










Servicios Personalizados
Revista
Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista de Medicina Veterinaria
versión impresa ISSN 0122-9354versión On-line ISSN 2389-8526
Rev. Med. Vet. n.20 Bogotá jul./dic. 2010
Valor genético estimado e QTL afetando
porcentagem de sólidos totais na raça bovina gir
Valor genético estimado y QTL que afecta el porcentaje
de sólidos totales en la raza bovina GYR
Ariosto Ardila Silva*
* Licenciado en Ciencias de la Educación, Zootecnista Especialista en Producción en Ganado de Leche, M.Sc. en Genética de Poblaciones, Ph.D. en Genética y Mejoramiento de la Universidad Federal de Viçosa. Correo electrónico: ariosto.silva@ufv.br
Fecha de recepción: enero 28 de 2010
Fecha de aprobación: septiembre 16 de 2010
Resumo
Além de todos os progressos que têm representado o melhoramento genético animal clássico, este apresenta algumas limitações, como em alguns casos onde o fenótipo não se expressa no indivíduo. Com o advento das enzimas de restrição, do seqüenciamento e amplificação do DNA, foi possível o uso dos marcadores moleculares associados a locos de características quantitativas (QTL), como ferramenta que auxilia e aprimora o melhoramento genético animal por meio da Seleção Assistida por Marcadores (MAS), aumentando o ganho genético para características de difícil mensuração, baixa herdabilidade e limitadas pelo sexo. A raça bovina gir é caracterizada por sua rusticidade e adaptação ao trópico, apresentando uma boa produção de leite para os países tropicais, e sendo fundamental na conformação da raça Girolando (5/8 Holandês e 3/8 gir). O propósito do presente estudo foi complementar a análise quantitativa com a ferramenta molecular, no melhoramento genético da característica porcentagem de sólidos totais na raça gir no Brasil. Quatorze famílias (touros) gir com 657 filhas foram analisadas, sendo a média de 0,095% e o desvio padrão 0,1923 do valor genético estimado para porcentagem de sólidos totais. Ao empregar 27 marcadores microssatélites na análise através das 14 familias para porcentagem de sólidos totais no cromossomo 6, um QTL foi identificado com P < 0,05 e F = 2,34 na posição 58 cM, próximo ao marcador MNB-208 (60,21 cM). Ao empregar as famílias 3 e 4, as mais significativas, o mesmo QTL foi identificado com P < 0,01 e F = 7,61, mas na posição 58 cM, próximo ao marcador MNB-208.
Palavras chave: bovinos de leite, controle leiteiro, delineamento de filhas, microssatélites.
Resumen
Además de todos los progresos que ha representado el mejoramiento genético animal clásico, este presenta algunas limitaciones como en algunos casos donde el fenotipo no se expresa en el individuo. Con el advenimiento de las enzimas de restricción, del secuenciamiento y de la amplificación del ADN, fue posible el uso de marcadores moleculares asociados con loci de características cuantitativas (QTL), como herramienta que auxilia y perfecciona el mejoramiento genético animal por medio de la selección asistida por marcadores (MAS), lo que aumenta el mérito genético para características de difícil medida, baja heredabilidad y limitadas por el sexo. La raza bovina gyr se caracteriza por su rusticidad y adaptación al trópico, y presenta una buena producción de leche para los países tropicales, por lo que es fundamental en la conformación de la raza gyrholando (5/8 holstein y 3/8 gyr). El propósito del presente estudio fue complementar el análisis cuantitativo con la genética molecular, en el mejoramiento genético de la característica porcentaje de sólidos totales en la raza gyr en Brasil. Catorce familias (toros) gyr con 657 hijas fueron analizadas, siendo el promedio 0,095 % y la desviación estándar 0,1923 del valor genético estimado para porcentaje de sólidos totales. Al utilizar 27 marcadores microsatélites en el análisis a través de las 14 familias para porcentaje de sólidos totales en el cromosoma 6, un QTL fue identificado con P < 0,05 y F = 2,34 en la posición 58 cm, próximo al marcador MNB-208 (60,21 cm). Al emplear las familias 3 y 4, las más significativas, el mismo QTL fue identificado con P < 0,01 y F = 7,61, pero en la posición 58 cm, próximo al marcador MNB-208.
Palabras clave: bovinos de leche, control lechero, diseño de hijas, microsatélites.
Introdução
O Brasil é o sexto país colocado na classificação mundial de produção de leite bovino, o que equivale a uma produção de mais de 25 bilhões de toneladas de leite por ano, porém, a produção de leite caracterizase pela baixa produtividade média, menos de 1.500 litros/vaca/ano. A vaca Holandesa apresenta maior capacidade de produção de leite no mundo, porém, a dificuldade de adaptação aos trópicos torna difícil e onerosa à produção de leite. Nesse contexto é que a raça Gir mostra a sua grande importância, tanto como raça pura, como nos cruzamentos, contribuindo para a formação do Girolando (5/8 Holandesa: 3/8 Gir). Calor e umidade são fatores reconhecidamente prejudiciais para a reprodução das raças taurinas, e seus efeitos são agravados pela alta produção do leite. A incapacidade do bovino europeu em eliminar o excesso de calor pela deficiência de seu aparelho termo-regulador torna difícil sua adaptação aos trópicos, diminuindo sua produção de leite.
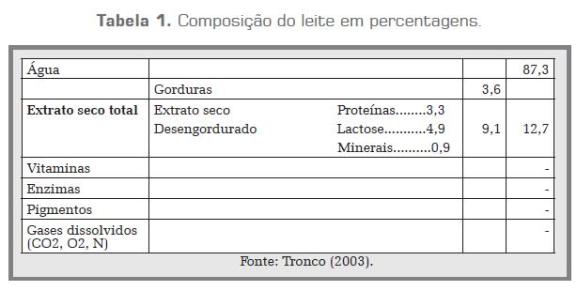
Segundo Madalena (2008), aproximadamente 12% do leite é formado por substâncias sólidas: proteína, gordura, lactose e minerais, sendo a água os 88% restantes. São os sólidos, especialmente as proteínas, que fazem do leite um produto importante, com alto valor nutritivo, e os que servem de matéria-prima para a elaboração dos diferentes produtos lácteos, como queijo, manteiga, iogurtes, sorvetes, etc. A composição média do leite de vaca está especificada no quadro 1.
Segundo Martínez e Verneque (2001), a avaliação de um touro por meio do desempenho de suas progênies constitui-se na maneira mais segura e eficiente de se prever com precisão a capacidade do animal transmitir sua superioridade (ou inferioridade) genética aos seus descendentes. De acordo com estes mesmos autores, os touros depois de avaliados são classificados de acordo com seu valor genético (VG) ou pela capacidade prevista de transmissão (PTA, Predited Transmitting Ability) que é igual à metade do VG. Do anterior, pode-se deduzir que um teste de progênie vem sendo um teste de comparação de touros, e que os melhores classificados devem ser utilizados para produção de progênies da próxima geração.
O BTA6 (Bos taurus autosome 6) é um dos cromossomos mais estudados, sendo que QTL significativos e genes candidatos, associados às características de produção de leite, têm sido mapeados, principalmente nas raças taurinas. Portanto, espera-se que no mesmo cromossomo da raça Gir Leiteiro sejam encontrados QTL associados às características de porcentagem de sólidos totais. O maior objetivo de mapear QTL é encontrar genes/marcadores que possam ser utilizados em programas de melhoramento, por meio da MAS e, especificamente, em gado de leite podem ser empregados para selecionar touros jovens, como futuros candidatos ao teste de progênie, assim, incrementar o diferencial de seleção, reduzir o intervalo de gerações e aumentar o ganho genético. O objetivo do trabalho foi calcular os valores genéticos estimados e mapear QTL no cromossomo 6 associado à característica de porcentagem de sólidos totais na raça bovina Gir leiteiro.
Material e métodos
Valor genético para porcentagem de sólidos totais
O valor fenotípico da característica porcentagem de sólidos totais foi obtido do Arquivo Zootécnico Nacional (AZN) do ano 2008. O modelo estatístico empregado (y = Xb + Za + Wpe + e) na avaliação genética dos animais considera como efeitos fixos, rebanho-ano de parto, estação de parição da vaca, composição genética da filha do touro e a idade da vaca ao parto e os efeitos aleatórios foram os erros e o efeito do animal (vaca, pai e mãe). Para avaliação genética das vacas foi incluído o efeito aleatório de meio permanente por possuir medidas repetidas no mesmo animal.
O modelo de repetibilidade usualmente é da forma:
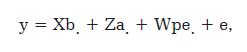
Em que y= vetor de observações; b= vetor de efeitos fixos; a= vetor de efeito animal, aleatório; pe= vetor de efeitos ambientais permanentes e efeitos genéticos não-aditivos; e= vetor de efeitos residuais aleatórios; X,Z,W, são matrizes de incidência de efeitos fixos, animal e de ambiente permanente, respectivamente. O vetor "a" unicamente inclui efeito animal aleatório aditivo, consequentemente, efeitos genéticos não-aditivos estão incluídos no termo pe. É assumido também que, efeito de ambiente permanente e efeito residual, estão independentemente distribuídos com medias zero e variâncias σ2pe e σ2e, respectivamente. O software MTDFREML (Multiple Trait Derivate Free Restricted Maximum Likelihood) ou Estimação pelo Método da Máxima Verossimilhança Restrita adotando-se o procedimento livre de derivadas (DF) foi utilizado para o cálculo dos componentes de variâncias e os valores genéticos no presente estudo.
Familias utilizadas para o mapeamento de QTL no rebanho GIR Leteiro
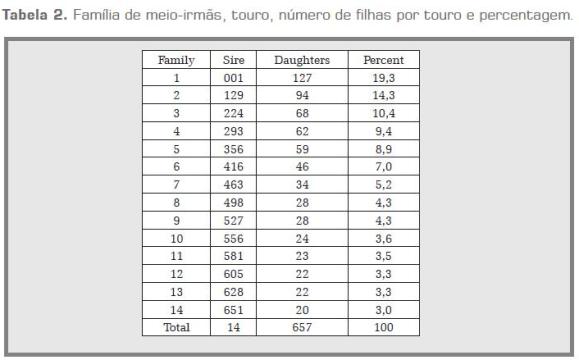
Para o desenvolvimento deste trabalho foram utilizadas 657 vacas Gir pertencentes ao controle leiteiro, filhas de 14 touros Gir, dos quais 12 foram positivos para o teste de progênie e os outros 2 foram pais de touros positivos para o mesmo teste. O critério utilizado para o número de filhas por touro baseou-se na existência de amostras de DNA para as vacas filhas por touro, sendo vinte o número mínimo de filhas para ser considerada uma família. Na tabela 2 são apresentados famílias, número de filhas por touro.
Genotipagem com marcadores microssatélites
Para a extração do DNA seguiu-se o protocolo modificado de Sambrook e Russel (2001). Para quantificação do DNA foi usado o equipamento Nanodrop e a qualidade foi conferida pela razão de absorbância 260/280 nm. Para a genotipagem do cromossomo 6 foram utilizados marcadores microssatélites, escolhidos do mapa consenso e disponibilizado pelo MARC/USDA (Meat Animal Research Center e United States Department of Agriculture, http://www.marc.usda.gov/genome/genome.htlm). Na escolha dos marcadores levaram-se em conta sua posição, número de alelos, e um mínimo de 50% de heterozigosidade. Foram selecionados 27 marcadores microssatélites ao longo do cromossomo 6, com distância média entre marcadores de 4,9 cM, que foram utilizados para genotipar 14 touros e 657 vacas, em um total de 671 animais.
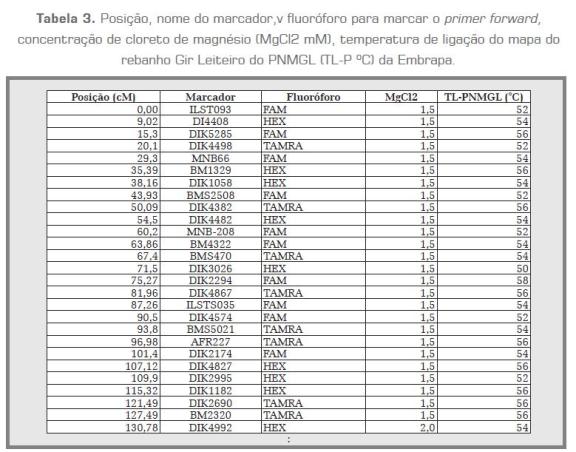
Para as reações de PCR, foram testadas três concentrações de MgCl2 (1,5 mM; 2,0 mM e 2,5 mM). Também foram testadas cinco temperaturas de anelamento (TA) para cada primer. Os produtos das amplificações foram genotipados por eletroforese capilar no equipamento MegaBACE 1000 (GE Healthcare, NYSE, Germany). Os genótipos foram analisados no programa Fragment Profiler e os dados exportados para uma planilha no Excel. Foram definidas as condições ótimas de TA e MgCl2 para cada marcador (tabela 3).
Delineamento para detecção de QTL
No presente estudo foi utilizado o delineamento de meio-irmãs ou delineamento de filhas, proposto inicialmente por Neimann-Soressen e Robertson (1961). As análises preliminares de QTL para todos os marcadores foram realizadas pelo seguinte modelo:
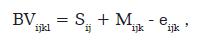
Em que: BVijkl é o EBV (Estimated Breeding Values) para a característica i da vaca l, que recebeu o alelo paterno k; Sij. é o efeito do touro j sobre a característica i; Mijk é o efeito do alelo paterno k do touro j sobre a característica i, e e é o resíduo aleatório associado com cada registro. Um efeito de alelo paterno significante é indicativo de um QTL ligado ao marcador molecular segregando. As análises para identificação de QTL no cromossomo 6 foram realizadas em mapeamento por intervalo, com um delineamento de meioirmãs, utilizando o software GridQTL (http://www.gridqtl.org.uk) (Seaton et ál., 2006). Os limiares de significância a nível cromossômico (1% e 5%) foram calculados no programa GridQTL a partir de testes de permutação (10.000) (Churchill e Doerge, 1994).
Resultados e discussão
Mapa de ligação
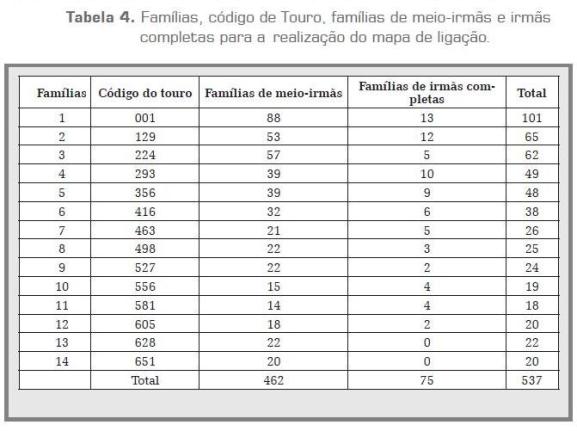
As distâncias de mapa entre os 27 marcadores analisados foram computadas com a opção "fixed" do Crimap (Green et ál., 1990). Para a organização dos dados, uma família de meio-irmãs foi conformada pela filha, pai e a mãe. Na maioria dos touros, foram encontradas filhas partilhando à mesma mãe, estas foram consideradas como famílias de irmãs completas. Na tabela 4 encontram-se as famílias de meioirmãs e irmãs completas para cada touro e o total de famílias por touro.
Segundo Lynch e Walsh (1998) um mapa de ligação está em função do número de marcadores, dos animais utilizados e do tipo de delineamento em estudo. Para gerar o mapa do cromossomo 6 dentro do lineamento de filhas, foi montada uma planilha em que os dados foram organizados pelas famílias com as informações de pais e filhas (tabela 5). Esta planilha continha 537 famílias para o mapa gerado da raça Gir Leiteiro do Programa Nacional de Melhoramento do Gir Leiteiro (PNMGL) e 19 marcadores microssatélites, sendo retirados os marcadores com maior distorção.
Na acurácia do mapa de ligação, o número de indivíduos está relacionado, uma vez que, quanto maior o número de meioses informativas, maior a probabilidade de detectar recombinação entre pequenas distâncias, característica em populações comerciais. O número médio de meioses informativas encontradas dentro da população Gir Leiteiro (167,9) foi menor que a média do mapa MARC (467,3). Isto pode ser explicado pelo número de indivíduos e o delineamento utilizado. O marcador ILSTS0936 foi o único que apresentou maior número de meioses informativas (164) para o mapa da população Gir. A ordem dos marcadores não foi alterada após a formação dos novos mapas. Para todas as análises de mapeamento foi utilizado o mapa consenso, porque tem sido o mapa comumente aceitado e porque mostrou que as diferenças em freqüências de recombinação estimadas não apresentaram viés no teste para QTL ou efeitos de QTL estimados (Haley e Knott, 1992).
Mapeamento de QTL
Na tabela 6 encontram-se os touros e as filhas genotipadas por marcador, e o número de touros heterozigotos por marcador que foram utilizados para as análises de mapeamento de QTL. O loco que apresentou maior número de filhas genotipadas foi o DIK4574 com 614 filhas, correspondente a 93%. Já os marcadores com menor número de filhas genotipadas foram DIK4382 e DIK3026 com 527 (80%) e 532 (80%), cada um para as respectivas análises. Todos os 14 touros foram heterozigotos para o marcador BMS2508, que sua alta heterozigosidade esperada o converte em um forte candidato para testes de identidade e paternidade nas raças zebuínas por seu alto poder de discriminação individual. A metade dos touros foi heterozigota para os marcadores DIK4867 e DIK4992, sendo os marcadores que apresentaram o menor número de touros heterozigotos. O touro 651 apresentou o maior número de marcadores heterozigotos, 25 em total, enquanto que o touro 628 foi heterozigoto para 16 marcadores microssatélites.
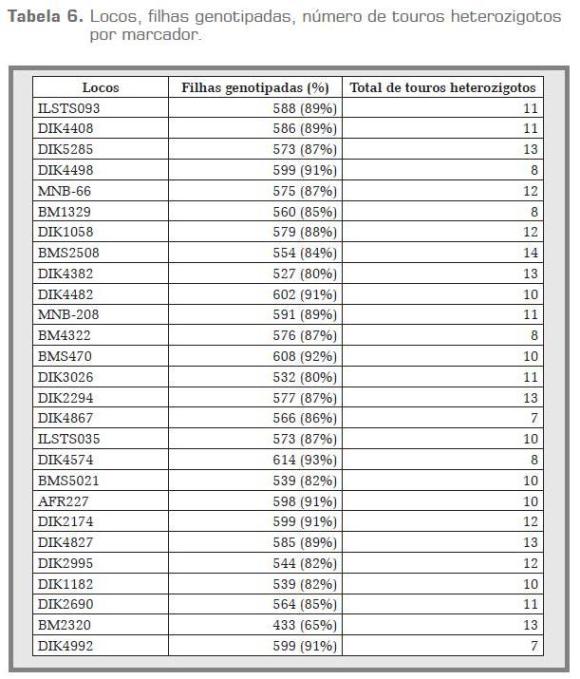
De acordo com as leis de Mendel, cada pai e progênie devem apresentar ao menos um alelo comum. Discrepâncias entre genótipos de uma vaca e seu suposto pai (touro) podem ser devido a mutações, errosna genotipagem ou erros nos registros de paternida de. As fêmeas que não apresentaram pelo menos um alelo do touro para mais de cinco locos (> 5) foram descartadas para a realização das diferentes análises. Um total de 35 vacas (5,32%) não foi considerado filhas por incorreta paternidade, estando de acordo com Ron et ál. (1996) e Ron et ál. (2001), que aplicando delineamento de filhas consideraram uma paternidade mínima de 90%. Para as famílias 498 e 628, todas as fêmeas foram filhas, portanto, assumidas nas análises. A família 651 apresentou o maior número de filhas descartadas, 4 de 20, o que é equivalente a 20% (tabela 6).
O conteúdo de informação (CI) para a porcentagem de sólidos totais está representado na figura 1. O CI foi calculado da variância das probabilidades condicionais de herdar uma região cromossômica a cada cM como uma proporção da variância quando o descendente verdadeiro é conhecido. O CI através de todas as famílias ao longo do cromossomo 6 foi maior que nas famílias 3 e 9 juntas, exceto na região compreendida entre os marcadores DIK4482 e MNB-208.
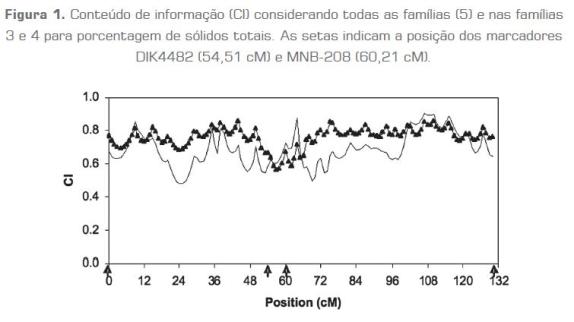
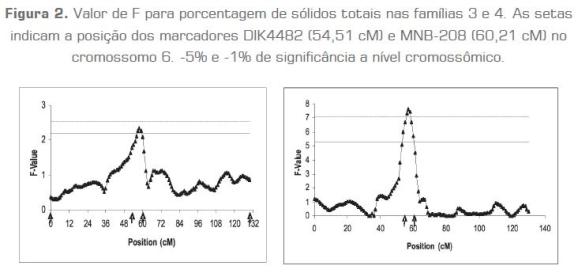
Após a análise, considerando todas as famílias para a característica porcentagem de sólidos totais, foram identificadas as famílias mais significativas de acordo com os valores absolutos de "t", que são as significâncias dos efeitos dentro de cada família do alelo marcador paterno. As famílias que apresentaram um nível de significância igual ou maior a 5% para um QTL específico foram consideradas significativas e levadas em conta para a detecção de QTL ao longo do cromossomo 6. Considerando todas as famílias para porcentagem de sólidos totais, um QTL foi identificado com P< 0,05 e F = 2,34 na posição 58 cM, próximo ao marcador MNB-208 (60,21 cM). Ao empregar as famílias 3 e 4, as mais significativas, o mesmo QTL foi identificado com P < 0,01 e F =7,61, mas na posição 58 cM, próximo ao marcador MNB-208 (figura 2).
Para produção e composição do leite as pesquisas foram feitas, inicialmente para genes candidatos, especialmente genes para proteínas lácteas, as quais apresentam efeitos apenas modestos sobre produtividade. O primeiro experimento utilizando marcadores distribuídos por todo o genoma, com um grande número de touros e utilizando delineamento de filhas (Grand-Daughter Design) na raça Holandesa foi conduzido nos Estados Unidos por Georges et ál. (1995) que identificaram evidências de QTL para características produtivas nos cromossomos 1, 6, 9, 10 e 20.
Khatkar et ál. (2004) utilizando a metodologia de Meta-análise encontraram QTL praticamente em todos os 29 autossomos do genoma bovino leiteiro, mas os cromossomos 3, 6, 9, 14, 20 e 23 foram reportados como possuidores de QTL com efeitos pleiotrópicos sobre características múltiplas de produção de leite. De maneira similar Smaragdov (2006), em um estudo de mapeamento genético de locos responsáveis para características de produção em gado de leite, encontrou que o número de QTL e as posições estavam especialmente nos cromossomos 1, 3, 6, 14, 20 e 23.
Chen et ál. (2006), utilizando 14 marcadores microssatélites numa região com cobertura de 63,5 cM no cromossomo 6 bovino, em uma população da raça Holandesa Chinês, mediante delineamento de filhas por meio de 26 famílias, com as 2 famílias mais significativas e com um modelo de QTL fixado, encontraram um QTL afetando produção de gordura, perto do marcador BMS470 (32 cM), com um nível de significância 5% genômico; com o modelo de 2 QTL fixados encontraram dois QTL afetando as três características de produção: leite, proteína e gordura.
Conclusões
Considerando todas as famílias para porcentagem de sólidos totais, um QTL foi identificado com P< 0,05 e F = 2,34 na posição 58 cM, próximo ao marcador MNB-208 (60,21 cM). Ao empregar as famílias 3 e 4, as mais significativas, o mesmo QTL foi identificado com P < 0,01 e F = 7,61, mas na posição 58 cM, próximo ao marcador MNB-208 (figura 2).
Foram identificados novos marcadores microssatélites com alta Ho e PIC, para ser empregados em estudos de identificação de QTL, identidade genética e teste de paternidade, principalmente nas raças zebuínas que requerem ser mais exploradas com a genética molecular.
Embora exista uma grande diversidade genética observada entre raças taurinas e indianas, os QTL encontrados no cromossomo 6 da raça Gir relacionados com características de produção de leite são conservados, o que prova indiretamente que alelos de genes candidatos relacionados com essas características de produção no mesmo cromossomo foram fixados.
Bibliografia
Chen, H. Y., Zhang, Q., Yin, C. C., Wang, C. K., Gong, W. J. and Mei, G. "Detection of Quantitative Trait Loci Affecting Milk Production Traits on Bovine Chromosome 6 in a Chinese Holstein Population by the Daughter Design". Journal Animal Science 89 (2006): 782-790. [ Links ]
Churchill, G. A. and Doerge, R. W. "Empirical treshold values for quantitative trait mapping". Genetics 138 (1994): 963-971. [ Links ]
Georges, M. D., Nielsen, M., Mackinnon, A., Mishra, R., Okimoto, A. T., Pasquino, L. S., Sargeant, A., Sorensen, M. R., Steele, X., Zhao, J., Womack, E. and Hoeschele, I. "Mapping Quantitative Trait Loci Controlling Milk Production in Dairy Cattle by Exploiting Progeny Testing". Genetics 139. (1995): 907-920. [ Links ]
Green, P., Falls, K. and Crooks, S. CRIMAP Documentation. Version 2.4. St. Louis: Washington University School of Medicine, 1990. [ Links ]
Haley, C. S. and Knott, S. A. "A simple regression method for mapping quantitative loci in line crosses using flanking markers". Heredity 69. (1992): 315-324. [ Links ]
Khatkar, M. S., Thomson, P. C., Tammen, I. and Raadsma, H. W. "Quantitative Trait Loci Mapping in Dairy Cattle: Review and Meta-Analysis". Genetics. Selection. Evolution 36. (2004): 163-190. [ Links ]
Lynch, M. and Walsh, B. Genetics and Analysis of Quantitative Traits. Sunderland/Massachusetts: 9809, 1998. [ Links ]
Madalena, F. E. Estratégias de uso de recursos genéticos visando melhorar a qualidade do leite e derivados. VII Simpósio Brasileiro de Melhoramento Animal. São Carlos, SP, 10 e 11 de julho de 2008. [ Links ]
Martínez, M. L. e Verneque, R. S. Programa nacional de melhoramento genético. Revista Balde Branco 439. (2001): 8-10. [ Links ]
Neimann-Sorensen, A. and Robertson, A. "The Association between Blood Groups and Several Production Characteristics in Three Danish Cattle Breeds". Acta Agri. Scand. 11. (1961): 163-196. [ Links ]
Ron, M., Blank, Y., Band, M., Ezra, E. and Weller, J. I. "Misidentification rate in Israeli Dairy Cattle Population and its Implications for Genetic Improvement". Journal Dairy Science 79. (1996):676-681. [ Links ]
Ron, M., Kliger, D., Feldmesser, E., Seroussi, E.; Ezra, E. and Weller, J. I. "Multiple Quantitative Trait Locus Analysis of Bovine Chromosome 6 in The Israeli Holstein Population by a Daughter Design". Genetics 159. (2001): 727-735. [ Links ]
Sambrook, J. and Russell, D. W. Molecular Cloning: A Laboratory Manual. Third edition. Cold Spring Harbor: Cold Spring Harbor Laboratory Press, 2001. [ Links ]
Seaton, G., Hernández, J., Grunchec, J. A., White, I., Allen, J., De Koning, D. J., Wei, W., Berry, D., Haley, C. and Knott, S. "GridQTL: A Grid Portal for QTL Mapping of Compute Intensive Datasets". Proc. 8th World Congress on Genetics Applied to Livestock Production, Belo Horizonte, MG, Brasil, 2006. [ Links ]
Smaragdov, M. G. "Genetic Mapping of Loci Responsible for Milk Production Traits in Dairy Cattle". Russian Journal of Genetics 42 (1). (2006): 1-12. [ Links ]

