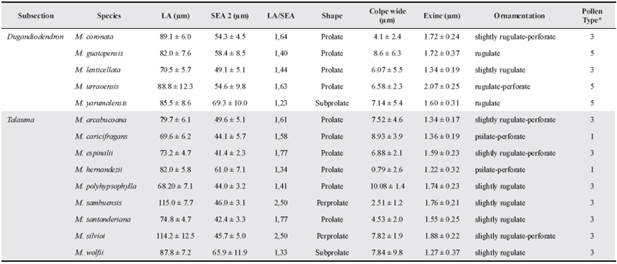
Abbreviations: LA = longest axis; SEA = shortest equatorial axis. *According to Praglowski (1974).
The pollen grain size was proportional to the flower size, especially for the species with large flowers. For instance, grains from M. silvioi and M. sambuensis were the largest, corresponding to the largest flowers of the Magnoliaceae in Colombia (tepals 7.5-8.5 cm long x 4.0-4.5 cm wide and 5.0-6.5 cm long x 4.0-4.5 cm wide, respectively). The smallest pollen grains were found in the species M. caricifragans and M. polyhypsophylla (3.5-4.5 cm long x 1.5-2.5 cm wide and 2.0-2.3 cm long x 1.5-2.6 cm wide, respectively). However, the smallest flowers among the Colombian magnolias are found in M. coronata (2.5 - 3.0 cm long and 1.0 - 1.5 cm wide).
Two general types of ornamentation were observed: rugulate and psilate with perforations. In the rugulate type, some variations were found: slightly rugulate, rugulate and rugulate with perforations. In this study, based on size and type of ornamentation, the pollen grains correspond to three out of the seven pollen types according to Praglowski (1974) (Table 2).
Pollen grains of the Subsection Dugan-diodendron were considered large, varying from 70.5 ± 5.7 to 89.1 ± 6.0 in the LA and 49.1 ± 5.1 to 69.3 ± 10.0 in their SEA. Based on sculpture, pollen grains were Type 3 and 5: pollen grains slightly rugulate and rugulate with or without perforations (Table 2).
For Subsection Talauma, pollen grains were considered large. varying from 68.2 ± 7.1 to 115.0 ± 7.7 in the LA and 41.4 ± 2.3 to 65.9 ± 11.9 in the SEA. Two general types of ornamentation were found: psilate-perforate (2 species) and rugulate (8). In the rugulate type, two subtypes of grains were observed: slightly rugulate (4) and slightly rugulate-perforate (4).
DISCUSSION
The main pollen characters described in this work such as sculpture, structure, and shape, were similar to the information previously published for species of Magnoliaceae (Praglowski 1974, Xu & Kirchoff 2008). The sulcate aperture, related to heteropolarity in Magnoliaceae, was found in all species analyzed. All the observed grains were large, corresponding to Praglowski's (1974) Types 1, 3 or 5. In this study, the dominant shape was boat-shaped, which is characteristic of Magnoliaceae (Xu & Kirchoff 2008). Boat-shaped elliptic, boatshaped oblong and boat-shaped elongate variations were also found in all Colombian species.
Other studies have described several ornamentation types in Asian species in Magnoliaceae as follows: foveolate in Michelia L. (Xu 1999, Xu & Kirchoff 2008); microperforate in Alcimandra Dandy, Manglietia Blume, Michelia (Xu & Kirchoff 2008), Lirianthe Spach, Houpoëa N. H. Xia & C. Y. Wu, Parakmeria Hu and Cheng and Talauma (Zhang et al. 2014); perforate in some species of Manglietia, Michelia and Lirianthe (Zhang et al. 2014); rugulate in Woonyoungia Law (Zhang et al. 2014); coarsely rugulate in Kmeria (Pierre) Dandy (Xu & Kirchoff 2008), and regulate in Michelia and with elaborated projections in Talauma (Zhang et al. 2014). In the case of Magnolia grandiflora L., Xu & Kirchoff (2008) described the pollen grains as microperforate, while Zhang et al. (2014) described the pollen grains as perforate and microperforate. These results show the high variability among species of the same genus. The results presented here demonstrate the high variability among species from the same subsection.
Lozano (1983) characterized pollen grains of four Colombian magnolias. He described the pollen grains of these species as monocolpate, prolate and subprolate. Apparently, he found some differences in the exine sculpture between Talauma and Dugandiodendron, and reported grains of Subsection Talauma species such as M. caricifragans and M. santanderiana as smooth with small and scattered perforations. On the contrary, he described M. argyrothricha and M. mahechae from Subsection Dugandiodendron as having microreticulate or reticulate pollen grains.
Later, Lozano (1994) described pollen grains of Magnoliaceae as ellipsoidal with rounded ends and classified the species based on Praglowski's types (1974). Magnolia calophylla (Lozano) Govaerts, M. caricifragans and M. santanderiana, were described as Type 2. Pollen grains of M. mahechae and M. urraoense were classified as Type 4, while Magnolia argyrothricha (Lozano) Govaerts, Magnolia colombiana (Little) Govaerts and M. yarumalensis grains belonged to Type 5. Our results concur with those of Lozano (1983, 1994) for M. yarumalensis but not for M. caricifragans (Type 2 in Lozano vs. Type 1 in this work), M. santanderiana (Type 2 vs. Type 3) and M. urraoensis (Type 4 vs. Type 5) (Table 2). These differences can be explained by the maturity level of grains, considering that Lozano (1983) observed immature pollen.
According to this study, two species from Subsection Talauma presented Type 1, the same type reported in species from the West Indies, United States and South America (Praglowski 1974). Seven species from Subsection Talauma and three species from Subsection Dugandiodendron, exhibited the same pollen type as M. grandiflora (Type 3), while three species from Subsection Dugandiodendron share the same type as Liriodendron tulipifera L. (Type 5), native species from North America.
The Colombian species from Subsection Dugandiondendron and Subsection Talauma (Section Talauma) share similar morphological patterns. Despite morphological characters such as exine structure, sculpture and pollen size that have some taxonomic significance, it is not possible to establish clear differences between these groups (Praglowski 1974, Nooteboom 1984).
The shape size, ornamentation, polarity and symmetry of pollen in colombian species of Magnolia are consistent with descriptions of pollen previously described in Asian and American species. Although some variability in pollen shape, ornamentation, and size was observed among the species examined in this study, the variability does not correspond to the previous taxonomic classification. For instance, Type 3 pollen grains were found in both subsections. The uniformity of pollen grain morphology among the Colombian species supports the classification proposed by Figlar and Nooteboom (2004) for neotropical Magnoliaceae, in which all Colombian species are included in Section Talauma.
The scarcity of available material and the acetolysis process limited the number of species and the total amount of pollen that could be sampled to obtain good photographic evidence for this study. Future studies could include increased sampling from herbarium collections and ideally new field work, in order to characterize the pollen of the remaining species.
AUTHOR CONTRIBUTIONS
MSG collected material from field and herbarium specimens, performed laboratory work and measurements. MSG and CVR described the pollen grain shape and ornamentation, obtained photographs and wrote the draft of the manuscript. Both authors improved the final manuscript based on the reviewers' comments and suggestions.

