











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIntroduction
In the search for sustainable sources of energy, plant materials for the production of biofuel have recently attracted attention. Jatropha curcas (Euphorbiaceae) produces seeds containing 27-43% inedible oil that can be converted into biodiesel (Chhetri et al., 2008). This shrub adapts easily to various tropical climatic conditions and can grow on marginal lands of low fertility and does not compete with traditional crops (Matos et al., 2014). These characteristics extend perspectives for the plant's cultivation in northwestern South America. However, these features (low requirements in mineral nutrients and water, low labor demand, and tolerance for salinity, pests and diseases) were recorded for genotypes from the wild. Little information on these characteristics is available in the field under Colombian conditions (Campuzano, 2008; Arévalo et al., 2011).
One of the factors that limits growth and dry mass production in J. curcas is a deficiency of macronutrients in the soil (Silva et al., 2009; Ahmed et al., 2016). Fertilization that is nitrogen-poor could severely diminish biomass production in J. curcas, reducing plant height and affecting leaf formation (Wang et al., 2011). Mohapatra and Panda (2011) found that growth and yield of J. curcas in poor soils were highly influenced by the application of NPK fertilizers. According to Kalannavar et al. (2009), seed yield in J. curcas treated with 100-100-150 kg ha-1 per year of N-P2O5-K2O increases 4.5-fold, reaching 3,937 kg seeds ha-1 per year as compared to absolute control without fertilization (875 kg ha-1 per year). Also, Patil and Parameshwarappa (2007) report significant increases in plant height, basal stem diameter, number of branches, and a respective seed yield of 1,475 kg ha-1 per year following the application of 80-80-80 kg ha-1 N-P2O5-K2O. In Thailand, a significant increase in seed yield up to 1,559 kg ha-1 per year was achieved in plants of 75 cm height and fertilized with 15-15-15 N-P2O5-K2O at a rate of 312.5 kg ha-1 (Suriharn et al., 2011). Several studies carried out in Brazil emphasize the need for N fertilization in J. curcas, mainly due to the high losses of N caused by its volatilization and leaching in tropical soils (Collier et al., 2006; Lara and Souza, 2008).
Plant height (Hp) and basal stem diameter (BSD) are used in J. curcas to estimate plant biomass as reported by Makungwa et al. (2013), who obtained an allometric model to estimate woody biomass using BSD under field conditions. These authors do not recommend a generalized application of models since these are determined for each specific site and, as far as possible, according to the authors should be applied on fully developed trees. In the wild this species can reach a height of more than 5 m with stems of a sympodial growth, since several stems produce branches close to the soil surface (Tigere et al., 2006). Bártoli (2008) states that a J. curcas crop should be pruned to a maximum of 2.5 m height to induce lateral branching. This increases the number of fruits per plant, facilitates control of pests and weeds, and provides easy harvesting by gaining access to all sectors of the crop canopy. According to de Lima et al. (2016), J. curcas tends to increase height at high sowing densities due to interspecific competition, a condition that accelerates plant development and fruit production in this species. The same authors obtain the minimum seed yield per plant at the highest sowing density (2,500 plants ha-1). In this context, plant height is determined by agronomic practices that influence growth and dry mass allocation and in J. curcas has a direct influence on crop yield (Tjeuw et al., 2015).
In Colombia, there are few studies on J. curcas growth. Pedraza and Cayón (2010) evaluate environmental effects on growth and plant morphometry of J. curcas (Brazil ecotypes) in the provinces of Vichada and Santander. The plants at these locations did not significantly differ in height or number of secondary stems per plant; however, leaf dry weight and leaf area vary according to leaf strata (Pedraza and Cayón, 2010). These results suggest that J. curcas could increase leaf area to intercept more solar radiation, an aspect reported in other woody species of C3 photosynthesis (Larcher, 2003).
More than 600,000 ha of land in Colombia are considered to be appropriate for the cultivation of J. curcas, with the upper Magdalena region of the country exhibiting high potential for crop production (Campuzano, 2008). The growth response of J. curcas to fertilizer treatments should be studied with emphasis on arid soils of low fertility, such as the ones present in this region. The objective of the present study was to evaluate the effect of N and K rates on Hp and BSD of J. curcas under agroclimatic conditions of Colombian tropical dry forests in order to contribute to agronomic practices that would allow predictable levels of productivity.
Materials and methods
Experimental site and plant material
The study was conducted between March 2012 and August 2013 beginning from planting at day one to day 435 of field growth. The research took place at the Nataima Experimental Center, Colombian Corporation of Agricultural Research (AGROSAVIA) located in Espinal (Tolima province, Colombia). The crop was established in a tropical dry forest (bs-T) zone according to Holdridge (1967). The area was located at coordinates 4°11'14" N and 74°57'22" W at 371 m a.s.l. and characterized by mean 28°C air temperature, 70% relative air humidity, and 1,270 mm annual precipitation (IDEAM, 2017).
Jatropha curcas seeds of elite ecotype M-3 were obtained from the "Genetic Improvement Program of Jatropha curcas" of Corpoica 2006-2011. The seeds were planted in black plastic bags with a 4 kg capacity and were placed under a plastic mesh providing 60% shading. The substrate was a 2:1:1 mixture (v/v/v) of sandy loam soil: rice husk 60% burnt: compost. The substrate had pH 5.5, 4.1% organic matter, and available contents of 67.7 mg kg-1 phosphorus and 0.55 cmol+ kg-1 potassium.
Sprinkler irrigation was applied twice a week and seedlings emerged 4 d after planting (DAP). Eight days later, the plastic mesh was removed and seedlings were left free to solar exposure. At 45 DAP the plants had between 5 and 6 true leaves and a height of 35 cm and were transplanted into a field during the rainy season that was characterized by a mean air temperature of 28°C and a mean precipitation rate of 80 mm m-2. From May 2012 to July 2013, the mean air temperature was 27.8°C and relative air humidity oscillated between 70 and 85%. In June-August 2012, the experimental area had supplementary irrigation to avoid a water deficit balance. During 2013, the dry season had low precipitation during January and February (28.9°C mean air temperature and 65.3 mm m-2 monthly precipitation) and a rainy period during March-May had a mean air temperature of 27.0°C and a monthly precipitation of 160.4 mm m-2.
Plant establishment in the field and fertilizer treatments
The planting distance in the field was 3x2 m which is considered adequate for monoculture of J. curcas for providing high yields, reducing intraspecific competition, and facilitating fruit harvest and weed control (Ghosh et al., 2007). A randomized complete block design was established with 4 replicates and 24 plants per experimental unit, separated by a border row, for a total of 1,184 plants ha-1. The experiment was carried out in an inceptisol sandy loam soil identified as Typic Haplusterts with 1.2% organic matter (Tab. 1) and good drainage capacity.
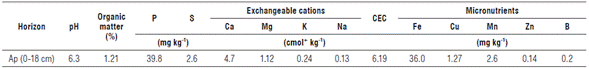
Considering the soil characteristics and responses to fertilizer applications reported for J. curcas in earlier studies (Patil and Parameshwarappa, 2007; Kalannavar et al., 2009; Suriharn et al., 2011), the doses of fertilizers were established between 0 and 150 kg ha-1 per year for N and between 0 and 180 kg ha-1 per year for K2O. These doses were arranged in a scheme comprising 12 treatments including control treatments with either N or K applied and an absolute control without NK application (Tab. 2). The sources of fertilizers were urea and potassium chloride. Other mineral elements were applied pre-transplant in equal doses for all treatments: 46 kg ha-1 P2O5, 44 kg ha-1 Ca, 18.8 kg ha-1 Mg, 27.2 kg ha-1 S, 3.18 kg ha-1 Mn, 2.4 kg ha-1 Zn, and 1 kg ha-1 B.

The annual doses of fertilizers in each treatment were fractioned into four applications starting one month after transplanting; fertilizers were applied evenly around the stem and covered with a soil layer of approximately 1 cm. Weed control was accomplished manually and complemented with a tractor-operated drag scraper between the rows. No pruning was performed on plants during the development of the experiment.
Plant sampling
Five destructive plant samplings (45 d after each fertilization) were carried out at 75, 165, 255, 345, and 435 DAP. Four plants were evaluated in each treatment for a total of 48 plants at each sampling moment. The plants were cut at soil level and plant height (Hp) was measured (cm) from the stem base at soil level to the apex of the principal orthotropic stem. Basal stem diameter (BSD) was measured at the base of the principal stem with an electronic digital caliper (Mitutoyo Absolute Digital Caliper 500-197-20, Japan).
Data analysis
Statistical analyses involved descriptive and inferential components using bivariate and univariate approaches. Initially, descriptive statistics (mean, standard error of mean, and standard deviation) for Hp and BSD were applied for each treatment and each moment of evaluation associated with factors of N and K2O doses. A three-dimensional dispersion diagram visualized Hp and BSD at each evaluation time for two contrasting treatments, absolute control and a treatment combining high rates of N (150 kg ha-1) and K2O (120 kg ha-1). The experiment involved a repeated measures design with inter-subject factors of fertilization rates (K2O dose was nested in N dose due to the origin of the K2O dose, so that different levels of the K2O factor shared a single level of the N factor) and an intra-subject factor of evaluation time (75, 165, 255, 345, and 435 DAP) (Davis, 2002; Montgomery, 2013). The Hp and BSD were used as response variables. A bivariate analysis of variance was applied to adjusted linear model. P values less than 5% were considered significant but the effects with significance less than 10% were taken into account; this was done in order not to rule out any effect that might be significant in the model. With simultaneous confidence intervals of Bonferroni, the evaluation moments were compared using error bars, since it is usual in growth models to search for significant interactions between inter-subject factors and evaluation time. Comparisons using error bars were made on the response profiles for each factor (Cumming et al., 2007). To complement the descriptive analysis, bar diagrams for Hp and BSD were constructed at the end of the evaluation period (435 DAP), combining factors to generate a single label of "treatments". This was done to obtain information on the behavior of 12 treatments (Tab. 2) at the end of the experiment for each response variable separately; in this case, the Tukey's test (P<0.05) was applied to compare the mean values. Finally, a multiple regression model was fitted for each response variable (Hp, BSD) using as predictors the N and of K2O doses and the moments of evaluation; this was done only by predictive interest and not to compare the effects. With this, while setting the evaluation time to 435 d, a cross-table was generated to present the estimates of combinations of N and K2O doses that were not originally used in the experiment.
Results and discussion
Effects of N and K applications on plant height and stem diameter
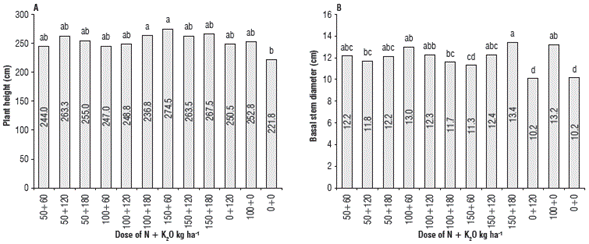
The vegetative growth phase of J. curcas in the field comprised 165 DAP indicating the moment at which the first flowering appeared. Mean Hp of J. curcas varied significantly between absolute control and the treatments with joint application of high N doses (100 or 150 kg ha-1) and K2O (120 or 180 kg ha-1) at 75, 165, 255, and 345 DAP, while at 435 DAP these differences were not observed (Tab. 3). The BSD had minimum values i n plants deprived of N fertilization throughout the experiment (Tab. 3). For both Hp and BSD variables, their behavior against NK rates presented significant differences at 435 DAP, in which the absolute control had the lowest Hp and BSD (Fig. 1). In the final evaluation, the plants treated with the highest doses of N and K2O had maximum Hp and BSD compared to absolute control (Fig. 1).
TABLE 3 Average plant heights (Hp, cm) and basal stem diameters (BSD, cm) of J. curcas at five evaluation moments.
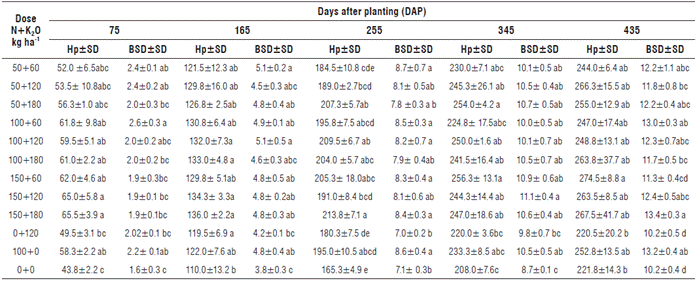
Hp - Plant height (cm), BSD - Basal stem diameter (cm), SD - standard deviation. Letters following standard deviations of each treatment identify the homogeneous groups generated from multiple comparisons (Tukey's test, P <0.05).
A three-dimensional diagram indicated the simultaneous relationship between variables Hp and BSD as a function of fertilizer rates and moments of evaluation (Fig. 2). Two treatments contrasting in Hp and BSD behavior, such as absolute control and N=150 kg ha-1 plus K2O=120 kg ha-1 (Tab. 3 and Fig. 1), were used in the diagram. It showed progressive increases in each variable in time and confirmed that plants were taller and thicker when treated with NK, while the lowest Hp and BSD were observed in absolute control without NK fertilization (Fig. 2). The highest dispersion in variables Hp and BSD occurred in the reproductive phase of growth at 345 and 435 DAP. Both variables possessed a similar growth pattern over time, as shown by Reis et al. (2018), but stood out in the treatment with the highest rate of N (150 kg ha-1) at 345 and 435 DAP. The diagram of dispersion did not reflect a trend, since the variable "treatments" was of nominal scale; however, an increasing trend in time for Hp and BSD could be observed for both treatments (Fig. 2).
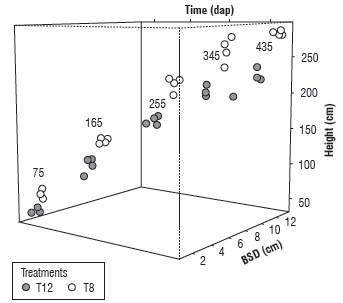
FIGURE 2 Three-dimensional dispersion diagram of plant height (cm) and basal stem diameter (cm) according to the moment of evaluation (DAP) and N+K2O treatments (kg ha-1). T8 denotes the treatment N=150 kg ha-1 plus K2O=120 kg ha-1 and T12 corresponds to absolute control.
To evaluate the effect of N and of K2O doses nested in N (K2O(N)) on Hp and BSD at different moments of growth, Wilks' Lambda statistics was used. It revealed the interactions of second and third order, both significant (P<0.05) and highly significant (P<0.01) for the main effects associated with doses of N, doses of K2O nested in N, and evaluation moments (Tab. 4). In order to use the results of the bivariate analysis of variance, it was necessary to recognize the presence of significant interactions; these interactions made it impossible to interpret the main effects (Tab. 4), so that one could appeal for the graphs of interaction (Sokal and Rohlf, 2012). Table 4 presents the summary of the two-way ANOVA of repeated measures design of some of the effects. It highlights the interactions and justifies the use of interaction profiles to select the best N doses at each evaluation time. Table 4 shows the effects of the reduced model eliminating effects with a significance level greater than 5%. The values close to 5% were left in the model, although they were higher than this limit. In double interactions of inter-subject factors with "time", the effect of K2O(N) could be interpreted, since the interaction was not significant (P=0.0589) (Tab. 4). In the absence of a significant interaction between "time" and "dose of K2O nested in N", the values of Hp and BSD at 435 DAP corresponded to 266.7 and 12.2 cm, respectively, for 180 kg ha-1 K2O nested in different N doses, while control plants had Hp of 239.1 cm and BSD of 11.2 cm. As for the effect of N, the interaction of "N dose" with "time" was revealed. This makes it impossible to select the best dose of N for all moments, so that the use of profiles facilitated choosing the best dose at each evaluation time (Figs. 3 and 4).
TABLE 4 Significance levels for individual effects and interactions in bi-variate analysis of variance.
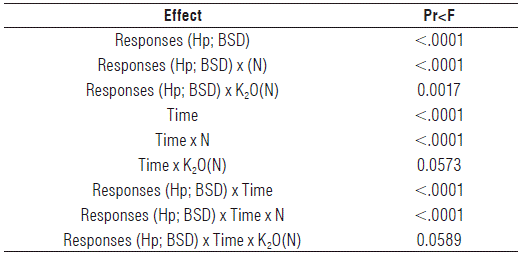
Hp - plant height (cm); BSD - basal stem diameter (cm); N - dose of N; K2O(N) - dose of K2O nested in dose of N. The effects, whose significance was higher than 5%, were not considered. The effects close to 5% significance were left in the model, although being higher than this limit.
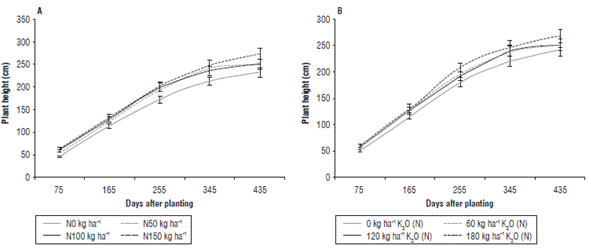
FIGURE 3 Plant height (cm) of J. curcas affected by different doses of N (A) and doses of K2O nested in N (B). The bars correspond to standard errors of the means.
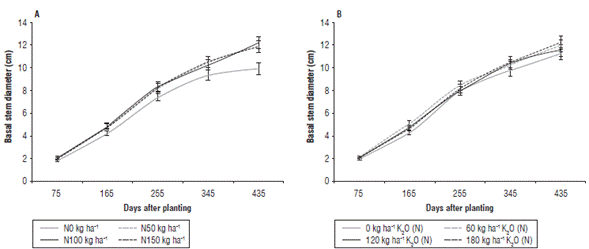
FIGURE 4 Basal stem diameter (cm) of J. curcas affected by different doses of N (A) and doses of K2O nested in N (B). The bars correspond to standard errors of the means.
The behavior of Hp discriminated by moments of evaluation (DAP), dose of N, and dose of K2O nested in N confirmed the major increases in plant height of J. curcas during the vegetative phase between 75 and 255 DAP (Fig. 3), a period when plants allocated most of their carbon resources to new branches and leaves. According to the slopes of the curves, the gain in height of 0.78 cm/day was observed for plants treated with N=150 kg ha-1 plus K2O=180 kg ha-1, while the absolute control gained 0.71 cm/day in height (Fig. 3A). A slower growth was detected during the reproductive phase between 345 and 435 DAP, probably due to translocation of nutrients from leaves, branches, and stem towards flowers and fruits (Larcher, 2003). The plants gained only 0.35 cm/day in height in this period when treated with 150 kg ha-1 N and 180 kg ha-1 K2O and only 0.17 cm/day in control plants without NK fertilization (Fig. 3A). The plants were taller in all treatments with N application presenting the highest Hp at all moments of the evaluation and reaching 285 cm at 435 DAP when treated with 150 kg ha-1 N. In contrast, the lowest heights of 228 cm corresponded to the treatments without N (Fig. 3A), which agrees with Su et al. (2013), who reports increases in height, stem diameter, and total number of branches in J. curcas as the doses of 15-15-15 NPK fertilizer increase. However, very high doses of N could prolong the vegetative phase of growth, thus, delaying the start of fruit production (Larcher, 2003). Likewise, increases in height above 2.5 m might affect agronomic practices in J. curcas by requiring pruning and obstructing crop harvest, since fertilization could affect branching in J. curcas (Nirala et al., 2017). When analyzing the effect of the K2O dose nested in N (K2O(N)) on Hp, a similar trend was observed, with the largest heights (267 cm) obtained with application of 180 kg ha-1 K2O for the average N doses, whereas the lowest heights (239 cm) corresponded to control plants without K application (Fig. 3B).
The plants were not significantly different in BSD during the first two evaluation moments according to values of standard errors (Fig. 4). It should be noted that BSD increased progressively in time. Thus, starting from 165 DAP all plants treated with NK differed in BSD from the absolute control; however, no differences in BSD were detected among the plants treated with non-zero doses of N or K (Fig. 4A). A similar result in BSD was observed affected by K2O dose nested in N but with fewer differences, with smaller BSD corresponding to control plants without K fertilization (Fig. 4B).
Multiple regression models for prediction of plant height and stem diameter
Multiple regression models for Hp and BSD were developed. These permitted predicting responses according to the levels of each factor and the development of two formulae as presented in Table 5. The models served to assess untested levels of factors (dose of N or dose of K2O or moment of evaluation) fitting these within their range of operation (an example highlighted in Table 6). According to the multiple linear regression analysis, the behavior of Hp fit this model well and explained 92.99% of the variability and, analogously, the model for BSD adjusted to 94.68% of the variability. These mathematical models allow estimating Hp and BSD for any dose of N and K2O at five evaluation moments within the evaluated range (Tab. 5). Therefore, N and K fertilization rates can be adjusted to a desired plant height and, thereby, they can be used to schedule some agronomic labors, such as pruning. In J. curcas, Hp and BSD could be further used to estimate the plant biomass (Makungwa et al., 2013).
TABLE 5 Multiple regression models for the prediction of plant height and basal stem diameter of J. curcas as a function of N dose (kg ha1), K2O dose (kg ha-1), and moment of evaluation (DAP).

Range of settings of prediction models: 0 ≤ K2O ≤ 180, 0 ≤ N ≤ 150, 75 ≤ t ≤ 435, where K2O - dose of K2O (kg ha-1), N - dose of N (kg ha-1), t - time (DAP). Ŷ corresponds to estimated model and R2 denotes the adjusted coefficient of determination.
Hp had a highly significant (P<5%) correlation with BSD. As the doses of N and K increased, both Hp and BSD increased progressively, an aspect observed for all moments of evaluation. These allometric relationships vary considerably by species (Clough and Scott, 1989), but are especially important for trees (Lott et al., 2000; Wang et al., 2000; Salis et al., 2006; Levia, 2008), where stem diameter is used to calculate some parameters, such as plant height and biomass, an aspect that could be used in J. curcas. In this sense, tree growth is supported by the trunk and must be biomechanically balanced (Montagu et al., 2005); in other words, stem diameter is related to the length and distribution of branches and should prevent uprooting or branch split. In J. curcas, stem diameter and number of branches (Laviola et al., 2012) as well as plant height (Laviola et al., 2012; Shabanimofrad et al., 2013; De Lima et al., 2016) were shown to correlate positively with seed yield per plant.
Employing the models obtained, the values of Hp and BSD for J. curcas at 435 DAP were estimated for different levels of N and K fertilizers. Also, randomly chosen and experimentally untested doses (N=75 kg ha-1 and K2O=90 kg ha-1) were introduced within the operational ranges of each dose to verify the predictive behavior of the models (Tab. 6).
TABLE 6 Plant height (cm) and basal stem diameter (cm) of J. curcas estimated at 435 DAP with different combinations of N and K2O doses including a dose not tested experimentally (highlighted).

The values estimated by the models for both evaluated doses and untested doses experimentally showed a clear increase in Hp and BSD for J. curcas with increasing doses of N and/or K fertilizer (Tab. 6). Therefore, these models could be applied for easy, rapid and non-invasive predictions of J. curcas growth in the field for the region.
Conclusions
The growth of J. curcas was influenced by doses of N and doses of K2O nested in N during the vegetative and reproductive phases. The tallest plants with the largest stem diameters were those subjected to the highest rates of N and K fertilizers. For the management of fertilization of J. curcas in the given edaphoclimatic conditions of Colombia, doses of 100 kg ha-1 N and 120 kg ha-1 K2O can be recommended. Both basal stem diameter and plant height in J. curcas increased proportionally during growth. The predictive models for plant height and basal stem diameter integrated rates of N, rates of K2O, and time had an adjusted coefficient of determination R2 over 92%. The fertilizer rates N=75 kg ha-1 and K2O=90 kg ha-1 were not evaluated in the field, but they fitted well within the operational ranges of the models and data generated were consistent with the behavioral tendencies of the studied variables. The models obtained could be used for rapid and non-invasive predictions of J. curcas growth in the field.

