Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkColombia Forestal
versión impresa ISSN 0120-0739
Colomb. for. vol.19 no.2 Bogotá jul./dic. 2016
https://doi.org/10.14483/udistrital.jour.colomb.for.2016.2.a05
Artículo de investigación
DINÁMICA DE BOSQUES EN DIFERENTES ESCENARIOS DE TALA SELECTIVA EN EL MAGDALENA MEDIO (COLOMBIA)
Forest dynamics under different levels of selective logging in the middle Magdalena River (Colombia)
Isabel C. Restrepo1, Ana M. Aldana2 & Pablo R. Stevenson3
1 Laboratorio de ecología de bosques tropicales y primatología (LEBTYP), Universidad de Los Andes, Bogotá, Colombia. ic.restrepo1444@uniandes.edu.co. Autor para correspondencia.
2 Laboratorio de ecología de bosques tropicales y primatología (LEBTYP), Universidad de Los Andes, Bogotá, Colombia. am.aldana262@uniandes.edu.co.
3 Laboratorio de ecología de bosques tropicales y primatología (LEBTYP), Universidad de Los Andes, Bogotá, Colombia. pstevens@uniandes.edu.co.
Restrepo, I.C, Aldana, A.M. & Stevenson, P.R. (2016). Dinámica de bosques en diferentes escenarios de tala selectiva en el Magdalena medio (Colombia). Colombia Forestal, 19(2), 195-208.
Recepción: 31 de agosto de 2015 / Aprobación: 8 de febrero de 2016
RESUMEN
La tala selectiva es una actividad de uso forestal utilizada con frecuencia, la cual ha demostrado tener un menor impacto sobre la biodiversidad que la tala generalizada. Sin embargo, tanto la magnitud como la dirección del cambio ecológico después de la tala dependen de su intensidad y de la subsecuente dinámica del bosque. Por esto, es importante realizar estudios que permitan comprender el funcionamiento de diferentes ecosistemas tras practicar tala selectiva. En este estudio se analizó la dinámica del bosque de la reserva El Paujil (Magdalena medio, Colombia) en términos demográficos, de regeneración, dinámica de claros, acumulación de biomasa y composición florística al comparar dos parcelas de una hectárea en un fragmento del bosque poco perturbado (primario y dos parcelas de una hectárea en un fragmento del bosque que fue objeto de tala selectiva en el pasado. Como se esperaba, la estructura del bosque y la acumulación de biomasa se ven alteradas por efecto de la tala selectiva, sin embargo, no generó un impacto significativo en los demás aspectos mencionados ya que, al parecer, las fuertes pendientes de la zona provocan una elevada mortalidad y promueven la formación de claros tanto en bosques intervenidos como en los que han sido poco alterados.
Palabras clave: Biomasa aérea, bosque primario, composición florística, demografía, serranía de las Quinchas.
ABSTRACT
Selective logging is a growing forestry activity with less impact on biodiversity than complete deforestation. Despite this, both the magnitude and the direction of ecological change after logging, strongly depend on the intensity, and subsequent temporal forest dynamics. For this reason, it is important to conduct studies to understand the dynamics of different ecosystems after selective logging. We analyzed forest dynamics at El Paujil Reserve (middle Magdalena River, Colombia), in terms of demography, gap dynamics, changes in floristic composition, and ability to gather biomass, comparing two 1 ha plots in a fragment of undisturbed “primary” forest, and two 1 ha plots in a fragment of forest that was subject to selective logging in the past. As expected, forest structure and biomass accumulation were altered as a result of selective logging; however, it did not generate a significant impact on the other aspects evaluated. We suggest that perhaps local steep slopes generate high mortality and gap formation in both forest types.
Keywords: Aboveground biomass, primary forest, floristic composition, demographics, Serranía de las Quinchas.
INTRODUCCIÓN
La tala selectiva en bosques tropicales es una actividad creciente que ha demostrado producir menos daños en el suelo y el dosel frente a la deforestación generalizada (Asner et al., 2004). Además, es la actividad de uso forestal con menor impacto sobre la diversidad, pues los bosques talados selectivamente presentan una alta riqueza de especies de bosque maduro (Gibson et al., 2011; Norden et al., 2009). Sin embargo, tanto la magnitud como la dirección del cambio ecológico después de la tala dependen fuertemente de su intensidad y de la subsecuente dinámica espacial y temporal del bosque afectado (Asner et al., 2004).
Entre las consecuencias más importantes que tiene la tala selectiva en los bosques está la pérdida de biomasa, no solo durante el periodo de extracción, sino también en el tiempo después de la actividad. En estudios realizados en el Amazonas brasilero, Figueira et al.(2008) encontraron que durante los cuatro años siguientes a las actividades de extracción el bosque perdió biomasa. La mortalidad fue mayor a la predicha para este tipo de bosque, posiblemente por los efectos de daño mecánico, dado el aumento en la exposición al viento en los árboles adyacentes a los sitios de tala (Figueira et al., 2008). Sin embargo, estudios a largo plazo como el de Gourlet-Fleury et al. (2013) han encontrado que la tala selectiva favorece el aumento en la biomasa, el crecimiento y la supervivencia de árboles en todas las clases de diámetro inferior a 70 cm, así como el reclutamiento de especies de árboles de crecimiento rápido (pioneros), lo que aumenta las tasas de ganancia de biomasa aérea en el bosque. Otro efecto importante de la tala selectiva sobre los bosques es la formación de claros, que permiten la entrada de una mayor cantidad de luz al sotobosque, lo cual beneficia a las lianas y especies pioneras, pues son plantas que en su mayoría crecen rápidamente cuando hay una alta disponibilidad de luz (Philipson et al., 2014). La dinámica subsecuente es variable, los árboles pioneros pueden alcanzar la altura del dosel y establecerse o morir por falta de luz cuando el dosel se forme nuevamente sobre ellos (Philipson et al., 2014). Las lianas, si logran crecer hasta el dosel y establecerse sobre árboles de bosque maduro, pueden llegar a derribarlos con su peso, fomentando la formación de nuevos claros (Schnitzer & Bongers, 2002). De este modo, Schnitzer & Bongers (2002) demuestran que hay una relación negativa en la densidad de lianas y la densidad de árboles de bosque maduro, mientras que la densidad de lianas se relaciona positivamente con la densidad de especies pioneras.
En países neotropicales como Colombia, que presenta diversidad de clima y relieves, se vuelve aún más difícil predecir el rumbo que tomará la recuperación de los bosques objeto de tala selectiva. A pesar de tener un 52.6% (60 millones de hectáreas) del territorio nacional ocupado por bosques (IDEAM, 2014), Colombia presenta altas tasa de deforestación, con pérdidas de 5.4 millones de hectáreas de bosque en los últimos 20 años (García-Romero, 2013). La cuenca del Magdalena es la más deforestada de Sudamérica y la décima del mundo (García-Romero, 2013), pero presenta regiones como la serranía de las Quinchas, en el Magdalena medio, donde se conservan algunas áreas de bosque continuo. Por ejemplo, la reserva de aves El Paujil incluye 3419 hectáreas protegidas bajo la figura de Reserva privada de la sociedad civil (ProAves, s.f.) y presenta al menos cuatro tipos de bosque identificados por Aldana et al. (2008): bosque con un nivel moderado de la tala selectiva (bosques talados), bosques en llanuras aluviales, bosque secundario joven (bosque secundario) y bosque con tala de subsistencia (bosque primario).
El objetivo de este estudio fue analizar el efecto de la tala selectiva en la dinámica de la comunidad de árboles en el bosque de la reserva El Paujil en el Magdalena medio colombiano. Esta dinámica evaluada en términos de capacidad de acumular biomasa, dinámica de claros, regeneración, demografía y composición florística. Dados los efectos de aumento en disponibilidad de luz y exposición al viento después de la tala se espera que los bosques talados presenten una mayor tasa de reclutamiento y mortalidad, una tasa de cambio poblacional negativa y menor capacidad de acumular biomasa. Así mismo, por efectos directos de la tala se espera encontrar una menor densidad de individuos de tallas grandes y una menor cobertura del dosel, lo que permitirá que haya además una mayor densidad de plántulas y juveniles en el bosque talado. Adicionalmente, dado que la tala selectiva no ha sido considerada de gran impacto en la diversidad de los bosques, se espera encontrar similitudes en composición y recambio de especies vegetales entre los dos tipos de bosque, pero mayor abundancia de plantas pioneras en el bosque talado.
MATERIALES Y MÉTODOS
Área de estudio
Este estudio se realizó en la reserva de aves El Paujil, en los departamentos de Santander y Boyacá (74° 11´ W, 5° 56' N) con altitud geográfica desde los 150 m hasta los 1200 m y temperatura promedio anual de 27.8°C (Aldana et al., 2008). En el año se presentan dos periodos de lluvias, entre abril y mayo el primero y entre septiembre y noviembre el último. La humedad relativa oscila entre 85% y 89% (Balcázar-Vargas et al., 2000 citado en Aldana et al., 2008).
La reserva fue creada en noviembre de 2003 para conservar El Paujil de pico azul (Crax alberti) (ProAves, s.f.) y su hábitat, el bosque húmedo tropical. Antes del establecimiento de la reserva, la reforestadora Bosques del Futuro practicó tala selectiva para obtener madera durante cinco años en una parte del bosque. Silva-Herrera (1999) reportó que la reforestadora planeaba explotar 50 m³ de madera en pie en el bosque por hectárea al año, distribuidos así: 15 m3 de maderas finas y 35 m3 de maderas ordinarias. De acuerdo con esto, la reforestadora taló aproximadamente cinco árboles por hectárea, lo cual pudo ser verificado por los investigadores al momento de establecer las parcelas en 2006. Este bosque está ubicado en el departamento de Boyacá, en la vereda Puerto Pinzón del municipio de Puerto Boyacá. El bosque primario, fue explotado por los dueños del predio para la subsistencia, que corresponde a menos de un árbol por ha por año, como pudo ser evidenciado por los investigadores durante el establecimiento de las parcelas. Este bosque se encuentra en el departamento de Santander, en el municipio de Bolívar. La distancia entre los dos tipos de bosques (los lugares de muestreo) es de aproximadamente 8 km. Los bosques muestreados presentan una topografía con pendientes de hasta 40 °, y una altitud geográfica desde los 194 msnm hasta los 471 msnm.
Toma de datos
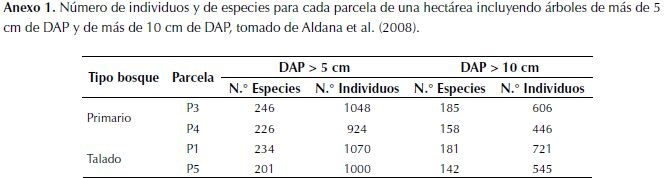
Para cada tipo de bosque se muestrearon dos parcelas de una hectárea, establecidas en el año 2006 por Aldana et al. (2008). Ellos realizaron mediciones de diámetro a la altura del pecho (DAP) para todos los individuos con DAP mayor a 5 cm y los identificaron hasta especie (o morfoespecie de no ser posible). La decisión de incluir individuos a partir de los 5 cm de DAP y no desde los 10 cm como generalmente suele hacerse, obedece a la presencia de una gran cantidad de especies de árboles que no alcanzan esta talla (anexo 1), y por ende se quedarían por fuera del muestreo, lo que llevaría a una subestimación en la diversidad y biomasa acumulada del lugar (Baraloto, 2013). En 2013 se censaron nuevamente, midiendo el DAP para los individuos muestreados en 2006, reportando los individuos muertos o desaparecidos y tomando nota de las causas de mortalidad en los casos que fuera posible, siguiendo el protocolo de Phillips et al. (2009). Se incluyeron como nuevos reclutas todos los individuos que entraron a la categoría de tamaño mayor a 5 cm de DAP. Se colectaron muestras de madera para determinar su densidad, usando un barrenador y tomando muestras de mínimo cinco individuos de las especies más abundantes de los dos tipos de bosque.
Para los estimativos de regeneración se comparó la cantidad de plántulas y juveniles presentes en los dos tipos de bosque tomando datos de 100 parcelas de 2x2 m para plántulas y 100 de 5x5 m para juveniles (Stevenson, 2011). Este muestreo se realizó de manera sistemática, dentro de las parcelas de 1 ha. Para cuantificar la entrada de luz al sotobosque se trabajó con 50 subparcelas de 20x20 m por tipo de bosque, ubicadas dentro de las parcelas de 1 ha. Se tomaron dos fotos del dosel en el punto central de cada una de las subparcelas con la cámara paralela al suelo a 1 m de altura. Se utilizó un lente ojo de pez y se programó la cámara para tomar las fotos a blanco y negro, con 22 cm de apertura constante y velocidad variable.
Análisis de datos
Se comparó la información obtenida en el año 2006 con la de 2013 para establecer las tasas anuales de crecimiento, mortalidad y reclutamiento de la comunidad de árboles para cada parcela utilizando las formulas presentadas por Sherman et al. (2012). Luego, para poder comparar estadísticamente estos comportamientos y que sean equiparables con la información obtenida de dinámica de claros y regeneración, se realizaron comparaciones a una escala de 20x20 m, con 50 subparcelas para cada tipo de bosque. Utilizando el programa estadístico R versión 3.0.1 (R Core Team, 2013)se hicieron pruebas t de una cola para dos muestras, con la aproximación de Welch cuando las varianzas no eran similares.
Para el análisis de claros se utilizaron las fotos tomadas en el punto central de cada una de las subparcelas de 20x20 m, escogiendo una por subparcela para determinar el valor de gris por pixel (donde 0 significa negro y 255 significa blanco) con el programa Image J (Image J, 2007). Este valor representa la intensidad lumínica en el sotobosque y es comparado con una prueba t de una cola para dos muestras entre los dos tipos de bosque utilizando el programa estadístico R versión 3.0.1 librería Vegan (R Core Team, 2013). Adicionalmente, como indicadores del proceso de regeneración de los bosques después del disturbio, se comparó la cantidad de plántulas y juveniles presentes en los dos tipos de bosque con pruebas t de una cola para dos muestras, con la aproximación de Welch cuando las varianzas no eran similares en el programa estadístico R versión 3.0.1 librería Vegan (R Core Team, 2013).
La densidad de las muestras de madera obtenidas en campo se determinó como la gravedad específica (peso seco / volumen verde) siguiendo el protocolo de densidad de madera de Chavé (2005). Para las especies de las que no se tomó muestra de madera se utilizaron los datos de densidad madera de Casas et al. (datos sin publicar) y los de Zanne et al. (2009). Posteriormente, se estimó la altura de cada árbol utilizando la ecuación alométrica derivada de la función de Weibull adaptada para Sur América presentada por Feldpausch et al. (2012), y se calculó la biomasa acumulada de cada árbol utilizando la ecuación I.3 de Alvarez et al. (2012)para bosque húmedo tropical en Colombia, que tiene en cuenta el diámetro a la altura del pecho, la altura del árbol y la densidad de la madera y, según Alvarez et al. (2012), es la mejor ecuación para estimar carbono en Colombia, dada la baja incertidumbre y variabilidad con respecto a las demás ecuaciones generadas por ellos. Con los valores de biomasa inicial y final se determinó el cambio de biomasa anual en Mg ha-1 año-1.
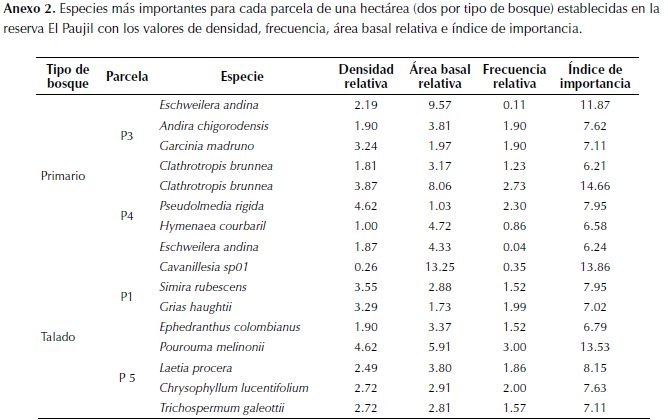
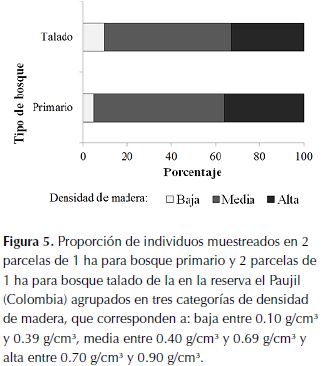
Finalmente, se identificaron las cuatro especies más importantes en cada parcela con el índice de valor de importancia que se determina con la densidad, frecuencia y área basal relativa de cada especie. Se buscó si estas especies son explotadas por su madera y la calidad de la misma (Fern, 2014) para relacionar algún posible cambio en las especies dominantes con la tala. Se buscó además el estado de conservación en Colombia de estas especies (Bernal et al., 2014)para indagar la vulnerabilidad en la que se encuentran los arboles maderables presentes en este bosque. Adicionalmente, se realizó una comparación de las especies presentes y su abundancia en las parcelas con un cluster representado en un dendrograma. Dado que las especies pioneras juegan un papel importante como indicadores de disturbios (tala o caída de árboles), se categorizaron los individuos muestreados en tres grupos dependiendo de la densidad de su madera, por ser uno de los rasgos funcionales más importantes en la determinación de especies de bosque primario y especies pioneras (Philipson et al., 2014). Se consideraron especies de árboles con densidad de madera baja aquellas con valores entre 0.10 g/cm³ y 0.39 g/cm³, media con madera entre 0.40 g/cm³ y 0.69 g/cm³ y alta las de madera entre 0.70 g/cm³ y 0.90 g/cm³. Posteriormente se comparó la proporción de árboles de cada categoría entre tipos de bosque con una prueba G en R versión 3.0.1 (R Core Team, 2013).
RESULTADOS
Demografía
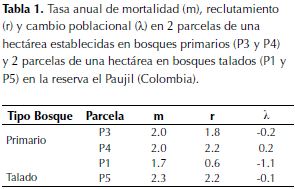
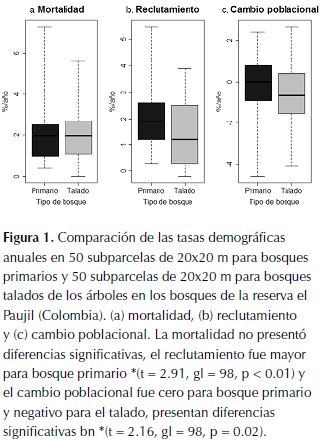
Los bosques talado y primario en la reserva El Paujil no presentaron diferencias significativas en la tasa anual de mortalidad (t = 0.13, gl = 98, p = 0.45; figura 1a), a pesar de la variación encontrada al hacer el análisis por subparcelas de 20x20 m, se puede ver que al comparar entre parcelas de 1 ha, el promedio es igual para los dos tipos de bosque, con una tasa anual de mortalidad promedio igual a 2 %. El reclutamiento fue mayor para el bosque primario (t = 2.91, gl = 98, p < 0.01; figura 1b), y, al igual que en el caso de la mortalidad a escala de 1 ha se corrobora el resultado (Tabla 1). El cambio poblacional fue cero para bosque primario y negativo para el talado, a escala de 20 x 20 m se encuentran diferencias significativas entre tipos de bosque (t = 2.16, gl = 98, p = 0.02; figura 1c).
Análisis de claros y regeneración.
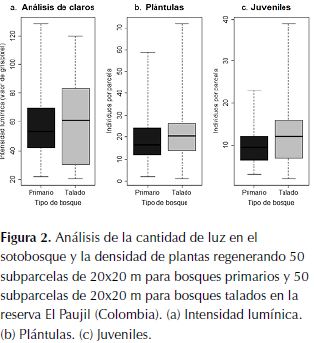
La cantidad de luz que llega al sotobosque en los bosques de la reserva El Paujil es similar en ambos tipos de bosque (t = 0.32, gl = 98, p = 0.63; figura 2a). Así mismo, tanto la cantidad de plántulas como la de juveniles no presentan diferencias significativas entre tipos de bosque (t de welch = -2.18, gl = 197, p = 0.98; t de welch = -4.09, gl = 155, p = 1 respectivamente; figura 2b y 2c).
Cambio de biomasa
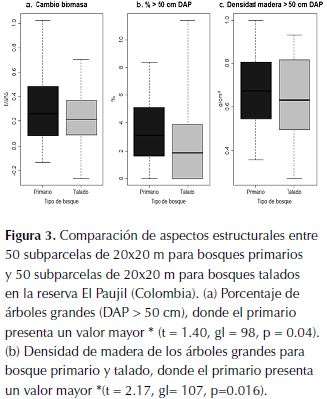
El bosque primario ganó en promedio 4 Mg de biomasa ha-1 año-1, pasando de tener en promedio 459.5 Mg ha-1 en el 2006 a tener 487.6 Mg ha-1 en el 2013, mientras que el talado ganó menos biomasa, con un promedio de 0.9 Mg ha-1 año-1, pasando de tener 440.6 Mg ha-1 en el 2006 a tener 447.1 Mg ha-1 en el 2013. Encontramos que hay menos árboles con un DAP mayor a 50 cm en el bosque talado (t = 1.40, gl = 98, p = 0.04; figura 3a), y, que la densidad de madera de estos árboles (<50 cm DAP) es menor que en el bosque primario (t = 2.17, gl = 107, p = 0.016; figura 3b).
Composición
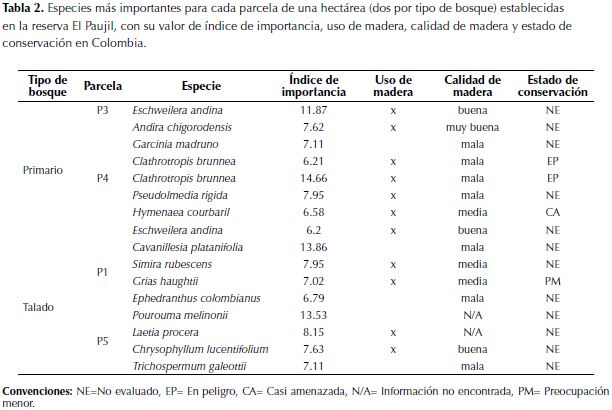
Las especies más importantes de las parcelas de bosque primario difieren de las del bosque talado, además, son en su mayoría árboles con madera de buena calidad (Fern, 2014), objetivo de las reforestadoras (tabla 2). El componente que más influyó en la determinación del índice de importancia fue el área basal relativa (anexo 2). El estado de conservación de estas especies en su mayoría no ha sido evaluado, no obstante se conoce que Clathrotropis brunnea está en peligro, Hymenaea courbaril se encuentra en preocupación menor y Grias haughtii es vulnerable (tabla 2).
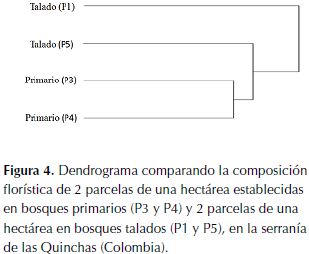
Se encontró una alta afinidad florística en las parcelas del bosque primario, mientras que el bosque talado es más heterogéneo, con una parcela más similar al grupo de bosque primario que la de su mismo tipo de bosque (figura 4).
Como resultado de la clasificación por densidad de madera, se encontró que la proporción de árboles en cada categoría de densidad de madera depende del tipo de bosque (g = 44.266, gl = 2, p < 0.01), encontrando una mayor proporción de especies con densidad de madera baja en el bosque talado (figura 5).
DISCUSIÓN
Demografía
Los componentes demográficos analizados para los bosques de la reserva El Paujil no se comportan como era esperado. La mortalidad fue igual para los dos tipos de bosque (tabla 1 y figura 1a), a pesar que se esperaba mayor en el bosque talado por daño mecánico a causa de una mayor exposición al viento de árboles contiguos a zonas de tala (Figueira et al., 2008). El reclutamiento, que se esperaba mayor en bosques talados por la mayor cantidad de luz disponible en consecuencia de la tala, resultó mayor en bosque primario (tabla 1 y figura 1b) y de hecho no hubo diferencias en la cantidad de luz que entra al sotobosque en los dos tipos de bosque (figura 2a). La población del bosque primario no presentó cambios, mientras que la del bosque talado disminuyó (tabla 1 y figura 1c), presentando diferencias significativas entre los dos tipos de bosque.
Como se mencionó anteriormente, los sitios de bosque muestreados tienen una altitud geográfica que oscila desde 194 m hasta 471 m, en zonas con pendientes de hasta 40o. Se ha demostrado que la pendiente del suelo es un factor ecológico primario, que controla la tasa de mortalidad por caída de árboles y, por tanto, induce un fuerte gradiente de luz en el sotobosque que favorece el crecimiento y reclutamiento de especies pioneras (Ferry et al., 2010). En el bosque de la reserva El Paujil, donde la alta mortalidad es casi igual en áreas con tala selectiva y de bosque primario, se puede decir que el efecto de la inclinación del terreno sobre la mortalidad de los árboles puede llegar a enmascarar el efecto de la tala como lo sugiere Ferry (2010).
Por otro lado, una alta abundancia de lianas en el bosque primario contribuiría a la formación de claros, pues afecta negativamente a los árboles de bosque maduro (Schnitzer, 2002). Para evaluar esta explicación se determinó y comparó la abundancia relativa de lianas en ambos tipos de bosque, lo cual corroboró lo predicho, pues resultó mayor en el bosque primario (t de Welch = 2.09, gl = 85, p = 0.02). Sin embargo, en comparación con otras parcelas de la misma extensión establecidas por investigadores del Laboratorio de ecología de bosques tropicales y primatología (LEBTYP) en Colombia, se pudo ver que el porcentaje de lianas en el bosque primario de la reserva El Paujil tiene un valor alto (2.98% en promedio), pero no significativamente mayor en comparación con las demás parcelas de tierra firme (1.46% en promedio N=20) (t de Welch = 2.92, gl = 2 p = 0.93). Por lo tanto, aún hay incertidumbre sobre el efecto de la abundancia de lianas sobre la dinámica de estos bosques.
Análisis de claros y regeneración
La magnitud de los claros en los bosques de la reserva El Paujil, similar en ambos tipos de bosque (figura 3) al igual que los anteriores resultados, lleva a pensar en una dinámica moldeada principalmente por el efecto de la pendiente del terreno, causando una alta mortalidad por caída de árboles semejante o en mayor magnitud que la ocasionada por el efecto del disturbio (viento) tras actividades de tala. Los vientos se han determinado como un factor clave en la mortalidad y las dinámicas post disturbio principalmente en zonas relativamente planas, como por ejemplo en el Amazonas, donde la influencia del viento puede ser mayor (Etter & Botero, 1990; Laurance & Curran, 2008).
A pesar de existir una proporción de claros y una cantidad de plántulas y juveniles similar en los dos tipos de bosque, el mayor reclutamiento observado en el bosque primario podría ser resultado del efecto del daño mecánico causado a plántulas y juveniles en el bosque talado, por la circulación constante de personas (por ejemplo: turistas e investigadores), que por su fácil acceso tiene una afluencia mucho mayor a la del bosque primario.
Cambio de biomasa
Los estimativos de reservas de biomasa por hectárea para los dos tipos de bosque son relativamente altos cuando se comparan con estimaciones realizada para estos bosques en Colombia (Phillips et al., 2011), sin embargo, esto se explica con el hecho de que en este estudio se incluyeron árboles desde 5 cm de DAP. Esta no es una práctica común en estudios de estimación de biomasa, sin embargo, es importante incluir los individuos de menor talla, dado que no solo aumentan los cálculos de reservas biomasa por ha (Baraloto et al., 2013), por efecto del aumento del número de individuos censados, que puede ser de casi el doble, sino también los cálculos de diversidad de especies (anexo 1).
Como era de esperarse el bosque primario tuvo una ganancia neta de biomasa por hectárea mayor que el bosque talado. Esto concuerda con lo que reportado en estudios de dinámica de biomasa en bosques primarios tropicales, donde se han reportado incrementos anuales de biomasa por hectárea del orden de 3 a 20 toneladas (Meister et al., 2012). Sin embargo, es notable que los menores valores, reportados en el presente estudio para el bosque talado, se asemejan a los valores reportados por otros estudios de dinámica de biomasa en bosques fragmentados (Nascimento & Laurance, 2004). Adicionalmente, la mayor cantidad de árboles grandes observada en el bosque primario (figura 2b), sumada que estos tienen árboles con maderas de mayor densidad (figura 2c), pone en evidencia un efecto de la tala selectiva que no puede ser comparado tan fácilmente con factores intrínsecos de la dinámica del bosque y que tarda largos periodos de tiempo en volver a la normalidad. Es clara la disminución en las poblaciones de ciertas especies de árboles, que son objeto de explotación forestal por la calidad de su madera. A la luz de estos resultados, es importante resaltar que, de continuar la fragmentación y la tala selectiva en esta región, los efectos sobre las dinámicas del carbono podrán ser extremadamente negativos hasta el punto que estos bosques pueden dejar de ser reservorios de carbono para convertirse en fuentes de emisiones de CO2, como se ha previsto para bosques fragmentados de la amazonia (Laurance et al., 2011).
Composición
Asumiendo que los bosques tenían una composición similar antes de la tala, las diferencias al comparar la composición y dominancia entre tipos de bosque (tabla 2 y figura 4) se pueden relacionar con esta actividad. Ya que, por ejemplo, las especies de árboles de grandes tallas (mayor área basal) y madera con densidad alta (considerado un factor importante de calidad) son menos importantes en el bosque talado (tabla 2, figura 5). Nuevamente, se pone en evidencia que el efecto más fuerte de la tala selectiva sobre el bosque fue generar cambios en la composición de especies, al disminuir las poblaciones de especies de árboles maderables. Este factor debe ser considerado en planes de manejo de las industrias madereras, pues algunas de estas especies se encuentran en estado de vulnerabilidad (tabla 2) o se desconoce su estatus de conservación actual (Bernal et al., 2014). Adicionalmente, puede tener implicaciones en las interacciones tróficas del ecosistema, donde habitan dispersores de gran importancia como el mono araña (Ateles hybridus) críticamente amenazados y con poblaciones decrecientes (Aldana et al., 2008).
Otros estudios han reportado cambios en la composición de especies de bosques de la región, como una respuesta a los cambios en el clima (Duque et al., 2015), que sumados a los efectos de la tala selectiva que evidenciamos en este estudio, podrían causar fuertes disminuciones en las poblaciones de especies vegetales de estos bosques.
CONCLUSIONES
La dinámica del bosque de la reserva El Paujil no muestra una gran diferenciación a causa de la tala selectiva practicada en el pasado. Es importante resaltar el efecto de la topografía del terreno sobre la dinámica del bosque y se recomiendan estudios sobre el efecto de las lianas y el tránsito de personas. La estructura del bosque cambia y las reservas de biomasa disminuyen a causa de la tala selectiva, pues hay una importante disminución en las poblaciones de árboles grandes de especies con madera de buena calidad en el bosque talado, lo que puede repercutir en las interacciones tróficas del bosque y en su capacidad de ser sumideros de carbono. Este tipo de especies deben ser manejadas para evitar la disminución irreversible de sus poblaciones.
AGRADECIMIENTOS
Por los fondos proporcionados para la toma de datos agradecemos al Fondo de apoyo doctoral de la Facultad de Ciencias de la Universidad de los Andes. A la fundación ProAves, que gracias los convenios de investigación en la Reserva Natural de las Aves El Paujil permitió el establecimiento de las parcelas y posterior remuestreo. Al equipo de trabajo de Aldana por el establecimiento de las parcelas y a Guillermo Rivas, Angela Perilla, Pablo Negret, Sebastian Gonzales, Alejandra Jimenez, Diana Pizano y Sasha Cárdenas por la ayuda en el remuestreo de las parcelas.
REFERENCIAS BIBLIOGRÁFICAS
Aldana, A. M., Beltrán, M., Torres-Neira, J., & Stevenson, P. R. (2008). Habitat characterization and population density of brown spider monkeys (Ateles hybridus) in Magdalena Valley, Colombia. Neotropical Primates, 15(agosto), 46-50. [ Links ]
Álvarez, E., Duque, A., Saldarriaga, J., Cabrera, K., de las Salas, G., del Valle, I., Lema, A., Moreno, F., Orrego, S., Rodríguez, L. (2012). Tree above-ground biomass allometries for carbon stocks estimation in the natural forests of Colombia. Forest Ecology and Management, 267, 297-308. [ Links ]
Asner, G. P., Keller, M., & Silva, J. N. M. (2004). Spatial and temporal dynamics of forest canopy gaps following selective logging in the eastern Amazon. Global Change Biology, 10(5), 765-783. [ Links ]
Balcazar, M. P., Rangel, J. O., & Linares, E. L. (2000). Diversidad floristica de la serrania de Las Quinchas, Magdalena medio (Colombia). Caldasia, 22(2), 191-224. [ Links ]
Baraloto, C., Molto, Q., Rabaud, S., Hérault, B., Valencia, R., Blanc, L., Fine, P. V. A. & Thompson, J (2013). Rapid simultaneous estimation of aboveground biomass and tree diversity across Neotropical forests: a comparison of field inventory methods. Biotropica, 45(3), 288-298. [ Links ]
Bernal, R., Robbert Grandstein, S., & Celis, M. (2014). Catálogo de plantas y líquenes de Colombia. Recuperado de: http://catalogoplantasdecolombia.unal.edu.co/ [ Links ]
Chave, J. (2005). Measuring wood density for tropical forest trees. A field manual for the CTFS sites. Touluse, France. 1-7. [ Links ]
Duque, A., Stevenson, P. R., & Feeley, K. J. (2015). Thermophilization of adult and juvenile tree communities in the northern tropical Andes. Proceedings of the National Academy of Sciences of the United States of America, 112(34), 10744-9. [ Links ]
Etter, A., & Botero, P. J. (1990). Efectos de los procesos climáticos y geomorfológicos en la dinámica del Bosque Húmedo Tropical de la Amazonía Colombiana. Colombia Amazonica, 4(2), 7-21. [ Links ]
Feldpausch, T. R., Lloyd, J., Lewis, S. L., Brienen, R. J., Gloor, M., Monteagudo Mendoza, A., Lopez-Gonzalez, G., Banin, L., Abu Salim, K., Affum-Baffoe, K., Alexiades, M., Almeida, S., Amaral, I., Andrade, A., Aragão, L. E. O. C., Araujo Murakami, A., Arets, E. J. M. M., Arroyo, L., Aymard C. G. A., Baker, T. R., B'anki, O. S., Berry, N. J., Cardozo, N., Chave, J., Comiskey, J. A., Alvarez, E., de Oliveira, A., Di Fiore, A., Djagbletey, G., Domingues, T. F., Erwin, T. L.,Fearnside, P. M., Franca, M. B., Freitas, M. A., Higuchi, N., Honorio C., E., Iida, Y., Jiménez, E., Kassim, A. R.,Killeen, T. J., Laurance, W. F.,Lovett, J. C.,Malhi, Y., Marimon, B. S., Marimon-Junior, B. H., Lenza, E.,Marshall, A. R., Mendoza, C., Metcalfe, D. J., Mitchard, E. T. A., Neill, D. A., Nelson, B.W., Nilus, R.,Nogueira, E. M., Parada, A., Peh, K. S.-H., Pena Cruz, A., Peñuela, M. C., Pitman, N. C. A., Prieto, A., Quesada, C. A., Ramirez, F., Ramirez-Angulo, H.,Reitsma, J. M., Rudas, A., Saiz, G., Salomao, R. P., Schwarz, M., Silva, N., Silva-Espejo, J. E., Silveira, M. Sonké, B., Stropp, J., Taedoumg, H. E., Tan, S., Ter Steege, H., Terborgh, J., Torello-Raventos, M., van der Heijden, G. M. F., Vasquez, R., Vilanova, E., Vos, V. A., White, L., Willcock, S.,Woell, H. & Phillips, O. L. (2012). Tree height integrated into pan-tropical forest biomass estimates. Biogeosciences Discussions, 9(3), 2567-2622. [ Links ]
Fern, K. (2014). Useful Tropical Plant Database. Recuperado de: http://tropical.theferns.info/. [ Links ]
Ferry, B., Morneau, F., Bontemps, J.-D., Blanc, L., & Freycon, V. (2010). Higher treefall rates on slopes and waterlogged soils result in lower stand biomass and productivity in a tropical rain forest. Journal of Ecology, 98(1), 106–116. [ Links ]
Figueira, A. M. E. S., Miller, S. D., de Sousa, C. A. D., Menton, M. C., Maia, A. R., da Rocha, H. R., & Goulden, M. L. (2008). Effects of selective logging on tropical forest tree growth. Journal of Geophysical Research, 113(G00B05), 1–11. [ Links ]
Garcia Romero, H. G. (2013). Deforestación en Colombia :Retos y perspectivas. En: F. Dane (Ed.), El Desafio del Desarrollo Sustentable en América Latina, 123-142. Rio de Janerio: Konrad Adenauer Stiftung. [ Links ]
Gibson, L., Lee, T. M., Koh, L. P., Brook, B. W., Gardner, T. A, Barlow, J., Peres, C. A., Bradshaw, C. J. A., Laurance, W. F., Lovejoy, T. E., Sodhi, N. S. (2011). Primary forests are irreplaceable for sustaining tropical biodiversity. Nature, 478(7369), 378-81. [ Links ]
Gourlet-Fleury, S., Mortier, F., Fayolle, A., Baya, F., D., Quédraogo, F., & Picard, N. (2013). Tropical forest recovery from logging: a 24 year silvicultural experiment from Central Africa. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 368(1625), 2012-3020. [ Links ]
IDEAM (Instituto de Hidrología, Meteorología y Estudios Ambientales), Subdirección de Ecosistemas e Información Ambiental, Grupo de Bosques. (2014). Proyecto Sistema de Monitoreo de Bosques y Carbono. Bogota, D. C., Colombia. [ Links ]
ImageJ. Rasband, W. (2007). ImageJ: Image Processing and Analysis in Java. Bethesda, Maryland: US National Institutes of Health. [ Links ]
Laurance, W. F., Camargo, J. L. C., Luizão, R. C. C., Laurance, S. G., Pimm,S. L., Bruna, E. M., Stouffer, P. C., Bruce Williamson, G., Benitez-Malvido, J., Vasconcelos, H. L. (2011). The fate of Amazonian forest fragments: A 32-year investigation. Biological Conservation, 144(1), 56-67. [ Links ]
Laurance, W. F., & Curran, T. J. (2008). Impacts of wind disturbance on fragmented tropical forests: A review and synthesis. Austral Ecology, 33(4), 399-408. [ Links ]
Meister, K., Ashton, M. S., Craven, D., & Griscom, H. (2012). Managing Forest Carbon in a Changing Climate, 51-75. [ Links ]
Nascimento, H. E. M., & Laurance, W. F. (2004). Biomass dynamics in amazonian forest. Fragments, 14(4), s127-s138. [ Links ]
Norden, N., Chazdon, R. L., Chao, A., Jiang, Y.-H., & Vilchez-Alvarado, B. (2009). Resilience of tropical rain forests: tree community reassembly in secondary forests. Ecology Letters, 12(5), 385-94. [ Links ]
Phillips J.F., Duque A.J., Yepes A.P., Cabrera K.R., Navarrete D.A., Álvarez E., Cardenas D. (2011). Estimación de las reservas potenciales de carbono almacenadas en la biomasa aérea en bosques naturales de Colombia. Informe Final. Instituto de Hidrologia, Meteorologia, y Estudios Ambientales-IDEAM-Bogota D.C., Colombia. 68 pp. [ Links ]
Phillips, O., Baker, T., Feldpausch, T., Brienen, R. (2009). Manual de campo para la remedición y establecimiento de parcelas. En: RAINFOR (Ed., 2nd ed.), Proyecto PAN-AMAZONIA. [ Links ]
Philipson, C. D., Dent, D. H., O'Brien, M. J., Chamagne, J., Dzulkifli, D., Nilus, R., Philips, S. Reynolds, G., Saner, P., Hector, A. (2014). A trait-based trade-off between growth and mortality: evidence from 15 tropical tree species using size-specific relative growth rates. Ecology and Evolution, 4(18), 3675-3688. [ Links ]
ProAves (s.f.). Reserva Natural de las Aves El Paujil. Recuperado de: http://www.proaves.org/rna-el-paujil/ [ Links ]
R Core Team. (2013). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. [ Links ]
Schnitzer, S. A., & Bongers, F. (2002). The ecology of lianas and their role in forests. Trends in Ecology and Evolution, 17(5), 223-230. [ Links ]
Sherman, R. E., Fahey, T. J., Martin, P. H., & Battles, J. J. (2012). Patterns of growth, recruitment, mortality and biomass across an altitudinal gradient in a neotropical montane forest, Dominican Republic. Journal of Tropical Ecology, 28(05), 483-495. [ Links ]
Silva Herrera, L. J. (1999). Plan de factibilidad y estrategico de la reforestadora Bosques del Futuro S.A. ubicada en el Magdalena medio. Bogota: Universidad de La Salle. 1-130. [ Links ]
Stevenson, P. R. (2011). The Abundance of Large Ateline Monkeys is Positively Associated with the Diversity of Plants Regenerating in Neotropical Forests. Biotropica, 43(4), 512–519. [ Links ]
Zanne, A. E., Lopez-Gonzalez, G., D.A., C., Ilic, J., Jansen, S., Lewis, S. L. S. L., Miller, R.B. B.,Swenson, N.G. G., Wiemann, M.C. C., Chave, J. (2009). Global Wood Density Database. Dryad Digital Repository. Recuperado de:http://hdl.handle.net/10255/dryad.235 [ Links ]
ANEXOS