Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIntroduction
Climate change affects agriculture worldwide and Chile is also compromised. This context demands a higher water use efficiency through improved technologies and more efficient water management in the field (McKersie, 2015; Boisier et al., 2018). There is a growing interest in irrigation with saline water, which offers a broad possibility to start intensive and commercial cultivation of various halophyte species (Basílio et al., 2018; Mota et al., 2021). Glassworts (genera Salicornia and Sarcocornia of the Amaranthaceae family) are halophytes widely distributed and studied as edible and non-edible crops that can be used for rehabilitation of wetlands, glass manufacture, and ruminant and human consumption. The edible species have different characteristics such as a low quantity of external inputs for their cultivation, high nutritional value, and pharmaceutical benefits, turning these crops into a sustainable food source in the context of climate change (Patel, 2016; Loconsole et al., 2019). Moreover, other species of the Sarcocornia genus have been recently described as a good source of antioxidants (Antunes et al., 2021).
Chilean coastal marshes are able to develop sustainable halophyte agriculture with low financial involvement. This provides opportunities for productive diversification and resilience to small-scale fisheries and families of coastal farmers within the integrated aquaculture systems (Buschmann et al., 1996). There are three species of wild glassworts in Chile, all belonging to the genus Sarcocornia. Sarcocornia neei (Lag.) M.A. Alonso & M.B. Crespo is the only species distributed along the coast. The other two species (S. pulvinata and S. magellanica) are adapted to extreme environments and could be found in the Atacama Desert and Patagonia, respectively (Alonso & Crespo, 2008). S. neei has the potential to be cultivated in many bays along the coastline of Chile.
S. neei is a small shrub with a height up to 80 cm, erect to decumbent stem, and wide morphological variations in its habitats (Alonso & Crespo, 2008). Its potential for human consumption has been barely studied. However, recent studies have been carried out to determine the seed germination requirements and nutritional value of the wild and cultivated plants as well as the plant's potential to serve as an indicator of heavy metal pollution in central Chile (Riquelme et al., 2016; Alonso et al., 2017; Meza et al., 2018). In southern Chile, the wild plant sprouts are collected in summer. Additionally, the locals let their cattle graze on the S. neei natural pastures in coastal marshes. These informal practices are evidence of both the potential of S. neei as an emerging crop and the need to develop agricultural practices for its use while avoiding the ecological depletion of these fragile ecosystems.
The cultivation of S. neei could be one of the first attempts to establish saline agriculture in Chile. Thus, this research aimed to study the yield and phenology of the halophyte S. neei as a potential novel crop in Chile. This study evaluated the effect of different fertilizer treatments on the species performance under controlled and natural conditions.
Materials and methods
Plant material and location
Whole plants were collected from the coastal marshes of Chamiza, Puerto Montt, Chile (41°29'18.9" S, 72°51'45.8" W). Trials were established in a nearby location (Lenca, 41°36'20.7" S, 72°41'20.6" W) characterized by a temperate rainy climate with coastal influence according to Köppen classification (Sarricolea et al., 2017). The average temperature is 10.6°C; the warmest month, January, has an average maximum temperature of 19.8°C and the coolest month, July, has an average minimum temperature of 3.5°C. The total average rainfall reaches up to 1,613 mm with 218 rainy days (World Weather Information Service, 2018).
Trial management
Two trials were established, the first one in a tunnel-type greenhouse (Fig. 1F) and the second one in the open field next to the greenhouse. The maximum temperature inside the greenhouse was 41.9°C in January and the average air humidity was 90% in summer. Each trial consisted of nine platforms of 3.2 m long, 1.2 m wide, and 0.2 m high, with an individual area of 3.84 m2 containing a mixture of 50% decomposed leaf litter with soil and 50% sand. Plants were collected directly from the marsh and the stems were cut 10 cm above the crown before planting. Within the platforms, the plants were arranged at a distance of 0.35 x 0.40 m from each other (16 plants per platform, Fig. 1E) to distinguish each plant at harvest. Both trials had the same fertilizer treatments: 40 g N per plant, 80 g N per plant, and a control without fertilization, with three replicates of 16 plants each. The fertilizer source was potassium nitrate (15 N - 0 P - 17 K), and 50% of the total dose was applied at plant establishment (November 2018) in each treatment. The second half of the N fertilizer was split into three applications, and each dose was applied two weeks after each harvest. All platforms received the same management and were irrigated with saline water (30 g L-1 NaCl) according to soil friability. The total water volume applied through the entire period was 3,046 L. Soil analysis at the end of the trial revealed a sandy soil texture, 88.4% sand, 5.4% clay, 6.2% silt, pH 6.5, and cation exchange capacity (CEC) of 45.65 cmol kg-1.

Yield evaluation
All plants were cut in January 2018 to obtain uniform growth before the yield evaluation. This practice was not considered for the analysis but employed the same criterion used for the regular harvest. Thus, for yield evaluation, four plants from each platform were randomly selected; the same plants were harvested in three seasons: April (fall harvest), September (spring harvest), and February 2019 (summer harvest). Based on preliminary results, we determined the best timepoint to harvest when 50% of the shoots were larger than 10 cm. The plants were harvested by collecting the above-ground vegetative biomass, including all plant parts located 5 cm above the crown, where the roots originate. Results were expressed in grams of fresh matter per plant (g FM per plant). Then, all plant material was dried in a drying chamber (model F23, BINDER, Tuttlingen, Germany) at 70°C until constant weight to determine the water content (%) and dry matter weight per plant (g DM per plant).
Determination of phenological stages
Four plants per replicate were randomly chosen (excluding those selected for yield evaluation) and examined once a week to determine the phenological stages throughout the seasons; these were identified when 50% of the chosen plants in each trial had the same stage. Trained personnel determined the phenological stages by visual inspection, based on the available bibliography (Alonso & Crespo, 2008). Additionally, wild plants growing in Lenca coastal marsh were assessed once a week along a random 100 m transect to compare the phenological stages in both experimental trials with those recorded in wild plants. Winter dormancy period (w, plants with no vegetative growth), vegetative growth (v, plants with active growth but without flowers), flowering (f, plants at flowering), and fructification (x, plants with fruits) were registered.
Statistical analysis
A two-way analysis of variance (ANOVA) was performed for the experimental data, considering the environment (open field, greenhouse) and fertilizer treatments (0, 40, and 80 g N); the effects were considered significant with a P-value <0.05. The Tukey's test was used to compare the treatments for each analysis. A repeated measures analysis was performed to analyze the effect of the seasons. Statistical analyses were performed using the software R version 3.4.1.
Results and discussion
Yield evaluation
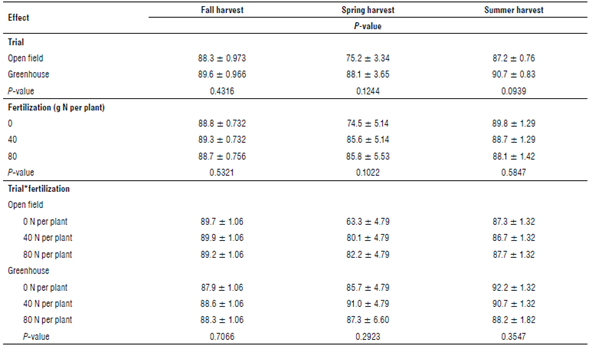
The yields were significantly different between the open field and greenhouse environments in all studied seasons. The highest yield was obtained in the greenhouse trial in summer, reaching 45.33 g DM per plant in the treatment with 40 g N per plant, which significantly differed from the 15.25 g DM per plant obtained in the same treatment in the open field (P<0.001). The treatment with 40 g N per plant yielded the highest biomass for both trials and at all harvests (Tab. 1). Table 1 shows the P-values of the analysis and the interaction between the environment, fertilization level, and harvest season. An experiment carried out in Brazil obtained 21.4 g FM per plant in an aquaculture system (Pinheiro et al., 2020). Our results indicated that, in the 40 g N treatment, the fresh biomass reached 462.5 g per plant in summer. Therefore, the area near Lenca could be a suitable environment to establish a crop of halophytes under temperate conditions.
TABLE 1 Mean yield of the Sarcocornia neii shoots (g dry matter per plant ± standard error) in each trial, fertilization treatment, and trial*fertilization at the fall, spring, and summer harvests.
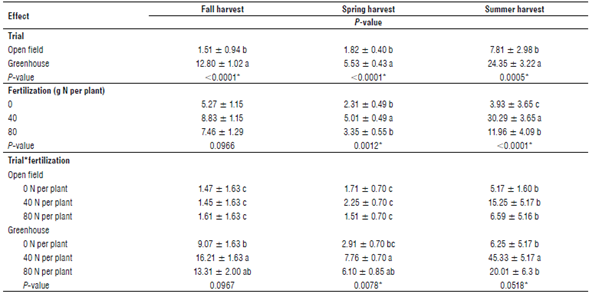
Different letters indicate significant differences between treatments or trials according to the Tukey's test (P<0.05). Asterisks indicate a P-value lower than 0.05.
Greenhouse trial
The greenhouse environment showed no differences between treatments in the fall, with a mean of 12.80 g DM per plant (Tab. 1). The spring harvest obtained a significantly higher yield in the 40 g N treatment and, surprisingly, the yields from the plants fertilized with 80 g N were no different from those treated with 0 g N. Finally, the summer harvest in the 40 g N treatment showed the highest yield in the experiment, which was significantly different from the yields obtained in the 0 and 80 g N treatments (Tab. 1). The season effect was significant in this trial, with the highest yield registered in summer, followed by the fall and spring yields (P<0.001). The greenhouse-grown plants collected at the three harvests showed an average water content of 90.2% (Supplementary material 1). The treatment with 40 g N produced the highest yield at all harvests. This confirms a need for the agronomic management of S. neei. Also, the yields were lower in the 80 g N treatment than in the 40 g N treatment, probably because of phytotoxicity. Recently, ammonium regulation was found to be an important trait in Salicornia europaea, and its detoxification plays a role in the tolerance to high salinity in these halophytes (Ma et al., 2020).
Open field trial
The open field trial showed no differences between the fertilizer treatments in all seasons, obtaining a mean of 3.71 g DM per plant. However, the season effect was significant, revealing the differences between the fall, spring, and summer harvests (Tab. 1). The plants from the open field trial had an average water content of 88.3% when all samples were pooled (Supplementary material 1).
Phenological stages
Figure 1 shows the four phenological stages observed in Lenca: winter dormant period (w), vegetative growth (v), flowering (f), and fructification (x).
Vegetative growth in the open field trial extended for six months, from July to December. Then, flowering occurred during January and February, and fructification was observed in March and April. The winter dormancy period was reduced in the plants from the greenhouse trial compared to wild plants, and it only lasted two months from May to June. The greenhouse conditions forced plants to avoid winter dormancy, flowering, and fructification periods during the season 2017-2018, and the vegetative growth was maintained throughout the year. According to previous studies, phenological stages are conditioned by day length and repetitive harvest regime (Ventura et al., 2011). Thus, our results demonstrated better crop performance in the greenhouse trial since commercial standards for S. neei depend on its vegetative growth and a requirement of year-round cultivation.
Conclusion
The results confirm the potential of the south of Chile to diversify its commercial matrix, through the inclusion of halophyte cultivation in greenhouses in the coastal areas. Here, we introduced the general framework to establish a successful crop, including the fertilization management and better environment. However, further research is needed to fully understand the challenges of this new crop, especially related to the inputs required through the different stages of crop development and the sustainability of the production. Overall, halophytes could become new crops to support the development of small farmers and fishermen along the coast of Chile.