











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
El caucho Hevea brasiliensis (Willd. ex A. Juss.) Müll. Arg. es un árbol originario de la cuenca del Amazonas, del cual se extrae látex natural (Bellacicco et al., 2018; Bottier, 2020). En Colombia se identifican cinco núcleos productivos (Orinoquía, Magdalena Medio, Magdalena Centro, Cordón Cauchero Cacaotero y Amazonía), los cuales están distribuidos a lo largo de la geografía del país (Confederación Cauchera Colombiana (CCC), 2016). No obstante, el 88 % de la producción se concentra en cuatro departamentos (Meta, Caquetá, Santander y Antioquia), entre los que Antioquia participa con el 10% (Castro-Navarro et al., 2020).
En las plantaciones de caucho es importante identificar el comportamiento fenológico, especialmente la dinámica estacional del crecimiento de las hojas, debido a que la defoliación reduce significativamente el índice de área foliar (IAF), lo cual afecta la capacidad de interceptación lumínica, la tasa fotosintética y, en consecuencia, la producción de látex y caucho seco (Leong et al., 1982; Saelim et al., 2019). Además, el árbol requiere de mayores reservas de carbohidratos para la producción de raíces finas y hojas nuevas durante la defoliación (Saelim et al., 2019).
De acuerdo con Gunasekera et al. (2013), el látex corresponde al citoplasma celular, y su biosíntesis sucede en los vasos que se encuentran en los tejidos del floema (Purwaningrum et al., 2019). En este sentido, la producción de látex y el crecimiento del árbol necesitan de los mismos asimilados derivados de la fotosíntesis, por lo que es necesario que se aproveche adecuadamente el látex para mantener el equilibrio entre la producción y el crecimiento (Chantuma et al., 2006). Se sabe que las tecnologías más eficientes para la extracción de látex debilitan el estado fisiológico de los árboles, y que estas deberán adaptarse al metabolismo cada clon y a las condiciones de cada región productora (Adou et al., 2018).
A nivel mundial, la explotación del caucho natural basada en sangría de baja intensidad con aplicación de estimulantes a base de etileno (de Faÿ & Jacob, 2018) ha favorecido ampliamente la disminución en los costos de producción y la mano de obra (Rodrigo et al., 2012; Vijayakumar et al., 2001). Por ejemplo, mediante la aplicación del ácido 2-cloroetil fosfónico (etefón), se retrasa el cierre de los vasos laticíferos, pues esto aumenta la permeabilidad de las membranas, prolonga la salida del látex y, por consiguiente, aumenta la producción (Liu, 2016).
Rivano et al. (2015) mencionan que la estimulación hormonal con etefón permite mejorar la productividad mediante la reducción de la frecuencia de sangría. También aclaran que, para no sobrepasar el potencial de producción de los clones y evitar su colapso fisiológico, es necesario conocer su metabolismo. Esto, debido a que la sobreestimulación puede inducir un estrés oxidativo al interior de las células laticíferas, lo cual conlleva a un desorden fisiológico conocido como panel seco o tapping pannel dryness (TPD), que da como resultado el cese de la producción de látex (Jacob et al., 1994). Lo anterior es causado por la producción de compuestos que deterioran tanto las membranas como la paredes de las células laticíferas, alteración que conduce a un desorden en el sistema laticífero del árbol (Liu, 2016; Putranto et al., 2015), principalmente mediante la formación de especies reactivas de oxígeno (ROS). Por lo tanto, el éxito de la estimulación enfocada al aumento de la producción sin ocasionar problemas de secamiento del panel dependerá de conocimientos relacionados con el material genético, los parámetros fisiológicos del látex, la edad de la plantación, la fenología, el estado nutricional, la frecuencia de sangría, la dosificación y el estado del clima y el tiempo (oportunidad de la aplicación) (Rivano et al., 2015).
Los contenidos de sacarosa, fósforo inorgánico, tioles y sólidos totales (parámetros fisiológicos) intervienen en el flujo de látex y la regeneración celular entre sangrías, lo cual es determinante para su producción. Por medio de la determinación de los parámetros fisiológicos, se han podido establecer las condiciones de sub- o sobreexplotación de las plantaciones de caucho natural, así como la posibilidad de implementar un programa de estimulación (Lacote et al., 2010; Nguyen et al., 2017). Lacote et al. (2010) reportan que la estimulación con etefón es esencial para incrementar la producción de látex tanto en grandes como en pequeños productores. Sin embargo, en Colombia, el estudio de los parámetros fisiológicos que permiten conocer el estado metabólico de los árboles es escaso. Por lo tanto, el objetivo de este estudio fue evaluar distintos parámetros fisiológicos y de producción en árboles de caucho del clon FX3864, los cuales fueron sometidos a distintas frecuencias de sangría con y sin estimulación.
MATERIALES Y MÉTODOS
Ubicación y generalidades de la zona de estudio
El estudio se realizó en una parcela ubicada en el municipio de El Bagre, Antioquia, Colombia. El suelo tenía textura arcillosa y un pH de 4.4. El régimen de lluvias era bimodal, con periodos secos entre diciembre y febrero y entre junio y agosto. La precipitación acumulada media anual fue de 4481 mm, con una temperatura máxima de 29 °C y una mínima de 23 °C (IDEAM), 2021).
Características de la plantación
Las observaciones de campo se llevaron a cabo durante el año 2021 en una plantación de caucho de 12 años, que inició la etapa productiva a los 8 años de establecida y bajo un sistema de explotación S/2 (media espiral), con sangría cada tres días (d3), siete días a la semana (7d/7), doce meses al año (12m/12). El material vegetal correspondió en un 80 % al clon FX3864, corroborado mediante pruebas de identidad genética a través de los cinco microsatélites más polimórficos (SSR). En 2020 se realizaron aportes de correctivos y fertilizantes al suelo, así: cal dolomita (333 kg.ha-1), diamonio fosfato (50 kg.ha-1), urea (100 kg.ha-1), cloruro de potasio (83 kg.ha-1), sulfato de magnesio (33 kg.ha-1) y agrimins (50 kg.ha-1). Las cantidades indicadas se repartieron en un total de 504 árboles (1 ha).
Descripción del estudio
Como factores fijos se establecieron cuatro frecuencias de sangría (FS): d3 (cada tres días, usado convencionalmente en la finca), d4 (cada cuatro días) y d5 (cada cinco días); y la aplicación del ácido 2-cloroetilfosfónico (etefón) al 2.5 % en el 50 % de los árboles, a razón de 1 mL por árbol. Esta mitad recibió estimulación (ET), y la otra permaneció sin aplicación ni estimulación (NET). Las fechas de aplicación correspondieron a los meses de octubre, noviembre y diciembre de 2020, así como marzo, abril y mayo de 2021 (una por mes).
Ontogenia foliar y parámetros fisiológicos
Para definir las épocas de defoliación y renovación foliar, entre noviembre de 2019 y mayo de 2021 se realizó una evaluación de la densidad foliar a partir de una escala de 0 a 10, en donde 0 corresponde al árbol completamente defoliado y 10 corresponde al 100 % de las hojas en el dosel (Cevallos et al., 2012; Rivano et al., 2016). En paralelo, se caracterizó la ontogenia foliar mediante la cuantificación del porcentaje de los estados de desarrollo (A, B, C, D) de las hojas (Lieberei, 2007). El ciclo foliar de cada año se representó con un número consecutivo (Gutiérrez-Vanegas et al., 2020). Así, los estados D1, D2 y D3 correspondían a los ciclos anules de la plantación para los años 2019, 2020, y 2021 respectivamente. Entre julio de 2020 y abril de 2021 se registró la producción de látex en todos los árboles objeto de estudio. Para ello se empleó una probeta graduada y se cuantificó el volumen de látex por árbol. Además, se utilizaron los registros de precipitación media mensual, obtenidos de la base histórica de datos del Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia (IDEAM, 2021).
Para conocer el estado metabólico de los árboles y evaluar el efecto del estimulante en cada FS, se cuantificaron los siguientes parámetros fisiológicos: sacarosa (SAC) en mmol.L-1 como insumo inicial para la biosíntesis de la molécula de caucho, fósforo inorgánico (Pi) en mmol.L-1 como indicador del gasto energético de las células y tioles (RSH) en mmol.L-1 como agente protector de las membranas de las células para disminuir el estrés oxidativo. Además, se midió el contenido de sólidos totales en porcentaje (CST). Los registros se suspendieron durante el período de defoliación/renovación foliar (enero a marzo de 2021). Para evitar síntomas de sobreexplotación que provocaran un deterioro de la plantación, se utilizaron los rangos descritos por D’Auzac y Jacob (1989), en los que se considera que una plantación está sobreexplotada si presenta un contenido de sólidos totales < 30.00%, de sacarosa < 5.00 mmol.L-1, de Pi > 20.00 mmol.L-1 y de RSH > 0.90 o < 0.40 mmol.L-1. Esto, aunado al secamiento de panel.
Para caracterizar los parámetros fisiológicos del látex se realizaron colectas mensuales mediante punción cerca al panel de sangría en diferentes árboles de cada unidad de observación (entre 8 y 12 árboles), en aras de conformar una sola muestra por tratamiento, de acuerdo con la metodología de Sainoi et al. (2017). Para la medición de los parámetros fisiológicos, se emplearon los métodos desarrollados por CIRAD y CNRA, adaptados en 1995 por IRRDB (Jacob et al., 1988). El contenido de SAC se obtuvo mediante el método de Antrona (Ashwell, 1957), y el de Pi a través del de Taussky y Shorr (1953).
Análisis de datos
Para determinar el efecto de la estimulación y la frecuencia de sangría sobre la fisiología del látex, se usó la ecuación: Y ij = μ + τ i + φ j + τ i * φ j + e ij , donde Y ij corresponde a cada una de las variables, μ es la media general, τ i es el efecto de la i-ésima estimulación, φ j es el efecto de la j-ésima frecuencia de sangría, τ i * φ j es la interacción entre niveles de estimulación y frecuencias de sangría, y e ij representa los residuales. A partir de esto, se realizó un análisis de varianza (ANOVA, α = 0.05) de dos vías con la ayuda del paquete agricolae (Mediburu, 2020) del entorno estadístico R (R Core Team, 2020). Cuando se presentó interacción entre los niveles, se analizaron los efectos simples; y cuando no, los efectos principales por el método de Tukey (α = 0.05) para comparar las medias. También se hizo un análisis de componentes principales (ACP) con las variables originales previamente descritas y, posteriormente, se realizó una representación Biplot, lo cual permitió observar la agrupación de los datos con base en las variables originales y el factor discriminatorio de estimulación (paquete ggplot2; Wickham, 2016) del entorno estadístico R (R Core Team, 2020).
RESULTADOS
Ontogenia foliar
Durante los meses de diciembre de 2019 y enero y febrero de 2020 y 2021, se registraron las menores precipitaciones (aproximadamente 100 mm.mes-1), fechas que coinciden con las épocas de defoliación y renovación foliar. No obstante, en el mes de julio de 2020 ocurrió una leve disminución en la precipitación, cercana a 440 mm.mes-1, pero no se generó un estímulo que causara defoliación. Para los años 2020 y 2021, el período de defoliación estuvo concentrado entre los meses de enero y febrero (Figura 1). En febrero de 2020 y marzo de 2021, posterior al periodo de defoliación, se presentó la brotación foliar; se encontraron todos los estados: hojas en estado D del ciclo anterior, ausencia de hojas, y todos los estados ontogénicos de las hojas, desde el estado A (desarrollo del meristemo) hasta el D (maduración total de la hoja). Este proceso tuvo una duración de un mes; en los meses posteriores a la renovación foliar, la densidad foliar estuvo entre 80 y 90 %.
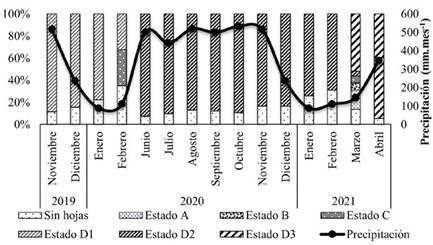
Figura 1 Densidad y ontogenia foliar de H. brasiliensis para los estados de desarrollo A (desarrollo inicial del meristemo foliar), B (hojas rojas inmaduras), C (hojas verde claro inmaduras), D1 (hojas maduras 2019), D2 (hojas maduras 2020) y D3 (hojas maduras 2021); y precipitación media mensual histórica en el municipio de El Bagre, Antioquia.
Parámetros fisiológicos del látex
Para la variable SAC, la interacción entre la frecuencia de sangría y la estimulación resultó significativa (p<0.05) (Figura 2a). Al evaluar los efectos simples, se encontró que las plantas con frecuencia de sangría d3 estimuladas tuvieron un contenido de SAC inferior (6.14 mmol.L-1) en comparación con las plantas en d4 estimuladas (13.26 mmol.L-1), d5 estimuladas (10.57 mmol.L-1), y d5 sin estimular (18.89 mmol.L-1).

Figura 2 Representación gráfica de la interacción entre los factores de estimulación y la frecuencia de sangría. a: sacarosa (SAC) en mmol.L-1; b: tioles (RSH) en mmol.L-1; c: producción (PROD) en mL.árbol-1 día-1. ET: árboles estimulados; NET: árboles sin estimulación.
Respecto a la variable contenido de Pi, no se observó una interacción significativa entre los factores FS y ET (p>0.05). Al evaluar los efectos principales, la estimulación resultó ser el único factor significativo (p<0.05). Así, independientemente de la FS utilizada, las plantas tratadas con etefón al 2.5 % presentaron valores medios de 11.32 mmol.L-1, los cuales fueron significativamente superiores (p<0.05) a la media encontrada en los árboles NET (7.60 mmol.L-1).
Para los RSH, la interacción entre los factores FS y ET resultó significativa (p<0.05) (Figura 1b), razón por la cual se procedió con la evaluación de los efectos simples. El mayor contenido de RSH se registró en la frecuencia d5 con y sin estimulante (0.28 y 0.28 mmol.L-1 respectivamente), superior a lo encontrado en los demás tratamientos (Tabla 1). Para el caso de la frecuencia d4 sin estimular, se obtuvo el menor valor medio, cercano a la mitad con respecto a los demás tratamientos (0.14 mmol.L-1).
Tabla 1 Valores medios de producción y parámetros fisiológicos del látex (sacarosa, fósforo inorgánico, tioles y contenido de sólidos totales) en árboles de caucho natural del clon FX3864
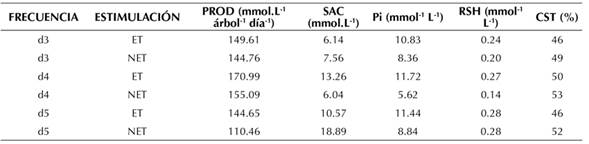
PROD: Producción de látex fresco; SAC: contenido de sacarosa; Pi: contenido de fósforo inorgánico; RSH: contenido de tioles;CST: contenido de sólidos totales; d3: sangrado cada 3 días; d4: sangrado cada 4 días; d5: sangrado cada 5 días; ET: árboles estimulados; NET: árboles sin estimulación.
El CST no presentó diferencias significativas para la interacción de los tratamientos (p>0.05). Al evaluar los efectos principales, la ET y la FS resultaron significativas (p<0.05), lo cual permite resaltar un valor superior en las plantas sin estimular, con un CST del 52 % en comparación con las estimuladas, que alcanzaron una media del 48 %. Sumado a esto, el CST de las frecuencias de sangría d4 (52 %) y d5 (49 %) fueron significativamente diferentes con respecto a d3 (47 %) (p<0.05).
La producción de caucho presentó diferencias significativas para la interacción entre la FS y la ET. Como se observa en la Figura 2c, las plantas estimuladas con etefón y sangradas cada cuatro días (d4) tuvieron una producción superior (170.99 mL.d-1 por árbol) en comparación con las demás frecuencias con o sin estimulación. También se destaca que, para todas las frecuencias de sangría, las plantas estimuladas tuvieron una producción superior cuando se aplicó el etefón, en contraste con aquellas que no recibieron el estimulante.
En la representación Biplot (Figura 3) se observa una separación (con algunos traslapes) entre los individuos estimulados y los no estimulados. Los individuos estimulados tuvieron una mayor producción y contenido de Pi, mientras que, en la mayoría de los casos, los individuos no estimulados tuvieron un mayor contenido SAC y CST. Al agrupar todas las variables, el contenido de RSH fue independiente a la estimulación.
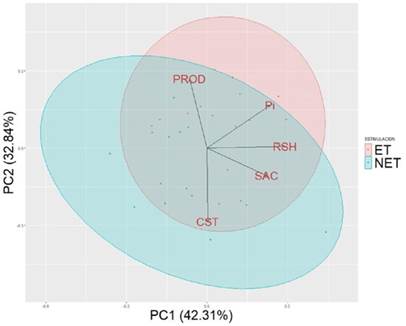
Figura 3 Representación Biplot a partir de un análisis de componentes principales con base en los parámetros fisiológicos del látex y el rendimiento. El factor de discriminación fue la estimulación. La explicación de la variabilidad de la información fue del 75.15 %. PROD: producción de látex fresco; SAC: contenido de sacarosa; Pi: contenido de fósforo inorgánico; RSH: contenido de tioles; CST: contenido de sólidos totales; ET: árboles estimulados; NET: árboles sin estimulación
DISCUSIÓN
Se observó una reducción en la precipitación, la cual, según el trabajo de Guerra-Hincapié et al. (2020) en la zona de estudio, está asociada a un déficit hídrico paralelo a la época de defoliación y renovación foliar en el clon FX3864. En este sentido, Paranjothy (2018) indica que la fenología foliar varía en respuesta a estímulos hídricos deficitarios, lo cual puede ocurrir en diferentes momentos y localidades, especialmente con relación a la ubicación geográfica, que está dada principalmente por la latitud. De acuerdo con los resultados, los procesos de defoliación y renovación foliar se presentaron en los meses de febrero y marzo respectivamente, con intervalos cortos de aproximadamente cuatro semanas cada uno. Se observaron resultados similares en la investigación de Guyot y Le Guen (2018), quienes afirman que este proceso podría tardar hasta 1.5 o 2 meses bajo diferentes condiciones ambientales, tal como lo reportan Gutiérrez-Vanegas et al. (2020) para los clones FX3864 e IAN873 en la Orinoquía colombiana.
Según Righi et al. (2001), en el trópico, los árboles de caucho producen látex durante todo el año. Sin embargo, en épocas de defoliación y renovación foliar, el área fotosintéticamente activa se reduce, disminuyendo así la fotosíntesis y la producción de látex. Conforme lo anterior, durante este período, el aprovechamiento productivo de la plantación agota la cantidad de reservas (carbohidratos), condición que puede alterar la renovación foliar y la calidad de las hojas nuevas que serán utilizadas como fuente de compuestos orgánicos para el crecimiento y mantenimiento de las plantas.
Como lo mencionan Silva et al. (2012), la reducción del área foliar por la defoliación influye directamente sobre la fotosíntesis neta y, por consiguiente, en el contenido de la sacarosa, que provee la energía requerida para la biosíntesis del látex y que además será demandada durante la renovación foliar y la floración. Seneviratne et al. (2020) evaluaron la producción de látex con periodos de descanso de sangría en árboles estimulados, y encontraron que el rendimiento del cultivo se ve significativamente reducido cuando se realiza el sangrado en épocas de defoliación, llegando al máximo en la renovación foliar y floración debido a la utilización de reservas. Además, existe el riesgo de mayor incidencia de TPD, al tener un estrés generado por el consecutivo aprovechamiento de los árboles en esta época.
Como la defoliación comienza en el mes de diciembre, es necesario suspender la estimulación para no generar desbalances metabólicos que conlleven a la utilización de las reservas indispensables para el brote de hojas nuevas. Así mismo, llegado el mes de enero, es necesario suspender el aprovechamiento de los árboles, ya que, en esta época, la defoliación se hace más intensa y los árboles modifican su fisiología para favorecer la renovación foliar. La reanudación del sangrado dependerá de la maduración de las hojas y estará sujeta al estado de desarrollo D. En la zona de estudio, la época de defoliación y renovación foliar se presentó durante los primeros meses del año, resultado que permitirá planear correctamente los tiempos de sangría y de aplicación del estimulante, con el fin de no generar un agotamiento en los árboles, principalmente de las reservas de azúcar.
De acuerdo con lo obtenido en las plantas sangradas en d3 estimuladas (6.14 mmol.L-1) y d4 sin estimular (6.04 mmol.L-1), las cuales representan los menores contenidos, no se alcanzaron niveles considerablemente bajos para el contenido de sacarosa, teniendo en cuenta, que por ejemplo, para esta misma frecuencia de sangría y en plantas estimuladas del clon PB260 en Indonesia, Purwaningrum et al. (2019) encontraron que los contenidos de SAC no superaban los 3.63 mmol.L-1. En contraste, estudios realizados en Brasil con el clon FX3864 bajo la frecuencia de sangría d3 sin estimular reportaron un contenido de SAC de 10.7 mmol.L-1 (Moraes et al., 2011).
Al respecto, Gohet et al. (2019) informan que la aplicación de etefón en la mayoría de los casos resulta en cantidades de SAC bajas, dado su alto consumo para regenerar el látex. De igual manera, Obouayeba et al. (2009) mencionan que los bajos contenidos de SAC podrían deberse a que la aplicación de estimulantes incide sobre el metabolismo de las células especializadas en la producción de látex, en vista de que esta es la materia prima que se sintetiza durante la producción del caucho natural. Conforme lo anterior, y con base en los rangos establecidos por D’Auzac y Jacob (1989), se indica que el resultado de este parámetro fisiológico no evidencia síntomas de sobreexplotación para las condiciones y zona de estudio evaluados, debido a que, bajo ningún tratamiento, el contenido de SAC fue inferior a 5 mmol.L-1.
En cuanto al Pi, la aplicación de etefón produjo un aumento en su contenido, independientemente de la frecuencia utilizada, respuesta análoga a las descripciones realizadas por algunos autores (Atsin et al., 2016; Lacote et al., 2010; Purwaningrum et al., 2019), quienes mencionan que este parámetro es un indicador del incremento de la actividad metabólica cuando se registran concentraciones más altas. Como se observa en la Tabla 1, el contenido de Pi en las plantas ET siempre fue superior, lo cual concuerda con los hallazgos de Atsin et al. (2016) para el clon PB260, quienes encontraron que el contenido medio Pi en el látex estuvo influenciado significativamente por la estimulación, que pasó de los 20.01 mmol.L-1 para los no estimulados hasta los 35.66 mmol.L-1 para los más estimulados, observándose una fuerte tendencia al aumento con la aplicación del regulador de crecimiento. A pesar de que el tratamiento d4 estimulado arrojó el valor más alto (11.72 mmol.L-1), para este parámetro, y bajo las particularidades del estudio, el clon FX3864 en ningún momento experimentó síntomas de sobreestimulación, pues no superó el límite de los 20 mmol.L-1, concentración limite establecida por D’Auzac y Jacob (1989).
D’Auzac y Jacob (1989) indican que valores de tioles por debajo de 0.3 mmol.L-1 podrían representar algún riesgo de sobreexplotación. Adicionalmente, Jacob et al. (1995) exponen que, cuando la intensidad de sangrado es menor, el nivel de tioles es mayor. Con respecto a lo anterior, el mayor contenido de tioles (0.28 mmol.L-1) se registró con la menor intensidad (d5), pero nunca alcanzó los 0.3 mmol.L-1 bajo ningún tratamiento. Coincide lo encontrado en este estudio con en el trabajo realizado por Sainoi et al. (2017), en el que se informa sobre contenidos de tioles de entre 0.17 y 0.24 mmol.L-1 para árboles de 8 años del clon RRIM600 con aplicación de estimulante. Además, Nor Mayati y Mohd (2020) observaron niveles bajos (menores a 1 mmol.L-1) en los clones RRIM2025 y PB350, mientras que Moraes et al. (2011) refieren un contenido medio de tioles de 0.29 mmol.L-1 para el clon FX3864 sin estimular y con una frecuencia de sangría d3. Por lo anterior, la cantidad de tioles obtenida en todos los tratamientos expone la necesidad de profundizar en estudios con respecto a este parámetro, con el fin de generar nuevas categorías para los materiales de origen suramericano plantados en Colombia.
Como lo mencionan Atsin et al. (2016), la sensibilidad de los clones al secamiento del panel está relacionada con bajos niveles de tioles, ya que esto provoca la pérdida de integridad de los lutoides, además de que este parámetro se relaciona con la protección de las membranas, actuando como un compuesto que evita la senescencia. En consecuencia, cabe la posibilidad que se haya generado alguna condición de estrés, principalmente en las plantas con frecuencia d4 sin estimular (0.14 mmol.L-1), probablemente debido a una práctica inadecuada del sangrado. Sin embargo, al observarse que dichos valores fueron inferiores a 0.3 mmol.L-1 (Tabla 1) en todos los tratamientos, se expone la necesidad de realizar un estudio más profundo de los tioles para clones de origen suramericano como el FX3864, posibilitando la construcción de un rango que se ajuste mejor a las condiciones de Colombia, pues los demás parámetros evaluados (SAC, Pi y CST) no arrojaron indicios de sobreexplotación.
En este estudio se encontró que las diferencias en el CST estuvieron ligadas de manera independiente a los tratamientos de estimulación y frecuencia de sangría, lo que permitió que la aplicación de etefón generara una reducción en su contenido por sí sola. Lo mismo ocurrió cuando la frecuencia fue d3. En el primer caso, las diferencias en los contenidos de CST están asociadas al aumento en la producción de látex (Tabla 1), y en el segundo, a lo repetitivo del proceso de sangrado, hecho que probablemente generó una mayor demanda de partículas de caucho durante cada sangría. Al respecto, An et al. (2016) informan que la estimulación con etefón aumenta el rendimiento del látex tan pronto como se realiza su aplicación, el cual generalmente alcanza su máximo un tiempo después (7 días). Los estudios realizados por Attanayake et al. (2018), donde se probaron diferentes concentraciones de etefón, señalan que los valores del CST en tratamientos estimulados disminuyeron significativamente, pasando de un 46 % sin estimulación a un 34 % cuando la concentración del estimulante fue del 5 %. Sin embargo, otros autores no han evidenciado respuestas significativas ante la aplicación del estimulante, tal como ocurrió en el estudio de Sainoi et al. (2017), quienes probaron distintas frecuencias de aplicación de estimulante en plantas de caucho del clon PB261 y obtuvieron CST entre 52 y 55 %. De acuerdo con los rangos establecidos por D’Auzac y Jacob (1989), el límite inferior del CST corresponde al 30 %; en las condiciones de este estudio, el CST nunca disminuyó por debajo del límite aludido, por lo que se descartan posibles síntomas de sobreexplotación con respecto a este parámetro fisiológico.
En términos productivos, los resultados muestran que la frecuencia d4 estimulada presentó la mayor producción (170.99 mL d-1 por árbol), lo cual concuerda con lo mencionado por Rojas M. et al. (2017), quienes hallaron que la frecuencia d4 tuvo una mayor producción en comparación con frecuencias de menor y mayor intensidad. Según los autores, con esa frecuencia se alcanza un equilibrio económico entre el rendimiento y el número de cortes. Aun así, es importante recordar que la producción en el árbol de caucho varía según el genotipo, el sistema de recolección, el uso de estimulantes y la frecuencia de sangría.
CONCLUSIONES
La producción de látex por sangría aumentó cuando los árboles fueron estimulados, y en mayor cantidad cuando la frecuencia de sangría fue d4 (cada cuatro días). Esto sugiere que es posible iniciar esta práctica en el clon FX3864 para las condiciones del Bajo Cauca antioqueño. El análisis de los parámetros fisiológicos del látex (sacarosa, fósforo inorgánico y sólidos totales) y sus registros de producción no arrojaron evidencias de sobreexplotación cuando se aplicó estimulante en las plantas.

