Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Biotecnología
versão impressa ISSN 0123-3475
Rev. colomb. biotecnol v.12 n.1 Bogotá jan./jun. 2010
ARTÍCULO DE INVESTIGACIÓN
Optimization of feeding strategy for the ergosterol production by yeasts Saccharomyces cerevisiae
Optimization of feeding strategy for the ergosterol production by yeasts Saccharomyces cerevisiae
Mojmir Rychtera1 , Josef Cermak2 , Jaroslav Votruba3 , Jan Nahlik4 , Karel Melzoch5 , Christopher A. Kent6 Waldir D. Estela Escalante7
1Professor of biotechnology, Ph.D., Institute of Chemical Technology Prague, Department of Fermentation Chemistry and Bioengineering, Prague, Czech Republic. Mojmir.Rychtera@vscht.cz
2Ph.D. student, Institute of Chemical Technology Prague, Department of Fermentation Chemistry and Bioengineering, Prague, Czech Republic.
3In memoriam, chemical engineer, Ph.D., former address: Institute of Microbiology, Czech Academy of Sciences, Prague, Czech Republic.
4Professor of technical cybernetics, Ph.D., Institute of Chemical Technology Prague, Department of Computer and Control Engineering, Prague, Czech Republic. Jan.Nahlik@vscht.cz
5Professor of biotechnology, Ph.D., Institute of Chemical Technology Prague, Department of Fermentation Chemistry and Bioengineering, Prague, Czech Republic. Karel.Melzoch@vscht.cz
6Reader in Bioengineering, Ph. D., University of Birmingham, School of Chemical Engineering, Birmingham B15 2TT,U.K. kentca@engmail.bham.ac.uk
7Reader in Fermentation Technology, Ph.D., Universidad Nacional Micaela Bastidas de Apurímac, Laboratorio de Biotecnología Agroindustrial, Abancay-Apurímac, Perú. waldirestela@unamba.edu.pe
Recibido: mayo 26 de 2009 Aprobado: junio 9 de 2010
Resumen
Abstract
Objective of this study was to optimize ergosterol production by yeast strain Saccharomyces cerevisiae with the use of computer controlled feeding of cultivation medium. Baker´s yeasts strain of Saccharomyces cerevisiae originally modified and selected as mutant D7 was further applied in an industrial scale and also in this investigation. Composition of cultivation medium was optimized with the use of a modified Rosenbrock´s method with regard to following components: glucose, yeast extract, ammonium sulphate, potassium dihydrogen phosphate, magnesium sulphate and calcium chloride. Cultivation of yeast culture was performed in 7 L laboratory bioreactor with a working volume of 5 L equipped with a control unit and linked to a computer, with dissolved oxygen tension measurement, oxygen and carbon dioxide analyzers. BIOGENES prototype software was created from the commercial control system Genesis for Windows 3.0 (GFW), from Iconics and CLIPS 6.04 for the PC-Windows platform. From various factors affecting sterol biosynthesis a specific growth rate was chosen. Feed rate was controlled according to mathematical model. In this case it dealt with a design of optimal profile of specific growth rate with consequent calculation of carbon dioxide profile. Sterol concentration in the dry biomass increased from 1.0 % up to 3 %.
Key words: Saccharomyces cerevisiae yeasts, ergosterol, fed-batch cultivation control, effect of the specific growth rate.
Resumen
El objetivo de este estudio fue optimizar la producción de ergosterol por una cepa de levadura Saccharomyces cerevisiae, controlando la alimentación de medio de cultivo por computadora. La cepa de levadura panadera Saccharomyces cerevisiae originalmente modificada y seleccionada como mutante D7 fue posteriormente utilizada a escala industrial y también para esta investigación. La composición del medio de cultivo fue optimizada usando el método modificado de Rosenbrock respecto a los siguientes componentes: glucosa, extracto de levadura, sulfato de amonio, fosfato dihidrógeno de potasio, sulfato de magnesio y cloruro de calcio. El cultivo de las células de levadura se llevó a cabo en un biorreactor de laboratorio de 7L con un volumen de trabajo de 5L, equipado con una unidad de control conectada a una computadora, con medición de la tensión de oxígeno disuelto y analizadores de oxígeno y dióxido de carbono. Un software prototipo BIOGENES fue creado a partir del sistema de control comercial Genesis para Windows 3.0 (GFW), de Iconics y CLIPS 6.04 para la plataforma de PC-Windows. A partir de varios factores que afectan la biosíntesis de esterol se escogió una tasa específica de crecimiento. La tasa de alimentación se controló mediante un modelo matemático. En este caso, se trató con un diseño de perfil óptimo de tasa de crecimiento específico con un consecuente cálculo del perfil de dióxido de carbono. La concentración de esterol en la biomasa seca se incrementó desde 1,0% hasta 3%.
Palabras clave: levaduras Saccharomyces cerevisiae, ergosterol, control de cultivo alimentado, efecto de la tasa de crecimiento específico.
Introduction
Sterols form a large group of organic compounds occurring in plants, fungi and animal cells (Parks and Weete, 1991). Their spectrum is quite wide and characteristic of various groups of micro-organisms. Ergosterol -(22E)- Ergosta-5,7,22-trien-3-ß-ol, a precursor of vitamin D2 is an important intracellular product of yeast cells. In human body its main physiological function consists in promoting the body to absorb Ca2+, PO43- and in preventing rickets and osteoporosis. Ergosterol was so named as it was first isolated from a fungus that also produced the ergot alkaloids. Ergosterol is also the main precursor of cortisone and the hormone progesterone and an additive of fodder to increase the laying and hatching rates of fowls. Most of the key enzymes involved in the pathways have been investigated (Chen and Xiao, 1990). Fungi mainly produce ergosterol as the principal sterol accompanied by several C28 sterols.
Answer to the question which of the vitamins (D2 or vitamin D3) is more important in metabolism of Ca and P in human body is not yet accounted for. There is no contemporary evidence showing that vitamin D3 and D2 are equally efficient in metabolic functions. However, few studies have shown important biological differences in this respect between these forms giving preference to vitamin D3 (Trang et al. 1998).
Yeasts are often used as model microorganisms for studies of especially non-membrane functions of sterols and for studying the biochemistry and physiology of sterols. Yeast Saccharomyces cerevisiae is auxotrophic for ergosterol in the absence of oxygen. It was shown that complex changes in esterification of exogenously supplied sterols were also induced by anaerobiosis. Utilization of oleic acid for sterol esterification was significantly impaired in anaerobic cells (Valachovic et al. 2001). Sterol esters formed in high quantity during aerobic growth were decreased when the cells were put under anaerobic conditions and thus accompanied with an increase of free sterols (Taketani et al. 1978). During aerobic growth, the sterols (as esters) decreased from 80 to 45 % in the early exponential phase and then returned to 80 % (as esters) when the culture reached the stationary phase. Under anaerobic conditions, the percentage of sterol esters declined continuously. When growth stopped, only 15 % of the sterols remained esterified. It is possible that the biosynthesis of sterol is not tightly coupled to the cell growth rate (Taylor and Parks, 1978).
Shimizu and Katsuki (1975) proved that anaerobically grown yeasts Saccharomyces cerevisiae at temperature 40 °C when aerated synthesized only 32-35% squalene and sterols comparing with lower temperatures and this decrease was attributed to the repression of the enzymes involved in the synthesis of mevalonate from acetyl-CoA. In addition, at elevated temperature, the metabolic flux from squalene to ergosterol was blocked at squalene epoxidation, lanosterol demethylation, and ergosta-5, 7, 22, 24(28)-tetraene-3beta-ol reduction.
Baker´s yeast belonging to the genus Saccharomyces cerevisiae can under certain cultivation conditions increase the content of intracellularly formed sterols in comparison with other yeast genera. The concentration of yeast sterols changes significantly when extracellular or intracellular conditions are changed in ways that lead to physiological stress (Walker-Caprioglio et al. 1990). He et al. (2000) modified two haploidal yeast species by means of hybridization and obtained two yeast hybrid strains YEH-56 and YEH-28 demonstrating high content of ergosterol in dry biomass. However, they affirmed that it is difficult to get a strain with both higher biomass and higher ergosterol content by natural screening or mutagenesis. The highest content of ergosterol in dry biomass was found in hybrid strain s. cerevisiae YE 193 (7,3 % wt.) while biomass concentration was the lowest (8.5 g/L). Tan et al. (2003) prepared a new strain of s. cerevisiae yeasts by protoplast fusion and with increased internal concentration of ergosterol to 3.07 % dry biomass. He et al. (2003) devoted their interest in conversion of 5,7,22,24(28)-tetraen-3ß-ol (dehydroergosterol) to ergosterol by constructing recombinant expression plasmid carrying ERG4 gene and its overexpression in yeast strain YEH56 using different strong promoters. Ergosterol content in dry cell biomass reached then 4.7 % wt. Interest in industrial production of ergosterol provoked investigators to test ability of newly constructed recombinant yeasts to grow on economically suitable media, e.g. molasses. He et al. (2007) published results obtained with recombinant s. cerevisiae YEH56 (pHXA42) which contains in dry cell biomass around 5.3% wt ergosterol.
Most of the control methods for fedbatch processes already published use information concerning sterols synthesis dependence on the specific growth rate of micro-organisms. According to (Behalova et al., 1994) the highest formation of sterols takes place at the end of the exponential growth phase and in the course of the stationary phase. Production of all sterols in the stationary phase is significantly higher than in the exponential phase just because of the lower growth rate of the cells. In contrast, ergosterol as an important membrane component is formed more in the exponential phase. At the end of this phase the proportion of ergosterol among total sterols is already decreasing (Arnezeder and Hampel, 1990; Cermak et al., 1999; Cermak, 2002). It was also shown that a decrease of specific growth rate from 0.22 h-1 to 0.01 h-1 brought about a twofold increase of ergosterol (Arnezeder and Hampel, 1990).
A comparison of several control strategies for yeast Saccharomyces cerevisiae cultivation based on ethanol, dissolved oxygen, and carbon dioxide concentrations, and on specific growth rate as controlled variables has been published in CHISA´96 and CHISA´98 symposia materials (Rychtera et al., 1996; Rychtera et al., 1998). Tan et al. (2003) studied various fed-batch cultivation configurations (constant feed-rate, constant glucose concentration, exponential feeding, dissolved oxygen exponential fed-batch and dissolved oxygen control fed-batch) and found out that dissolved oxygen control around 12 % of oxygen saturation lead to highest ergosterol yield (around 2.5 % wt in dry cell biomass). Shang et al. (2006) applied in their laboratory experiments control of glucose feeding rate via ethanol monitoring and control. They reached dry yeast concentration 120 g/L and ergosterol concentration in dry biomass 1.25% wt. These authors (Shang et al., 2006a) carried out fed-batch experiments with ethanol control (under 1 % vol.) and under nitrogen limitation. Biomass concentration was a little lower than in previous case ( 95 g/L) but ergosterol content in dry biomass increased to 2.1% wt. Cibis et al. (2001) described in their article behavior of an industrial strain s. cerevisiae possessing higher ability to form ergosterol under continuous and fed-batch experiments. They found out that higher dilution rate lead to unwanted increase of dihydroergosterol while dehydroergosterol concentration in biomass decreased from the maximum attained at dilution rate at 0.07 h-1. A full description of sterol biosynthesis, sterols interchanges and their kinetics in yeasts have not been published so far.
Physiological properties of yeasts Saccharomyces cerevisiae that can be used for cultivation control were well described by Sonnleitner and Käppeli (1986). This yeast species often classified as facultative anaerobic micro-organism possesses a typical character of response to lack of oxygen and to excess of carbohydrate under conditions of optimal aeration. The former phenomenon is known as an alcoholic fermentation and the latter as Crabtree effect (De Deken, 1966). Under condition of Crabtree effect (characterized by a critical glucose concentration) there appears an accumulation of ethanol. This phenomenon was studied and described by many authors since its first disclosure in 1929 (Crabtree, 1929). Keulers (1993) already stressed importance of the specific growth rate for the control strategy of yeast growth but to overcome the problem of this non-measurable variable he suggested a simple observer developed on the basis of on-line measurement. This observer is able to estimate the specific growth rate and the cell concentration. In our paper presented on the CHISA conferences (Rychtera et al., 1996; Rychtera et al., 1996a) and also in a paper (Rychtera et al. 1998) there was described a comparison of several control strategies based on ethanol, dissolved oxygen, carbon dioxide concentrations and on specific growth rate (controlled variables) for the simple growth of yeasts where these parameters were either set constant or variable. Methods trying to increase ergosterol productivity of yeast strains and intracellular concentration of ergosterol in yeasts are based on two different approaches: genetic engineering (Lang et al. 1999), and the microbiological approach exploiting knowledge about the physiology of the yeast strain and its response to changing environmental conditions. A combination of both methodological approaches has the potential for robust operation combined with improved process performance. Most of the methods for control of the baker´s yeast fed-batch process already published are based on controlling the main state values. Parameters such as dissolved oxygen and ethanol concentrations, respiration quotient (RQ) are mostly maintained constant (Keulers, 1993), another variable - carbon dioxide concentration in gas phase must form an increasing function due to increasing biomass concentration. Keulers (1993) studied impact of ethanol formation coefficient on the fermentation activity of the baker´s yeasts in computer-controlled experiments. He suggested that the ethanol production rate coefficient had to be a variable parameter so as to obtain the maximum biomass yield. Consequently, ethanol concentration shows a profile characterized by an increase during the first process phase followed by a gradual decrease until the end of the process preventing the above stated economic loss due to excess of ethanol formed. Similar conclusions with practical control procedure were also claimed by Nahlik (1995). In order to achieve maximum of production parameters a series of different controllers were used. The simplest one is the PID-controller (Axellson, 1989; Cardello and San, 1988). Authors showed that it is difficult to obtain good regulation during the total duration of the cultivation. The results with the PID-controller demonstrate that this technique is limited which is caused by the changing dynamics during the microbial process (Dochain and Perrier, 2000). Numerous techniques utilizing an adaptive controller have been reported trying to compensate the dynamics and non-linearities during the growth of yeasts (Joergensen et al., 1992; Bastin and Dochain, 1990). Keulers (1993) emphasized importance of the specific growth rate for the control strategy but to overcome the problem of this non-measurable variable a simple observer was developed on the basis of on-line measurement. This observer is able to estimate the specific growth rate and the cell concentration. For reaching the maximum yield it is essential to maintain the substrate concentration below its critical level which according to Enfors et al. (1990) is about 11 g/L, according to Wang et al. (1979) 13 g/L, according to Dellweg et al. (1977) 20 g/L. Further, the dissolved oxygen level should be kept above a critical level, which is around 18 % of saturation value. Nevertheless, in a number of industrial fed-batch productions substrate feed rate profile is conventionally controlled on the basis of empirical knowledge, although recently a lot of papers on the application of artificial intelligence techniques such as neural (Linko and Zhu, 1992; Kosola and Linko, 1994), expert (Konstantinov et al., 1993), fuzzy (Park et al., 1993), fuzzy knowledge-based (Shi and Shimizu, 1992), and hybrid neuro-fuzzy systems (Shi and Shimizu, 1992; Schubert, 1994) have been suggested for state estimation, variable prediction and control.
In this paper we present development of optimization of ergosterol biosynthesis in laboratory scale from knowledge-based analysis of industrial production up to a design of fedbatch feeding strategy based on carbon dioxide concentration in exit gas and on specific growth rate (controlled variables). The main intention of our research is to control the specific growth rate, parameter of which is responsible for the activity of the culture and its final quality (ergosterol content). For this reason a deterministic mathematical model was applied and used for simulation of the process.
Material and Methods
Mathematical model
Mathematical model used in this investigation describing growth, substrate consumption, ethanol and other intracellular component production was based on assumptions and equations described in three earlier publications (Rychtera et al., 1998; Behalova et al., 1986; Sobotka et al., 1982). Statistical analysis of experimental results provided a good basis for model construction and optimization. This model consists of mass balances over a perfectly mixed, aerated fed-batch bioreactor and allows the simulation of the time course of concentrations of biomass (X), glucose (S), dissolved oxygen (c), ethanol (E) in the broth, intracellular concentration of sterols, and concentrations of carbon dioxide and oxygen in fermentation gases, as functions of feeding and aeration rates.
Several assumptions allowed us to simplify the original kinetic model and produce the latest version:
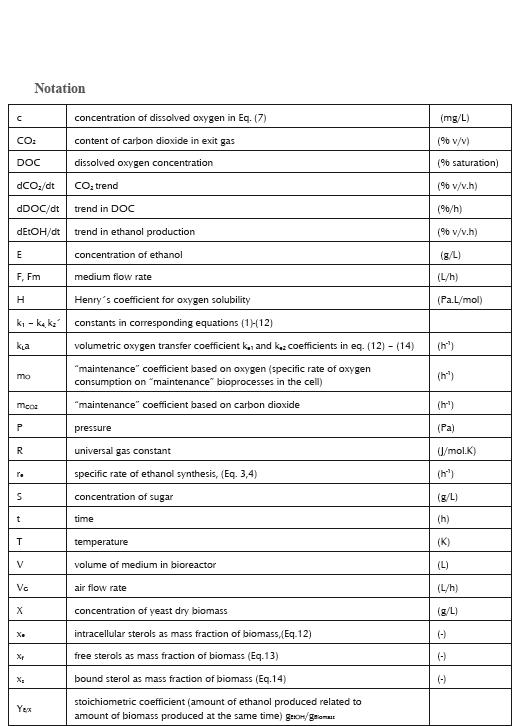
Specific growth rate on sugar (S): µ1 = k1 S. . . . . . . . . . . . . . . . . . . . (1)
Specific growth rate on ethanol (E): µ2 = k2E/(k2´ + E)......................(2)
Specific rate of ethanol formation: re = k3S. . . . . . . . . . . . . . . . . . . (3)
Application of the law of conservation of mass in the form of differential mass balances gives a description of the dynamics of the process, as follows:
Balance on ethanol (E):
dE/dt = re X - YE/X µ2 X - F E /V . . . . . . . . . . . . . . . . . . . . . . . . . (4)
Balance on biomass concentration (X):
dX/dt = µ1 X + µ2 X - F X/V . . . . . . . . . . . . . . . . . . . . . . . . . . . . . (5)
Balance on sugar (S):
dS/dt = -YS/X µ1 X - YS/EreX+ F (S0 - S)/V . . . . . . . . . . . . . . . . . .(6)
Balance on dissolved oxygen concentration (c - mg/L):
dc/dt = kLa ( yO2 P/H - c) - 1000
[YO/X (µ1 + µ2 ) + m O] X . . . . . . . . . . . . . . . . . . . . . . . . . . . . .(7)
Balance on oxygen in the gas phase as mole fraction (yO2):
ε dyO2/dt = P VG/(RT) [0.21 - yO2]/V -kLa (yO2 P/H - c)/32000 . . . . . . . (8)
Balance of carbon dioxide in the gas phase as mole fraction (yCO2):
ε dyCO2/dt = - P VG /(RT) yCO2 /V +[YCO2/X (µ1 + µ2) + m CO2] X/44 . . (9)
Balance of cultivation medium volume in the fermenter (V):
dV/dt = F . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . (10)
The following correlation was found to hold with the bioreactors used in this study, for a stirrer speed of 600 min-1 and an airflow rate of 3-5 L/min, with medium volume in the range of 3.5 – 5 L:
kLa = 32000 P VG (0.21 - yO2)/[(RTV)( yO2 P/H - c)]. . . . . . . . . . . . . . . (11)
Balance on intracellular sterols (xe is expressed as mass fraction of biomass):
dxe /dt = k4S - ke2xe - (µ1 + µ2) xe . . . . . . . . . . . . . . . . . .(12)
Balance of free sterol xF mass fraction dxF /dt = ke1S – ke2xF – (µ1 + µ2) xf. . . . . . . (13)
Balance of sterol bound into cell structures xs mass fraction dxs/dt = ke2xF - (µ1 + µ2) xs . . . . . . . . . . . . . . (14)
(Symbols and units are explained in the notation)
Several laboratory experiments were carried out for the initial model identification. The model was written in PSI/c simulation language. The simulation language PSI/c was used to solve the model equations and also for identification of the model parameters. PSI is an interactive, expression-oriented simulation program for studying the behavior of dynamic and discrete systems, which is considered to be a suitable tool for computer-aided design of process control systems (Bosch van den et al., 1997).
Micro-organism and cultivation medium
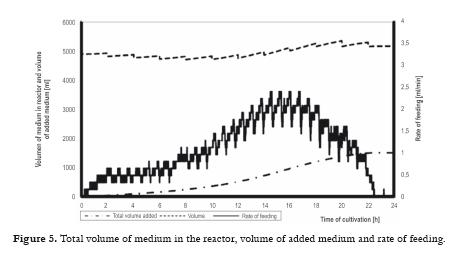
Saccharomyces cerevisiae yeast, strain D7, was provided by a yeast factory. Originally it was modified by UV light and then selected as strain D7 for further applications in an industrial scale. In order to standardize all experimental conditions, an optimized semi-synthetic medium consisting of glucose (125 or 250 g/L), yeast extract (DIFCO) (31.2 g/L), ammonium sulfate (7.8 g/L), potassium dihydrogen phosphate (3.7 g/L), magnesium sulfate (3.1 g/L), calcium chloride (1.25 g/L) in tap water, was used. Medium feed rate, Fm, was used as a manipulated variable, value of which was changed continually, to give required concentration of carbon dioxide in an exit gas.
Modified Rosenbrock´s method for optimization of cultivation medium
Original Rosenbrocks method (Rosenbrock and Storey, 1966) was modified by Votruba et al. (1975) for experimental optimization and as such was applied in this project. Method was modified as a VBA (Visual Basic Application) version of Rosenbrock´s method for multiparameter optimization. The product served as a tool for of computer-aided experimental optimal design of cultivation conditions.
Chemical assays
Biomass determination – 5 ml samples were centrifuged, the sediment washed in distilled water, spun down again, and gravimetrically determined after drying at 105 °C.
Sterols (ergosterol) – Sterols were analyzed using HPLC reverse-phase column, type C18 (250x4mm). Prior to HPLC determination a 3 hour alkaline hydrolysis of cells (3 ml of suspended cell + 3 g KOH) at 100 °C was performed and followed by extraction in diethyl ether, its evaporation and final dissolution in a mobile phase (methanol – water in the ratio 95:5). Evaluation of peak areas was carried out by a standard method.
Glucose and ethanol – These components were determined in the supernatant by HPLC using a column filled with OSTION 0800 in H+ cycle. The mobile phase was sulphuric acid (c = 0.005 mol/L) (Ondrousek and Basarova, 1986).
Bioreactor, monitoring and control
Experiments were carried out in laboratory bioreactor NEW MBR (Switzerland) of the total volume 7 liters (effective volume 5 liters) with control and monitoring system BIOGENES 2.1 which was created in milieu GENESIS 2.1 (ICONICS, USA). Bioreactor was provided with analogue control unit IMCS-2000 (PCS - Switzerland), peristaltic pumps (Verderflex), gas analyzers Hartmann & Braun: URAS 10 (paramagnetic determination of oxygen), MAGNOS 16 (infrared analysis of carbon dioxide). Among other continuously monitoring facilities belongs dissolved oxygen concentration, DOC (measured by polarographic oxygen probe, Mettler Toledo) measurement. The analogue control unit was used to stabilize the environmental conditions: bioreactor temperature, pH of the medium, stirrer speed, air flow, and foam level. BIOGENES® knowledgebased control system (developed by ICT Prague, Dept. of Computing and Control Engineering) running in a Pentium-based computer performed all real-time monitoring and control functions. This system consists of two levels. The basic control level was written on the GENESIS® for Windows 3.5 package (Iconics, USA).
Results and Discussion
Knowledge-based analysis
of an industrial fed-batch
production of sterol yeasts
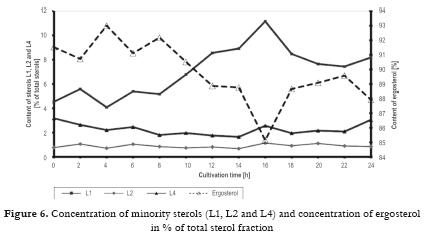
Data for the knowledge analysis of industrial bakers yeast production were collected during a few years in one of the baker´s yeast factory producing yeasts with higher content of ergosterol. These yeasts were further processed in chemical and pharmaceutical plant to obtain pure ergosterol. Purity of this product was the primary indicator for the yeast quality. Industrial cultivation process was carried out in a similar way as in baker´s yeast production (medium from molasses), i.e. by a fed batch method in a 100 m3 bioreactor with a final medium volume of 60-75 m3. The content of sterol fraction of the biomass consists of ergosterol (about 70- 80%) and three other unwanted sterol derivates contaminating the final product denoted (found by HPLC) as L1, L2 and L4 compounds. Symbol L3 in data logs was dedicated to ergosterol. No further studies on their structure were carried out. From literature and metabolic pathways of ergosterol biosynthesis we can presume that sterol L1 is ergosta-5,7,22,24(28)-tetraenol, L4 ergosta-5,7-dienol and L2 can be one of three following sterols: ergosta-7,22,24(28)-trienol, ergosta-5,7,24(28)-trienol or ergosta-5,8,22-trienol. Quality of sterol fraction declared by the ergosterol manufacturer formulates also limits on produced yeasts: content of ergosterol must be higher than 1.1% wt dry weight of yeast biomass, content of L1 in sterol fraction (= 100%) lower than 8%, content of L4 lower than 2.5%.
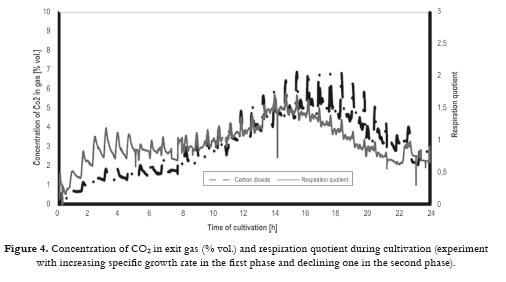
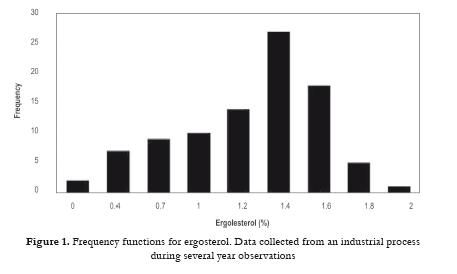
All data were analyzed by basic statistical package which is available in any spreadsheet calculation program. In this case QuatroPro 7 was used. Frequency functions for ergosterol (Fig. 1), L1 and L4 fractions were made. Unfortunately there was not found any strong correlation between ergosterol and L1 or L4 sterol fractions. On the other hand a relation between L1 and L4 fraction has been found. Another analyses using manufacturer data were useful for the development of mathematical model (e.g. course of ethanol concentration during fed batch cultivation, relationship between ergosterol concentration in biomass and ethanol concentration, the time course of specific growth rate). Concentration of ethanol should be controlled in the range 0.12 – 0.14 % vol. The first period (0 - 7 hour) is characterized by maximum growth (µ = 0.2 h-1). In the second period the specific growth rate decreased slowly to 0.02 h-1.
Feeding strategy in cultivation – laboratory experiments
Cultivation experiments carried out in a laboratory bioreactor which are presented in this article were performed mostly twice and only in case of some failure and inappropriate course were repeated for the third time. More experiments of one series were not possible arranged because of length and costs.
In the experiments where mathematical model was used for feeding control, the entire 24 hour cultivation period was divided into two phases differing from each other by time of the first phase and values of the specific growth rate, µ:
a) The first period - lasting 12 – 17 hours - was characterized by a constant value of µ (0.15, 0.125, 0.075 h-1), higher than that of the second phase,
b) The second phase was characterized either by a linearly decreasing value of µ, changing from an initial value to zero at the end of the experiment or by a sudden drop of specific growth rate.
c) In a few experiments was tested also effect of increasing value of specific growth rate in the first phase (0.21, 0.15, 0.1 h-1)
Linearly decreasing value of µ was performed by decreasing flow of the feed medium according to a prespecified profile of CO2. Second phase demonstrated by a sudden decrease of the specific growth rate was carried out by an instant shut-down of the feeding pump.
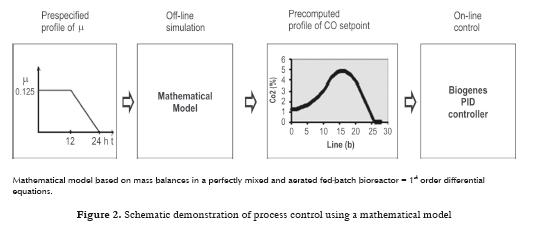
A number of fed-batch experiments were carried out in order to verify mathematical model and to optimize the whole process. In the first 24-hour experiment the cultivation was controlled by a digital PID controller to track a CO2 profile in exit gas calculated from a desired specific growth rate profile using mathematical model equations of which were written in Section 2.1. After one hour start-up phase where the substrate flow rate was kept constant the CO2 control was applied to manipulate the substrate supply. The CO2 (PID) controller closely tracked the CO2 set-point profile divided into 12 linear segments except for the initial three hours when oscillating (because of low biomass concentration) and the final three hours when the process behavior diverged from the simulation results due to limited validity of the mathematical model. The procedure is shown schematically in Fig 2 . Two profiles of the specific growth rate are demonstrated in Fig 3 .
The most likely use of the mathematical model is considered to be both for process simulation and for the development of profiles for real process control in combination with knowledge-based control systems, which would be set up in an industrial plant. The mathematical model and its construction were related closely to an understanding of the physiological character of the biological process and the physico-chemical factors affecting the process. Description of experiments A series of experiments was carried out with control based on the mathematical model. In all cultivations samples were taken every hour for the determination of biomass, glucose and ethanol concentrations in the medium, and for the analyses of sterols and ergosterol in the biomass. From the values of biomass concentration the specific growth rate was calculated. The control system continually monitored the dissolved oxygen tension (DOC), the concentrations of CO2, O2 and ethanol in the exit gas, and the medium feed rate, Fm, and logged these values every minute. From those quantities calculations of the respiration quotient, RQ and the trends dEtOH/dt, dCO2/dt and dDOC/dt were carried out. Evaluation of the experimental results was especially directed to optimization of this process and quality of the produced yeasts (percentage of ergosterol in the total sterols). Effect of specific growth rate profile and extended propagation of yeast culture Specific growth rate as physiological characteristics is not the only one affecting biosynthesis of ergosterol, but it is very important for proportion of various sterols in the sterol fraction. From our studies with the strain D7 the following procedure can be recommended: a) Inoculum must be prepared in at least three stages (adaptation of the culture) where its initial concentration should be around 2 – 3 g/L dry biomass. These stages must be performed step-by-step as a fed-batch process. b) Profile of CO2 should correspond to the course of µ = 0.12 h-1 for the first part of the process (12 h). Figs. 4 5 6 demonstrate a case with significantly improved production parameters. Table 1 summarizes changes both in concentration of total sterols in the biomass (% dry weight) and also in relative concentration of ergosterol in total sterols (%) in a series of experiments. Results of these experiments confirmed a significant role of the specific growth rate on sterol synthesis and on content of individual sterols. Decrease of the specific growth rate has a positive role in sterol synthesis but on the other hand leads to an increase of contaminating sterols, especially 24,28 - dehydroergosterol (L4). In Table 1 there are demonstrated three groups of experiments: a) In the first phase the specific growth rate is increasing, in the second phase the specific growth rate is steadily decreasing (controlled by a feeding regime), b) In the first phase the specific growth rate is constant (relatively high); in the second phase the specific growth rate immediately decreased to a lower value, c) In the first phase the specific growth rate is constant (relatively high); in the second phase the specific growth rate is steadily decreasing to zero value at the end of cultivation. Linearly decreasing value of µ was performed by decreasing flow of the feed medium according to a prespecified profile of CO2. Second phase demonstrated by a sudden decrease of the specific growth rate was carried out by an instant shut-down of the feeding pump (No. 1, 2, 3). Majority of experiments (11, 16, 19, 17, 7, 8) were performed at constant values of the specific growth rate (0.075, 0.125, 0.15 h-1) in the first phase (12 hrs). Another 12 hrs lasted the second phase. Lines in the table are listed stepwise (upwards or downwards) according to measured values of the specific growth rate. Measured values are considered responsible for the cellular changes of the internal content of sterols. Six experiments in the middle part of the Table I show a decreasing trend of the ergosterol content (at the end of cultivation, i.e. 24 h) in the dry cell biomass with increasing value of the measured specific rate in the first phase and linearly decreasing values of this parameter in the second phase. Maximal value of the ergosterol concentration in yeast dry biomass obtained in the industrial process (see Fig 1 ) was 1.4 % (w/w). This series of experiments also shows slightly increasing values of % ergosterol from the fraction of total sterols, it means that purity of produced sterol fraction is higher. This factor is very important for the industrial production of ergosterol. Our results given in the Table I confirm statement of several authors as far as impact of the specific growth rate is concerned. Production of all sterols in stationary phase is significantly higher than in the exponential phase (Behalova et al., 1994). In contrast, ergosterol as an important membrane component is formed more in the exponential phase. At the end of this phase the proportion of ergosterol among total sterols is already decreasing (Arnezeder and Hampel, 1990; Cermak et al., 1999; Cermak, 2002). It was also shown that a decrease of specific growth rate from 0.22 h-1 to 0.01 h-1 brought about a twofold increase of ergosterol (Arnezeder and Hampel, 1990). In our case decrease of the specific growth rate from 0.18 to 0.08 h-1 lead to a threefold increase of ergosterol from 1 to 3% (w/w). It can be concluded that lower specific growth rate (slowly decreasing) in the second phase of cultivation has positive effect on sterol biosynthesis. Lower values of the specific growth rate in the first phase and slowly decreasing values in the second phase lead to higher accumulation of sterols in cells. However, increase of total sterols in the biomass has a negative impact on the fraction of ergosterol in total sterols. Under such control a concentration of total sterols over 3 % can be reached especially when the specific growth rate in the first phase is lower (around 0.08 h-1). Higher growth rate in the first phase mostly leads to lower concentration of total sterols but with higher proportion of ergosterol in the sterol fraction. Another important factor which must be carefully controlled is the concentration of dissolved oxygen which must be kept above its critical value. Conclusions Mathematical model suggested for this bioprocess enables not only simulation of the process but also can be used for control purposes. This method allows to identify conditions under which both a substantial increase in ergosterol concentration can take place and also increasing contamination of ergosterol by other sterols can be avoided. This would have great advantages for downstream processing of yeasts and production of pure ergosterol and vitamin D2. Higher values of the specific rates in the first phase of cultivation (µ > 0.1 h-1) lead to a higher production of biomass with lower content of total sterols but containing higher relative concentration of ergosterol in the sterol fraction (over 90 %). These data are tightly bound to the yeast strain used and can differ from other strains. Mathematical model and its predictive properties can be successfully used in a more complex knowledge-based control system for large scale bioproduction of ergosterol. Acknowledgement This research project was supported by grants MSM6046137305, OK 95088, OK 163 and OK 241 of the Ministry of Education, Youth and Sport of the Czech Republic, and by the European Commission, under the Fourth Framework Programme Copernicus – contract number ERB-CIPA-CT94-0205. The authors highly acknowledge all financial supports. References 1 Arnezeder, C., Hampel, W. A. 1990. Influence of growth rate on the accumulation of ergosterol in yeast cells. Biotechnology Letters 4: 277- 282. [ Links ] 2 Axellson, J. P. 1989. Modelling and Control of Fermentation Processes. Ph.D. Thesis, Lund Institute of Technology, Lund, Sweden. [ Links ] 3 Bastin, G., Dochain, D. 1990. On-line Estimation and Adaptive Control of Bioreactors. Amsterdam, Netherlands: Elsevier. [ Links ] 4 Behalova, B., Blahova, M., Behal, V. 1994. Regulation of sterol biosynthesis in Saccharomyces cerevisiae. Folia Microbiologica 39 (4): 287-290. [ Links ] 5 Behalova, B., Votruba, J., Pichova, A., Beran, K. 1986. Fedbatch cultivation of Saccharomyces cerevisiae with increased content of Δ5,7-sterols. Folia Microbiologica 31 (2): 129-137. [ Links ] 6 Bosch van den, P. P., Butler, H., Soeterboek, A. R., Zaat, M. M. 1997. Modelling and Simulation with PSI. Nuenen, BOZA Automatisierung BV. p. 144. [ Links ] 7 Cardello, R. J., San, K. Y. 1988. The design of controllers for batch bioreactors. Biotechnology Bioengineering 32 (4): 519-526. [ Links ] 8 Cermak, J., Rychtera, M., Nechvile, P., Nahlik, J., Melzoch, K., Sindelar, J. et al. 1999. Influence of the specific growth rate on formation of sterols in yeast Saccharomyces cerevisiae during fed-batch cultivation. Czech Journal of Food Sciences. 18 (3): 11-114 (in Czech). [ Links ] 9 Cermak, J. 2002. Regulation of sterol biosynthesis in yeast cells. PhD Thesis. Institute of Chemical Technology Prague, Prague, Czech Republic. 176 p. [ Links ] 10 Chen, S. Y; Xiao, P. X. 1990. Biochemistry of Yeast Cells. Jinan, Shandong, China, Science and Technology Press. p 5-20. [ Links ] 11 Cibis, E., Garncarek, Z., Miskiewicz, T., Rychtera, M., Borowiak, D. 2001. Polish Journal of Food and Nutrition Sciences.10/51 (3): 29-34. [ Links ] 12 Crabtree, H. G. 1929. Observation on the carbohydrate metabolism of tumors. Biochemical Journal 23: 536- 545. [ Links ] 13 De Deken, R. H. 1966. Crabtree effect: a regulatory system in yeast. Journal of General Microbiology 44 (2): 149-156. [ Links ] 14 Dellweg, H., Bronn, W.K.; Hartmeier, W. 1977. Respiration rates of growing and fermenting yeasts. Kemia- Kemi 4: 611-615. [ Links ] 15 Dochain, D; Perrier, M. 2000. Bioprocess Control. In: Schuegerl, K; Bellgardt, K.-H. (eds.). Bioreaction Engineering. Berlin, Springer-Verlag, p 145-166. [ Links ] 16 Enfors, S.O., Hedenberg, J., Olsson, K. 1990. Simulation of the dynamics in the baker´s yeast process. Bioprocess Engineering 5: 191-198. [ Links ] 17 He, X., Huai, W., Tie, C., Liu, Y., Zhang, B. 2000. Breeding of high ergosterol-producing yeast strains. Journal of Industrial Microbiology & Biotechnology 25: 39-44. [ Links ] 18 He, X., Zhang B., Tan H. 2003. Overexpression of a sterol C-24(28) reductase increases ergosterol production in Saccharomyces cerevisiae. Biotechnology Letters 25: 773-778. [ Links ] 19 He, X., Guo, X., Zhang, B. 2007. Ergosterol production from molasses by genetically modified Saccharomyces cerevisiae. Applied Microbiology and Biotechnology 75 (1): 55-60. [ Links ] 20 Joergensen, S. B., Moeller, H. E., Andersen, M. Y. 1992. Adaptive control of continuous yeast fermentation, near critical dilution rate. IFAC Symposia Series No. 10. Modeling and Control of Biotechnical Processes. 107-112. [ Links ] 21 Keulers, M. 1993. Identification and Control of a Fed-Batch Process: Application to Culture of Saccharomyces cerevisiae. PhD Thesis. Technische Universiteit Eindhoven, The Netherlands. 207 p. [ Links ] 22 Konstantinov, K.B., Yoshida, T., Aarts, R. 1993. Expert systems in bioprocess control: requisite features. Advances in Biochemical Engineering and Biotechnology 48: 192-196. [ Links ] 23 Kosola, A., Linko, P. 1994. Neural control of fed-batch baker´s yeast fermentation. In: Bimbenet, J.J.; Dumoulin, E., Trystram, G. (eds.). Developments in Food Science 36, Automatic Control of Food and Biological Processes. Amsterdam: Elsevier. p. 321-328. [ Links ] 24 Lang, C., Polakowski, T., Stahl, U. 1999. Engineering des Sterol-Metabolismus in Hefe. In: Kreysa, G. and Nick, L. (eds.). Book of Abstracts, DECHEMA-Jahrestagungen ´99, 27.-29. April 1999, Wiesbaden.Rhein-Main- Hallen, Band I. Frakfurt am Main: DECHEMA e.V. p. 145. [ Links ] 25 Linko, P., Zhu, Y. H. 1992. Neural network programming in bioprocess estimation and prediction. In: Karim, M. N., Stephanopoulos, G. (eds.). Modelling and Control of Biotechnical Processes. London: Pergamon Press. p 163-166. [ Links ] 26 Nahlik, J. 1995. Method for the Control of Aerobic Fed-Batch Cultivations of Yeasts. Czech Pat. No. 2773. Appl. No. 2920-2993. [ Links ] 27 Ondrousek, S., Basarová, G. 1986. The use of high-pressure liquid chromatography on ion exchangers for alcohol determination in fermenting media. Sbornik UVTIZ, Potravinarske Vedy (Food Sciences). 4 (2): 91-98 (in Czech). [ Links ] 28 Park, Y. S., Shi, Z. P., Shiba, S., Chantal, C., Iijima, S., Kobayashi, T. 1993. Application of fuzzy reasoning to control glucose and ethanol concentration in baker´s yeast culture. Applied Microbiology and Biotechnology. 38: 649-655. [ Links ] 29 Parks, L.W., Weete, J. D. 1991. Fungal Sterols, In: Patterson G.W. and Nes, W.D. (Eds). Physiology and Biochemistry of Sterols. Champaign, Illinois: American Oil Chemist´s Society. p. 158-171. [ Links ] 30 Rosenbrock, H.H; Storey, C. 1966. Computational Techniques for Chemical Engineers - International Series of Monographs in Chemical Engineering, vol. 7. New York: Pergamon Press, 328 p. [ Links ] 31 Rychtera, M., Votruba, J., Paulova, L., Fiala, J., Melzoch, K., Nahlik J. 1996. Formulation and assessment of control strategies applied to fed-batch yeast cultivation under laboratory conditions. In: Abstracts of the 12th International Congress of Chemical and Process Engineering – CHISA´96, 25-30 August 1996, Prague, Czech Republic. [ Links ] 32 Rychtera, M., Votruba, J., Melzoch, K., Cermak, J., Fiala, J., Vedlichova et al. 1998. Knowledge- based control strategies of sterol biosynthesis in yeast cells. Proceedings of the 13th International Congress of Chemical and Process Engineering – CHISA´98, 23 – 28 August 1998 (CD-ROM of full texts), Prague, Czech Republic. [ Links ] 33 Rychtera, M., Paulova, L., Nahlik, J., Melzoch, K., Votruba, J. 1996a. Control strategy of fed-batch cultivations of yeasts. Chemical papers 50 (4): 238-244 [ Links ] 34 Schubert, J., Simutis, R., Dors, N., Havlik, I., Lubbert, A. 1994. Hybrid modelling of yeast production processes – Combination of a priori knowledge on different levels of sophistication. Chemical Engineering and Technology 17: 10-20. [ Links ] 35 Shang, F., Wen, S., Wang, X., Tan, T. 2006. High-Cell-Density fermentation for ergosterol production by Saccharomyces cerevisiae. Journal of Bioscience and Bioenginering 101 (1): 38-41. [ Links ] 36 Shang, F., Wen, S., Wang, X., Tan, T. 2006a. Effect of nitrogen limitation on the ergosterol production by fed-batch culture of Saccharomyces cerevisiae. Journal of Biotechnology 122: 285-292. [ Links ] 37 Shi, Z., Shimizu, K. 1992. Neuro-fuzzy control of bioreactor systems with pattern recognition. Journal of Fermentation and Bioengineering 74: 39-45. [ Links ] 38 Shimizu, I., Katsuki, H. 1975. Effect of temperature on ergosterol biosynthesis in yeast. Journal of Biochemistry (Tokyo) 77 (5): 1023-1027. [ Links ] 39 Sobotka, M., Votruba, J., Krumphanzl, V., Hradec, 1982. V. Bioengineering analysis of data from commercial bakers yeast production. Kvasny Prumysl 28 (4): 87-89 (in Czech). [ Links ] 40 Sonnleitner, B., Käppelli, O. 1986. Growth of Saccharomyces cerevisiae is controlled by its limited respiratory capacity. Formulation and verification of a hypothesis. Biotechnology Bioengineering 28: 927-937. [ Links ] 41 Taketani, S., Nagai, J., Katsuki, H. 1978. Quantitative aspects of free and esterified sterols in Saccharomyces cerevisiae under various conditions. Biochimica et Biophysica Acta 528 (3): 416-423. [ Links ] 42 Tan, T., Zhang, M., Gao, H. 2003. Ergosterol production by fed-batch fermentation of Saccharomyces cereviaise. Enzyme and Microbial Technology 33: 366-370. [ Links ] 43 Taylor, F. R., Parks, L. W. 1978. Metabolic interconversion of free sterols and steryl esters in Saccharomyces cerevisiae. Journal of Bacteriology 136 (2): 531-537. [ Links ] 44 Trang, H. M., Cole, D. E., Rubin, L. A., Pierratos, A., Siu, S., Vieth, R. 1998. Evidence that vitamin D3 increases serum 25-hydroxyvitamin D more efficiently than does vitamin D3. American Journal of Clinical Nutrition 68: 854-858. [ Links ] 45 Valachovic, M., Hronska, L., Hapala, I. 2001. Anaerobiosis induces complex changes in sterol esterification pattern in the yeast Saccharomyces cerevisiae. FEMS Microbiology Letters 197 (1): 41-45. [ Links ] 46 Votruba, J., Prokop, A., Pilat, P. 1975. Optimization of nutrient medium for culturing microorganisms by means modified Rosenbrocks method. Biotechnology Bioengineering 17 (12): 1833-1837. [ Links ] 47 Walker - Caprioglio, H. M., Casey, W. M., Parks, L. W. 1990. Saccharomyces cerevisiae membrane sterol modifications in response to growth in the presence of ethanol. Applied and Environmental Microbiology 56 (9): 2853-2857. [ Links ] 48 Wang, H. Y., Cooney, C. L., Wang, D. I. C. 1979. Computer control of baker´s yeast production. Biotechnology Bioengineering 21: 975-995. [ Links ]