










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkCaldasia
versão impressa ISSN 0366-5232versão On-line ISSN 2357-3759
Caldasia v.28 n.2 Bogotá dez. 2006
LA COMUNIDAD ÍCTICA DEL RÍO MESAY DURANTE EL PERIODO DE AGUAS ALTAS (CAQUETA, AMAZONIA COLOMBIANA)
The Fish Community of Mesay River during high level water period (Caquetá, Colombian Amazon)
IVONNE BEJARANO
MARÍA DEL PILAR BLANCO
JOSÉ IVÁN MOJICA
Departamento de Ciencias Marinas, Universidad de Puerto Rico, Mayagüez. PO Box 3121 Lajas 00667, PR. Puerto Rico. ibejarano@cima.uprm.edu
Universidad Nacional Autónoma de México, Posgrado en Ciencias del Mar y Limnología. México. pblanco@icmyl.unam.mx
Instituto de Ciencias Naturales, Universidad Nacional de Colombia. Apartado 7495, Bogotá, D.C., Colombia. imojica@unal.edu.co
RESUMEN
Se caracterizó la composición, la distribución y la estructura de la comunidad íctica del río Mesay y dos ambientes asociados (lagos y quebradas) durante el periodo de aguas altas. Entre julio y septiembre de 2000 se realizaron muestreos semanales en cinco estaciones representativas del área de Puerto Abeja. Se capturaron 79 especies, pertenecientes a cuatro órdenes y 16 familias utilizando trasmallo, jamas de anjeo, vara y cuerda.Veintitrés de las especies identificadas son nuevos registros para la Amazonia colombiana. No hubo patrones de distribución espacial entre biotopos. Las especies de tallas grandes abundaron en las estaciones con mayor plano de inundación, mientras las de tallas pequeñas abundaron en los ambientes lénticos. Aunque la riqueza de especies fue elevada, la diversidad fue baja posiblemente por la fuerte dominancia de pocas especies y la abundancia de especies representadas por pocos individuos.
Palabras clave. Amazonia, Colombia, Chiribiquete, Mesay, Peces.
ABSTRACT
The composition, distribution and structure of the fish community of Mesay River and two associated biotopes (lakes and streams) were studied during high level water. During July through September 2000, weekly surveys were done in five stations representative of the Puerto Abeja area. A total of 79 species belonging to four Orders and 16 families were captured by using gill nets, dip nets, hand lines, and rod and reel. 23 of the species surveyed are new registers for the Colombian Amazonic region. Although only the flooding period was sampled, species richness was still observed to be high. No spatial distribution pattern was observed between habitats. Larger sized fish species were most abundant in those stations possessing a larger flooded plain, while smaller sized fish species preferred lenthic habitats. Although fish richness was high, diversity was low possibly because of the strong dominance of few species and the elevated number of rare or infrequent species.
Key words. Amazon, Chiribiquete, Colombia, Fish, Mesay.
INTRODUCCIÓN
Los peces son el grupo de vertebrados más antiguo y numeroso. Se conocen cerca de 25 000 especies ícticas, la mayoría de ellas habitando en aguas tropicales (Nelson 1994). La cuenca amazónica presenta más riqueza de especies de peces que cualquier otro sistema fluvial en el mundo (Val & de Almeida-Val 1995). Sin embargo, este es uno de los grupos taxonómicos de vertebrados menos estudiados en esta región (Goulding et al. 1996) y particularmente en Colombia. El número de especies de peces de toda la región amazónica puede oscilar entre las 1200 y 1400 especies de peces (Géry 1984), de las cuales únicamente 264 especies han sido registradas para la Amazonia colombiana (Mojica 1999), cifra que denota la falta de exploración ictiológica en esta región del país. Debido a que la fauna íctica de la región del río Mesay es sin duda la menos conocida en la cuenca amazónica colombiana, el objetivo principal de este estudio fue caracterizar la estructura, composición y distribución de la comunidad de peces que habita el Rio Mesay y algunos ecosistemas asociados durante el periodo de aguas altas.
El río Mesay es un afluente del río Caquetá que nace en el Cerro Negro o la mesa Schultes, entre las serranías de Cuñare e Iguaje. Tiene una longitud total de 420 km, de los cuales 213 km están dentro de los límites del Parque Nacional Natural Serranía de Chiribiquete (PNNSC) (Hildebrand & Peñuela 1999). El río Mesay presenta numerosos meandros y lagos y en su recorrido por la Serranía del Chiribiquete forma un extenso conjunto de raudales y chorros. Posee un régimen hidrológico monomodal, con un período de aguas altas entre abril y septiembre, y de estiaje entre diciembre y febrero. Por sus características fisicoquímicas, corresponde a un río de aguas negras amazónicas (IGAC 1999), con valores de pH entre 5.7 y 6.1, una baja conductividad eléctrica que oscilla entre 3.3 y 5.9 µS.cm-1 y una transparencia entre 28 y 100 cm.
MATERIALES Y MÉTODOS
Este estudio se realizó en la zona del río Mesay, en el sector de Puerto Abeja, hacia la zona suroriental del PNNSC en el departamento del Caquetá, Amazonia colombiana (Figura 1). Las coordenadas geográficas de la estación científica son 0°04’27”de latitud norte y 72°27’05” de longitud oeste, y la altura es de 256 msnm.
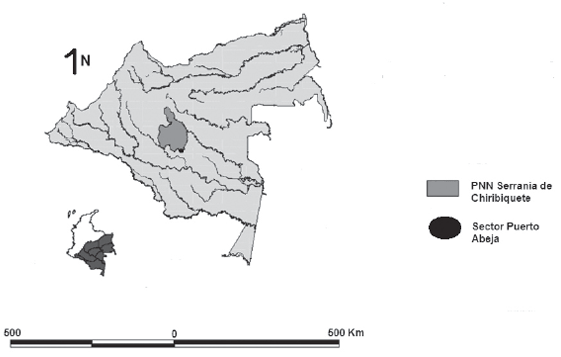
Figura 1. Ubicación geográfica y límites del Parque Nacional Natural Serranía de Chiribiquete, Puerto Abeja y Río Mesay.
El trabajo de campo se llevó a cabo entre el 2 de julio y el 18 de septiembre de 2000, en el periodo de aguas altas del río Mesay. Con el fin de abarcar diferentes ambientes acuáticos, se establecieron en campo cinco estaciones de muestreo que incluyeron al río Mesay, dos lagos asociados y dos quebradas afluentes. En cada estación se realizaron muestreos semanales, que consistieron en dos faenas diarias de pesca de cuatro horas cada una; la primera de 5:00 a 8:00 y la segunda de 16:00 a 19:00 horas. Se combinaron diferentes artes de pesca que incluyeron trasmallos de poliéster y nylon con ojo de malla de 5 y 11 cm respectivamente, redes de anjeo de marco circular y cuadrado, y cuerda y vara con anzuelos número 8, 12, 14 y 16. Se definió como muestra el conjunto de todos los individuos colectados en cada ambiente durante un día de pesca. Los ejemplares capturados se fijaron en formol al 10% y se transportaron al laboratorio de ictiología del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia en Bogotá (ICN-UNC), donde fueron determinados taxonómicamente hasta nivel de especie, cuando fue posible y posteriormente analizados e incluidos en la colección ictiológica.
Para comparar la composición de especies entre los tres biotopos analizados se utilizó el índice de Sorensen (CC), que se basa en la presencia y ausencia de las especies y está definido por la siguiente ecuación:
CC = 2 c / (s1 + s2)
c = Número de especies comunes entre los dos sitios.
s1 y s2 = Número de especies en la comunidad 1 y 2.
Con el programa PC-ORD versión 3.17 se realizaron análisis de agrupación (“cluster analysis”) basados en el coeficiente de distancia euclidiana, y la estrategia de grupo promedio, utilizando los datos de abundancia. También, se calcularon índices de diversidad (Shannon-Weaver en base dos (H’)) y riqueza (número de especies presentes en la muestra) para las muestras de cada estación y para la estación en general.
Adicionalmente se utilizó el programa EstimateS, versión 6.0b1 que incluye una serie de estimadores de riqueza basados en la abundancia relativa o en la incidencia. En este estudio se utilizaron los estimadores de riqueza de especies Chao2 y Jackknife2, debido a que se consideran los más adecuados cuando el número de muestras es reducido (menos de 50 muestras acumuladas) (Colwell & Coddington 1994). Chao2 asume los sesgos de la riqueza observada como un factor que depende de la presencia y distribución de los taxones “raros” dentro de las muestras (Heyer et al. 1999), mientras Jackknife2 se basa en el número de especies que ocurren únicamente en una muestra, así como las que aparecen en dos.
RESULTADOS
Se capturó un total de 5263 ejemplares pertenecientes a cuatro Ordenes, quince familias, 46 géneros y 79 especies. De estos individuos 817 corresponden a especies de peces de tallas grandes (con longitudes estándar de más de 10 cm) capturadas con redes y anzuelos y 4446 a individuos de especies pequeñas que se colectaron con jamas. Para su clasificación a nivel de Orden se siguió la clave propuesta por Nelson (1994), excepto para el Orden Characiformes para el que se adoptó la clasificación de Gery (1977). La lista de especies y sus respectivos números de registro en la colección ictiológica del ICN-UNC se muestra en la Tabla 1.
Tabla 1. Lista taxonómica y número de colección (ICN-MHN) de las especies ícticas del río Mesay capturadas entre los meses de julio y septiembre de 2000. Según Nelson (1994) para órdenes y se sigue a Géry (1984) para los Characiformes.



El Orden Characiformes predominó con un total de 63 especies (80%), seguido por los Siluriformes representados por nueve (11%), los Perciformes por seis (8%) y los Rajiformes por una sola especie (1%). A pesar de que se realizaron faenas de pesca al anochecer no se capturó ningún ejemplar del Orden Gymnotiformes. Se observó un predominio marcado de la familia Characidae con 33 especies que representaron el 42% del total, seguida por Serrasalmidae con trece (16%), Anostomidae con el 7.6%, Curimatidae y Pimelodidae con el 6.3% cada una, y Cichlidae con el 5% (Figura 2) Las familias Sciaenidae, Auchenipteridae, Erythrinidae y Hemiodidae en conjunto representaron un 10%, y Ageneiosidae, Aspredinidae, Characidiidae, Lebiasinidae, y Potamotrigonydae un 6.8%, cada una de ellas con un único individuo capturado.
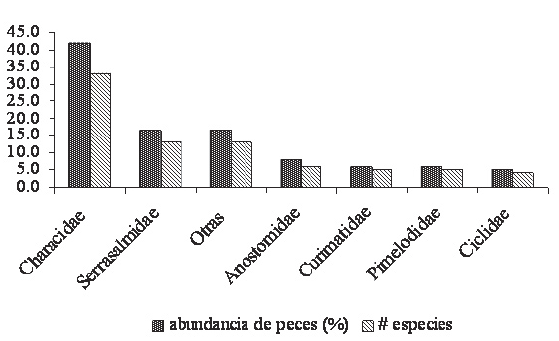
Figura 2. Abundancia de las familias ícticas según el número de especies y el porcentaje de individuos.
La composición de las especies presentes en los diferentes biotopos fue similar. La mayor similitud se presentó entre el río y los otros dos biotopos (índice de Sorensen: 63%). Los lagos y las quebradas fueron similares entre sí en un 58% (Tabla 2).
Tabla 2. Indices de similitud de Sorensen (%) entre biotopos

Las especies exclusivas por biotopo no se tuvieron en cuenta en este análisis debido a su baja representatividad numérica durante los muestreos. La riqueza de especies, así como los índices de diversidad y uniformidad presentaron valores similares al comparar el río, las lagunas y las quebradas (Tabla 3).
Tabla 3. Riqueza de especies, índices de diversidad (H’) y de uniformidad (J’)

La riqueza de especies calculada por los estimadores Chao2 y Jacknife2 oscilo entre 103 y 111 especies, lo que indica que con la metodología de campo utilizada en este estudio se capturó entre el 71% y 77% de las especies estimadas para el área en esta época climática (Figura 3).
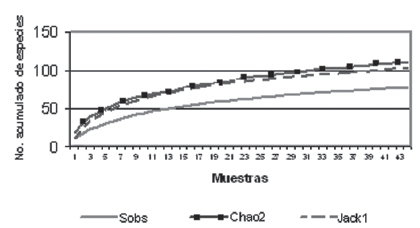
Figura 3. Curva de acumulación de especies observadas y curvas estimadas por Chao2 y Jackknife2 para la zona del río Mesay.
La comunidad íctica del Río Mesay y sus ambientes asociados durante su máxima inundación presenta una elevada riqueza dominada por pocas especies. De un total de 79 especies únicamente para 28 se colectaron más de diez individuos. Sin embargo, estas especies correspondieron a un 97% del total de peces muestreados. La dominancia de especies, expresada como el número de especies que aportan más del 50% de individuos muestreados en un biotopo se manifestó claramente en el río Mesay con dos especies, Brycon falcatus Muller & Troschel 1844 y Triportheus elongatus (Gunther 1864); en las quebradas la dominancia se manifestó con tres especies, Pimelodus blochii Valenciennes 1840, B. falcatus y Leporinus brunneus Myers 1950; y en lagos con cuatro especies, T. elongatus, T. albus (Cope 1872), Pristobrycon calmoni (Steindachner 1908) y Hemiodus unimaculatus (Bloch 1794). La dominancia en los peces de las tallas pequeñas fue mayor, expresada por tan sólo dos especies en los tres biotopos: Hemigrammus gracilis (Lutken 1875) e Hyphessobrycon gracilior Géry 1964 en el río, y H. bellottii (Steindachner 1882), H. gracilis (Lutken 1875) y H. microstomus Durbin 1918, en los otros biotopos. Por otro lado, 25 del total de especies capturadas estuvieron representadas solo por uno o dos individuos.
La agrupación de estaciones según el número de individuos de especies grandes mostró que las estaciones tres y cuatro son exactamente iguales. Este primer grupo se asemeja medianamente a la estación cinco, y muy poco a las estaciones dos y uno (Figura 4). Las especies pequeñas forman dos grupos principales, uno entre las estaciones tres y cinco correspondientes a las quebradas, con una similitud del 100% y otro entre las estaciones dos y cuatro (lagunas) similares aproximadamente en un 60% (Figura 4).
Figura 4. Dendrograma de similitud entre estaciones con base en el número de individuos de especies de tallas grandes (A) y de tallas pequeñas (B), utilizando la distancia euclidiana. (R: río Mesay; Q: quebrada; L: lago).
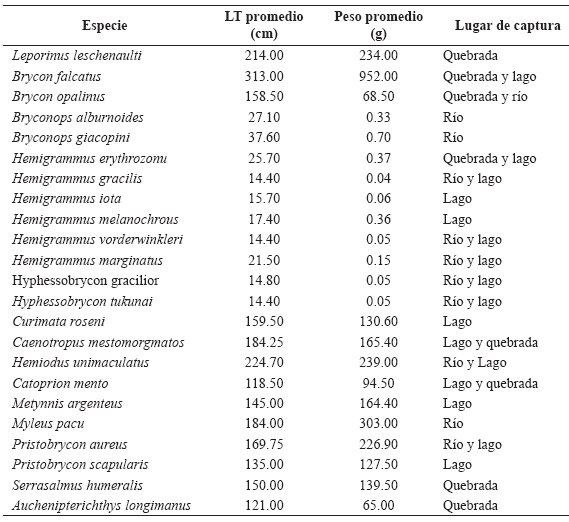
De las 79 especies capturadas en este estudio 23 corresponden a nuevos registros para la región amazónica colombiana (Tabla 4).
Tabla 4. Descripción en talla, peso y lugar de captura de las especies que son nuevos registros para la Amazonia colombiana.
DISCUSIÓN
En los planos inundables de la Amazonia, el pulso hídrico de los ríos es el factor estacional dominante y se ha encontrado que las comunidades ícticas fluctúan enormemente a lo largo del año en respuesta a el (Junk et al. 1989). Por lo tanto, los resultados de este estudio realizado en un solo período específico del pulso hídrico (aguas altas), deben tomarse como información puntual de la comunidad presente en este período en el río Mesay, y no como la generalidad de la comunidad íctica presente a lo largo del año.
La abundancia de especies en los Órdenes taxonómicos siguió el mismo patrón descrito para numerosos ecosistemas dulceacuícolas del neotrópico que se caracterizan por un predominio de peces Characiformes y Siluriformes (Lowe-McConnell 1987), considerados como los principales grupos de peces migratorios de la Amazonia (Goulding et al. 1988). Los Characiformes presentaron el mayor número de especies (63% del total de especies), seguido por los Siluriformes y los Perciformes. La familia Characidae fue la más predominante, posiblemente debido a su importancia numérica dentro del Orden por tener la mayor cantidad de especies (Géry 1984). En los lagos, donde el detrito es probablemente el recurso alimenticio más abundante y la base de la mayoría de cadenas tróficas importantes (Val & de Almeida-Val 1995), se presentó el mayor porcentaje de especies de la familia Curimatidae, conformada en su mayoría por especies detritívoras. La ausencia de peces Gymnotiformes en las muestras denota factores de selectividad inherentes a los métodos de muestreo más que una verdadera ausencia de estos individuos en el área. Sin embargo se desconoce la abundancia de este Orden en el Río Mesay, y se requiere de estudios adicionales para poder concluir a este respecto.
El número de especies encontradas en este estudio es alto a pesar de estar restringido a un único período hidrológico. Aparentemente las ventajas que obtienen los peces al usar hábitats muy variados y diferentes son el factor determinante de la riqueza de especies en esta región (Goulding 1996). Arbeláez (2000) y Prieto (2000) reportan riquezas similares (75 especies) para caños selváticos de aguas negras cerca de Leticia durante la época de aguas altas. Otros estudios, como Knöppel (1970) y Saint-Paul et al. (2000), realizados por 1 y 2 años consecutivos en ecosistemas amazonicos de aguas negras, reportan riquezas de 53 y 172 especies, respectivamente. Aunque estos estudios proporcionan una idea de la riqueza encontrada en otros lugares, las diferencias en la metodología de pesca y en los esfuerzos de captura no permiten una verdadera comparación con los datos de este estudio.
La similitud (CC> 50%) en la composición de las comunidades ícticas presentes en cada biotopo, al igual que el análisis de agrupación entre estaciones evidencian que en esta época no existen patrones de distribución espacial de las especies en los diferentes biotopos. Durante los períodos de aguas altas los peces se dispersan en el plano inundado en busca de alimento y hacen uso de la gran variedad de recursos disponibles. Este comportamiento resulta en importantes migraciones de peces tanto laterales, entre el plano de inundación y el río, como longitudinales, de las cabeceras a las partes bajas de los ríos. De este modo durante la época de inundación todas las especies utilizan en igual medida los ambientes disponibles, ya sea para alimentación, dispersión o reproducción. Aquellas especies exclusivas de un biotopo se vieron representadas únicamente por uno o dos individuos, lo que es más un problema de probabilidades o de limitaciones de las técnicas de pesca que una prueba certera de su presencia o ausencia en el biotopo. El alto número de especies raras o poco frecuentes, la enorme diversidad de habitas disponibles para los peces durante el periodo de inundación y las marcadas variaciones en la composición de la comunidad íctica determinan que la curva de frecuencia acumulada de especies no llegue a estabilizarse por completo y que el valor de riqueza sea inferior al esperado por los estimadores Chao2 y Jackknife2.
Las especies con mayor número de individuos capturados, B. falcatus, T. elongatus, T. albus y P. blochii, pertenecen a géneros de hábitos migratorios que suelen desplazarse en cardúmenes (Goulding et al. 1988), y por tanto, la probabilidad de ser capturados en grandes cantidades es muy alta. En este estudio se observó una posible relación entre el tamaño del plano de inundación y la abundancia de individuos de tallas grandes. Las estaciones que presentaban los planos de inundación mas reducidos (lago dos y quebrada uno), y por tanto una menor oferta de alimento y refugio, se agruparon por poseer un menor número de individuos, mientras el río Mesay que presentaba la mayor área de inundación resultó ser la estación mas disímil de todas por poseer el mayor número de individuos. Las especies pequeñas en cambio se vieron regidas por el carácter lótico/léntico de los ambientes. Las diferencias entre estos dos ambientes se atribuyen a la especiación de hábitat de algunas especies ícticas (Silvano, et al. 2000) y a la influencia que ejerce el flujo del agua en la composición de la comunidad íctica en la Amazonia (Ferreira 1993).
La diversidad de especies encontrada en este estudio puede considerarse baja debido a que sus valores fueron inferiores a 3.0 (Goulding et al. 1988). Esta es una característica de las comunidades ícticas de los ambientes tropicales estacionales, cuyos valores de diversidad suelen ser menores que los de las comunidades que habitan cuerpos de agua no estacionales, debido a la alta dominancia de especies (Lowe-McConnell 1987). El único valor de diversidad por encima de tres (3.1), se presentó para las especies grandes en la quebrada el Sábalo debido a que en esta estación el número de especies dominantes fue menor y la comunidad más uniforme. La diversidad de las especies de peces grandes fue superior a la de las especies pequeñas, posiblemente porque en este último grupo la dominancia estuvo determinada por un menor número de especies. Si se tiene en cuenta el número de especies que representan más de un 50% de la abundancia encontrada en un biotopo, se puede decir que la comunidad íctica del Río Mesay y sus ecosistemas asociados se caracterizó por poseer una alta riqueza dominada por unas pocas especies. Los ambientes estaciónales tienden a tener diversas comunidades ícticas con muy pocas especies dominantes. La dominancia de algunas especies es un proceso dinámico que depende de las condiciones ecológicas del medio, de modo que las variaciones temporales de estas condiciones conllevan a que otras especies se conviertan en dominantes, permitiendo que especies con requerimientos ecológicos similares puedan coexistir (Lowe-McConnell 1967).
En 1999, Mojica mencionó que de las 838 especies de peces dulceacuícolas registradas para Colombia, tan solo 264 lo habían sido para la Amazonia colombiana. Estudios realizados posteriormente en la cuenca del río Amazonas en Leticia: Arbeláez (2000), Prieto (2000), Santos (2000) y Vejarano (2000), aumentaron este numero a 367 especies. Aunque no fue posible determinar la totalidad de los individuos capturados a nivel específico, este estudio contribuye al conocimiento de la biodiversidad amazonica colombiana con 23 nuevos registros de especies. Se anexa una breve descripción de cada uno de ellos en la Tabla 4. La presencia de una alta proporción de especies descritas para el escudo Guayanés en la zona de la Serranía de Chiribiquete podría entenderse como una ictiofauna relictual, aislada del núcleo central del escudo. Por tanto, su estudio desde un enfoque biogeográfico podría aportar elementos de juicio para entender procesos y patrones de distribución de la ictiofauna regional.
AGRADECIMIENTOS
Los autores expresan su agradecimiento a la Fundación Puerto Rastrojo, al Centro de Investigaciones Científicas de la Universidad de Bogotá Jorge Tadeo Lozano y al Instituto de Ciencias Naturales de la Universidad Nacional de Colombia por el apoyo financiero y logístico. Agradecemos muy especialmente a Germán Galvis, Jaime Díaz y Arturo Acero por su asesoría, y a Curupira, Cuñado, Meche y Virgy por su colaboración en el trabajo de campo y la amistad que nos brindaron.
LITERATURA CITADA
1. Arbeláez, F. 2000. Estudio de la ecología de los peces en un caño de aguas negras amazónicas en los alrededores de Leticia (Amazonia colombiana). Tesis Biología Universidad Nacional de Colombia, Bogotá. [ Links ]
2. Colwell, R. K. & J. A. Coddington. 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society (Series B) 345. 101-118. [ Links ]
3. Ferreira, E. J. G. 1993. Composição, distribuição e aspectos ecológicos da ictiofauna de um trecho do rio Trombetas, na área de influência da futura UHE Cachoeira Porteira, Estado do Pará, Brasil. Acta Amazonica, 23:1–89.
4. Gery, J. 1984. The fishes of Amazonia. Págs. 353-370 en: H. Sioli (Ed.), The Amazon: Limnology landscape ecology of a mighty tropical river and its basin. Monographiae Biologicae. Vol. 56.Dr Junk Pub. Dordrecht. [ Links ]
5. Goulding, M., M. Carvallo & E. Ferreira. 1988. Río Negro, rich live in poor water. SBP Academic Publishing, Los Países Bajos. [ Links ]
6. Goulding, M., N.j.h. Smith, & D.j. Mahar. 1996. Floods of fortune. Ecology and Economy along the Amazon. Columbia University Press, Nueva York. [ Links ]
7. Heyer, W. R., J. Coddington, W. J. Kress, P. Acevedo, D. Cole, T. L. Erwin, B. J. Meggers, M. G. Pogue, R. W. Thorington, R. P. Vari, M. J. Weitzman & S. H. Weitzman, 1999. Amazonian biotic data and conservation decisions. Ciência e Cultura Journal of the Brazilian Association for the Advancement of Science. 51. (5/6): 372-385. [ Links ]
8. Hildebrand, P & M. C. Peñuela. 1999. Parque Nacional Natural Chiribiquete. Fundación Puerto Rastrojo, Bogotá. [ Links ]
9. IGAC. 1999. Paisajes fisiográficos de Orinoquia-Amazonia (ORAM) Colombia. Análisis Geográficos Nos. 27-28. Bogotá. [ Links ]
10. Junk, W. J., P. B. Bayley & R. E. Sparles, 1989. The flood pulse concept in river-floodplain systems. Págs. 110-127 en: D. P. Dodge (Ed), Proc. Int. Large River Symp. (LARS). Can Spec Publ Fish Aquat Sci. Vol 106. [ Links ]
11. Knöppel, H. 1970. Food of central Amazonian fishes, contribution to the nutrient ecology of Amazonian rain-foreststream. Amazoniana, 2, (3): 257-352. [ Links ]
12. Lowe-Mcconnell, R.H. 1967. Some factors affecting fish populations in amazonian waters. Atas do simpósio sôbre a biota Amazônica. (Conservacao da naturaleza e recursos naturais). 7: 177-186. [ Links ]
13. Lowe-Mcconnell, R.H. 1987. Ecological studies in tropical fish communities. Cambridge University Press, Gran Bretaña. [ Links ]
14. Mojica-C, J.I. 1999. Lista Preliminar de las especies de peces dulceacuícolas de Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Vol 23, suplemento especial. [ Links ]
15. Nelson, J. 1994. Fishes of the world. A Wiley interscience publication, Nueva York. [ Links ]
16. Prieto, E. 2000. Estudio ictiológico de un caño de aguas negras de la Amazonia colombiana. Tesis de Biología Universidad Nacional de Colombia. Bogotá. [ Links ]
17. Saint-paul, U., J. Zuanon, M. A. Villacorta, M. García, N. Fabré, U. Berger & W. J. Junk. 2000. Fish comunities in central Amazonian white- and blackwater floodplains. Environmental Biology of Fishes. 57: 235-250. [ Links ]
18. Santos, M.s. 2000. Aspectos ecológicos de la fauna íctica dominante en la laguna de Yahuarcaca, Leticia (Amazonia colombiana). Tesis de Biología Universidad Nacional de Colombia. Bogotá. [ Links ]
19. Silvano, R. A. M., B. D. Do Amaral & O. T. Oyakawa. Spatial and temporal patterns of diversity and distribution of the Upper Juruá River fish community (Brazilian Amazon). Enviromental Biology of Fishes. 57.2000. 25-35 p. [ Links ]
20. Val, A. L. & V.m.f. De Almeida-val. 1995. The fishes of the Amazon and their Environment: Physiological and Biochemical Aspect. Springer, Alemania. [ Links ]
21. Vejarano, S. 2000. Ictiofauna de la laguna Yahuarcaca y aspectos tróficos y reproductivos de cinco especies predominantes, Leticia – Colombia. Tesis de Biología Universidad Nacional de Colombia. Bogotá.
Recibido: 19/01/2005
Aceptado: 08/10/2005

