










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Psiquiatría
Print version ISSN 0034-7450
rev.colomb.psiquiatr. vol.33 suppl.1 Bogotá Oct. 2004
ARTÍCULO DE REVISIÓN
SOBRE LA ORGANIZACIÓN COLUMNAR DE LA CORTEZA CEREBRAL
ON THE COLUMNAR ORGANIZATION OF THE CEREBRAL CORTEZ
Gabriel Arteaga D.*
Hernán José Pimienta J.**
* Psicólogo, Master en Psicobiología State University of New York at Stoony Brook, profesor asistente del Instituto de Psicología de la Universidad del Valle, aspirante a Ph. D. en Ciencias Básicas Médicas: Neurociencias.
** Biólogo, profesor titular de la Universidad del Valle. Director del Centro de Estudios Cerebrales, Facultad de Salud, Universidad del Valle.
Resumen
En el presente trabajo se exponen, de manera sucinta, las dos principales formas de organización estructural de la corteza cerebral de los mamíferos, especialmente de los primates superiores: las denominadas organización horizontal y organización vertical, aunque se hace hincapié en esta última. El patrón de organización vertical se refiere a la disposición en 'láminas' paralelas a la superficie cortical, mientras que el patrón de organización vertical describe agrupamientos celulares, ortogonales a la superficie cortical, los cuales han recibido el nombre de columnas o módulos y han sido propuestos, por diversos autores, como la unidad mínima de procesamiento de la corteza cerebral y base de postulados psicobiológicos contemporáneos.
Se presentan algunos antecedentes relevantes para el concepto de columna cortical, especialmente la documentación experimental obtenida en las cortezas sensoriales, motoras y de asociación. De igual manera, se consideran los componentes celulares, asociados con la organización de las columnas corticales. Finalmente, se describen las características de la conectividad específica, de algunas de las mejor conocidas interneuronas de la corteza cerebral.
Palabras clave: corteza cerebral, módulo, psicobiología, neurona.
Abstract
This paper exposes succinctly the two main tendencies in the structural organization of the mammalian cerebral cortex, particularly that of superior primates: the so called ";horizontal organization" and the ";vertical organization", with an emphasis on the latter. The horizontal pattern of organization refers to the arrangement in ";layers" that run parallel to the cortical surface, whereas the vertical pattern describes clusters of cells orthogonal to the cerebral surface called ";columns" or ";modules", those arrangements have been proposed as the ";minimal processing unit" in the cerebral cortex and constitute the basis of contemporary psychobiological postulates.
Relevant background to the concept of column is presented, particularly the experimental data found in the sensorial, motor and associative cortex. Cellular components associated with the structure of the cortical columns are considered. Finally the specific connectivity of some of the best known interneurons of the cerebral cortex is described.
Key words: Cerebral cortex, module, psychobiology, neurone.
Introducción
Las primeras descripciones de la estructura de la corteza cerebral se realizaron durante la segunda mitad del siglo XIX, asociadas con el desarrollo de las técnicas de tinción celular. Hanover (1840) utilizó por primera vez la fijación del tejido nervioso en ácido crómico; asimismo, Von Gerlach (1858) introdujo la tinción celular por medio de dicromato de potasio y carmín amoniacal (1). Gracias a estos procedimientos fue posible describir la corteza cerebral como una estructura compleja, compuesta por diversos tipos de células, y eventualmente, a finales del siglo XIX y principios XX, el uso de estas metodologías permitió tres avances conceptuales que fundamentan el desarrollo de la moderna neurobiología de la corteza cerebral.
La primera de estas concepciones corresponde a la formulación del modelo de organización laminar de la corteza cerebral, particularmente la propuesta formulada por Meynert (1867-1872) y por Lewis y Clarke (1878). Estas propuestas establecieron, en general, que la corteza cerebral de los mamíferos, incluida la humana, está constituida por seis 'láminas' o estratos, emplazados en sentido paralelo a la superficie pial y que son diferenciables en función del tipo y el tamaño de las células que los componen (1),(2).
En segundo lugar, como una consecuencia de la formulación precedente se desarrollan los llamados mapas citoarquitectónicos, los cuales fraccionan la superficie de la corteza cerebral en cierto número de áreas, en función de presuntas variaciones sistemáticas en la composición celular, en toda la topografía cortical. De los diversos mapas propuestos, sin duda, el más conocido es el de Brodman (1906, 1909 y 1912), el cual adopta el modelo de las seis láminas e introduce una nomenclatura numérica todavía ampliamente utilizada, para la denominación de 52 áreas en la corteza cerebral humana (3),(4),(5).
Finalmente, hacia finales del siglo XIX, el neuroanatomista español Santiago Ramón y Cajal (1900) desarrolla la denominada teoría neuronal, ya propuesta por Waldeyer (1886), en la cual se establece que la neurona constituye la unidad estructural, funcional y trófica del sistema nervioso. A la vez, esta teoría caracteriza las neuronas como elementos celulares discretos, separadas por el espacio sináptico. Aunque los términos neurona y sinapsis fueron acuñados por Waldeyer y por Sherrington, respectivamente, fue sin duda el extenso trabajo de observación y descripción de la composición celular del tejido cerebral, desarrollado por Ramón y Cajal, lo que universalizó la ";doctrina neuronal del sistema nervioso" y le valió a su autor el premio Nobel de Fisiología en 1908 (6),(7).
Desde otra perspectiva, Marín-Padilla (1998) considera que el desarrollo contemporáneo de las concepciones sobre la estructura de la corteza cerebral se organiza en torno a dos escuelas que se caracterizan por la utilización de métodos diferentes de preparación y tinción del tejido nervioso. Por una parte, la Escuela Alemana, que utiliza métodos que permiten apreciar preferencialmente los cuerpos celulares o somas de las neuronas con la utilización de violeta de Cresyl o tinción de Nissl, la cual reacciona con los corpúsculos de este nombre localizados en el citoplasma de las neuronas y permite apreciar la organización de láminas horizontales. Este procedimiento condujo a esta escuela a las descripciones y mapas citoarquitectónicos, como el ya mencionado de Brodman (8),(9). Por otra parte, la Escuela Española de Citoarquitectura se enraíza en el trabajo monumental de Ramón y Cajal y sus discípulos, el cual a su vez se basa en las tinciones con dicromato de potasio y nitrato de plata, desarrolladas por el fisiólogo italiano Camilo Golgi (1886), quien por esta razón compartió la distinción del Nobel con Ramón y Cajal.
A diferencia de los métodos que permiten visualizar los somas neuronales, en el procedimiento de Golgi, si bien sólo reaccionan aleatoriamente una de cada cien neuronas, es posible a cambio la impregnación de todas las proyecciones dendríticas y axonales de cada una de las neuronas reactivas. De esta manera se hace evidente, en primer lugar, que las células nerviosas poseen una muy considerable profusión de procesos de interconexión (neuropilo) y, en segundo lugar, que éstos se extienden dentro del plano vertical, especialmente en el caso de las neuronas piramidales, las cuales, como lo señala Fedelman (1984), tienden a tener una marcada polarización en el sentido vertical de la corteza cerebral.
La descripción de columnas en la corteza cerebral
Desde el punto de vista experimental, la descripción de columnas en la corteza cerebral se ha realizado principalmente utilizando diversas especies de mamíferos, incluidos humanos; pero, sobre todo, la descripción de estas estructuras se ha desarrollado en una suerte de progresión desde las cortezas sensoriales, donde fueron originalmente identificadas, hasta las cortezas de asociación y la corteza motora primaria.
Además de la obra de Ramón y Cajal, la primera referencia a unidades de organización vertical de la corteza cerebral la hace Llorente de Nó (1938 y 1949), quien describe 'cilindros' funcionalmente dispuestos en torno a elementos tanto aferentes como eferentes, caracterizados por un patrón de conexiones internas. A estos cilindros el autor los considera como unidades "elementales de procesamiento" de la corteza cerebral. El término columna, sin embargo, fue utilizado originalmente por Von Economo y Koskinas (1925) en la descripción de la organización histológica del giro temporal superior de sujetos humanos, donde la presencia de módulos verticales parece muy obvia, especialmente cuando se utilizan tinciones de Nissl [citados por Buehoeveden y Casanova (5) y Mountcastle (10)].
Por otra parte, la propuesta formal de una organización en columnas de la corteza cerebral, lo que ha sido llamado la hipótesis columnar, fue formulada por Mountcastle [1957, 1959 y 1973, citado por Mountcastle (10),(11)], basado en sus trabajos experimentales en que utilizó la penetración de electrodos en sentido perpendicular a la superficie de la corteza somatosensorial de gatos y monos. Este procedimiento les permitió a los investigadores identificar series verticales de neuronas a lo largo del recorrido del electrodo, las cuales responden al mismo estímulo. De esta manera, describieron agrupamientos de células que forman columnas verticales, las cuales procesan en conjunto información procedente de la misma fuente de estímulo, es decir, poseen el mismo campo receptivo.
Una nueva y significativa validación experimental de la hipótesis columnar se produjo durante los años sesenta del siglo pasado, con el galardonado trabajo de Hubel y Wiesel (1980). Estos investigadores, utilizando la misma técnica de electrodos penetrantes ortogonales a la superficie cortical, pero esta vez emplazados en la corteza visual primaria de monos rhesus, describieron grupos celulares que los autores caracterizaron como columnas de dominancia ocular y columnas de orientación, definidas por las proyecciones aferentes procedentes del núcleo geniculado lateral del tálamo.
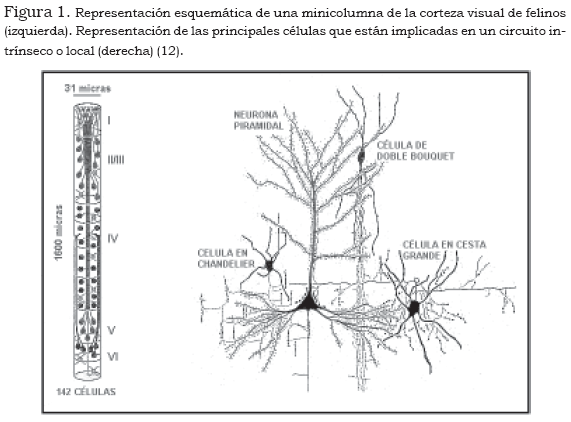
Estas estructuras están organizadas sobre el mismo principio de correspondencia de la organización vertical, bien sea con áreas delimitadas del campo receptivo o con características específicas de los estímulos procesados, como la forma o el color (13), (14), (15). De igual manera, se ha descrito la existencia de patrones organizativos modulares, más allá de la corteza estriada. Britten (1998), utilizando registro mediante electrodos penetrantes, señala que en la vertiente dorsal del sistema visual de macacos, correspondiente al área temporal medial, se encuentran agregados de células con patrones de respuesta similar.
Otro hito significativo en la consolidación de la hipótesis columnar en el ámbito de las cortezas primarias sensoriales lo constituyen los trabajos de Woolsey y Van der Loose (1970) y de Welker (1976), quienes describieron una estructura específica localizada en la lámina IV de la corteza somatosensorial de roedores, a la cual los investigadores denominaron barrels. Estas estructuras están asociadas con la percepción compleja del espacio y presentan una correspondencia punto a punto con los 'bigotes' (whiskers) del lado contralateral del hocico del animal, es decir, nuevamente, conforman un esquema de correspondencia receptotópica con regiones específicas del campo receptivo y áreas discretas de la corteza cerebral (16),(17).
La descripción de la organización columnar en las cortezas auditivas remite a la identificación de la proyección tonotópica en la corteza auditiva primaria de mamíferos. De acuerdo con Levine y Sheffner (18), la membrana basilar, localizada en el interior de la cóclea es un ";analizador de frecuencia", de tal manera que áreas específicas de su superficie responden preferencialmente dentro de ciertos rangos de frecuencia. Estos rangos, a su vez, están representados de manera sistemática en la superficie de la corteza auditiva primaria y conforman curvas de isofrecuencia en sentido anteroposterior (18). Por otra parte, en dirección ortogonal al vector anteriormente descrito, se organiza otro mapa en el sentido dorsoventral, el cual codifica la amplitud de las ondas sonoras.
De esta manera, por lo menos dos componentes de los estímulos auditivos (frecuencia y amplitud) están representados de manera sistemática en las cortezas auditivas primarias; pero, de igual manera, otras características de respuesta auditiva se encuentran segregadas en estas regiones corticales (19), (20). Por otra parte, la utilización de microelectrodos penetrantes en corteza auditiva primaria revela la presencia de columnas de células con preferencias de respuesta similares, tanto en frecuencia como en amplitud de las ondas sonoras (18). Igualmente, estudios con trazadores retrógrados sugieren una organización modular para el procesamiento de diversas propiedades de los estímulos auditivos, en la corteza auditiva primaria de mamíferos (20).
Una descripción anatómica de la organización modular en diversas regiones de la corteza cerebral de roedores fue realizada por Escobar et al. (21), quienes utilizaron la marcación de la proteína MAP-2, mediante el anticuerpo monoclonar 5f9, para estudiar las fibras del citoesqueleto de neuronas piramidales. Los autores reportan un patrón de organización de tipo 'panal' para las arborizaciones terminales de dendritas apicales de neuronas piramidales, con variaciones en la forma de las columnas, dependiente de la región cortical considerada.
En la corteza motora la descripción de una organización columnar ha sido menos evidente, quizá porque se trata del sistema de salida (eferente), el cual supone la síntesis de información, que depende de las áreas sensoriales, particularmente de la corteza parietal posterior. No obstante, Lev y White (22) han descrito una organización de tipo columnas en la corteza motora primaria de ratones. Estos investigadores, utilizando la marcación de las fibras del citoesqueleto, mediante el anticuerpo contra la proteína MAP-2, marcaron las dendritas de neuronas piramidales y encontraron un patrón de organización modular similar al descrito en las cortezas sensoriales.
Por su parte, Meyer [1983, citado por Mountcastle (11)] describe en la lámina V de la corteza motora primaria, de primates superiores, grupos de neuronas piramidales distribuidos intermitentemente en el plano horizontal. El investigador describe que tanto las neuronas piramidales como las no piramidales se organizan en agregados tipo columna, de unas 300 µm de diámetro, separadas por espacios de aproximadamente 100 µm, los cuales proyectan eferentes a grupos individuales de neuronas motoras de la médula espinal. Por otra parte, Keller (23) propone que en los primates la corteza motora está compuesta por agregados columnares o módulos de neuronas asociados con los diferentes aspectos de un movimiento particular. Adicionalmente, estas columnas suponen patrones internos de conexiones excitatorias e inhibitorias, cuya modificación tiene lugar en función de la experiencia, es decir, de la plasticidad cortical.
Rizzolatti y Luppino (24) señalan que la corteza motora primaria está constituida por un 'mosaico' de áreas anatómica y funcionalmente segregadas, conectadas profusa y recíprocamente con la corteza parietal posterior, las cuales constituyen las unidades básicas del sistema motor, cuyo papel general es transformar la información sensorial en acción.
En una dirección similar, Sanes y Donoghue (25) subrayan que los hallazgos empíricos de diversas fuentes apuntan a una consideración según la cual la organización somatotópica de la corteza motora primaria es aplicable tan sólo a regiones corporales relativamente grandes, como brazos, piernas, cara, etc. Internamente, cada una de esas divisiones estaría compuesta por redes distribuidas de conexiones, que reclutan poblaciones amplias de neuronas para la ejecución de patrones complejos de actividad.
Finalmente, en lo que concierne a la organización columnar de la corteza cerebral en áreas de asociación, se considera a continuación una muestra mínima, pero de alguna manera representativa, de los diversos trabajos de investigación sobre el tema en cuestión, realizados en primates superiores, tanto en lo que corresponde a las estrategias metodológicas como a los problemas y resultados que éstos han permitido esclarecer.
Quizá la primera descripción anatómica de módulos o columnas en áreas de asociación cortical se encuentra en la aproximación morfológica inaugurada por el trabajo de Goldman y Nauta (26), en el cual se realiza marcación retrógrada mediante [3H]leucina y [3H]prolina, en tres áreas de la corteza frontal de monos rhesus: corteza motora primaria, giro orbital medial y región prefrontal dorsolateral, comprendida en el labio superior del surco principal. En estas áreas los investigadores describieron columnas verticalmente orientadas, las cuales se extendían en todas las láminas corticales. Estas estructuras poseen diámetros entre 200 y 500 µm, interdigitadas con regiones de diámetro equivalente, pero pobres en composición celular.
En un estudio similar, Bugbee y Goldman-Rakic (27) utilizaron la técnica de trazado retrógrado para caracterizar la organización columnar de la corteza cerebral en dos especies de primates del Viejo y del Nuevo Mundo, especialmente en las áreas de asociación prefrontal. Los resultados mostraron que el tamaño de las columnas corticales es consistentemente similar entre las dos especies estudiadas, a pesar de diferencias considerables en el tamaño global del cerebro. Los investigadores sugieren, a partir de los resultados, que la tendencia evolutiva de la corteza cerebral probablemente se orientó hacia el aumento del número de las columnas, más que hacia el aumento del espesor cortical
Levitt et al. (28) estudiaron los patrones de conectividad ínter e intralaminar de las neuronas piramidales localizadas en la corteza prefrontal de macacos, correspondiente a las áreas 9 y 46 de la nomenclatura de Walker (1948). Con este propósito, los investigadores realizaron microinyecciones de biocitina para trazar las proyecciones ortógradas de las neuronas piramidales, de las láminas superficiales de la corteza (supragranulares) en estudio.
Las inyecciones produjeron un patrón de marcación lateral dentro de la misma área, en una superficie de 200-400 ìm de diámetro y 2,5 mm de longitud, centradas en el sitio de la inyección. De igual manera, desde este mismo sitio se describen fibras colaterales que forman bandas marcadas, interpuestas entre bandas sin marcar. De acuerdo con los investigadores, este patrón se forma a partir de las proyecciones que vienen de neuronas piramidales, localizadas en las láminas II, III y V, las cuales terminan en las láminas superficiales I-III (28).
Kritzer y Goldman-Rakic (29) investigaron, igualmente, la circuitería intrínseca de la corteza prefrontal de monos rhesus, pero utilizando marcación retrógrada con toxina de cólera (B). Los autores describen diferentes tipos de patrones ascendentes y descendentes, así como circuitos locales intra e interlaminares, que en general son compatibles con la organización en columnas verticales, la cual en conjunto constituye el patrón de bandas descrito en otros estudios. Los investigadores hacen notar que los circuitos que se encuentran en estas áreas corticales se diferencian de otros, como los descritos en cortezas sensoriales y que estas diferencias se corresponden seguramente con diferencias funcionales.
Finalmente, en un trabajo en el cual se combinan las técnicas de marcación mediante transporte anterógrado y retrógrado, Pücak et al. (30) se proponen la ";reconstrucción espacial" de los patrones de proyección de las neuronas piramidales, tanto a través de los axones principales como de los colaterales. Las neuronas de interés se encontraban localizadas en la lámina III, en la corteza prefrontal (áreas 9 y 46), de monos rhesus.
Los autores describen agregados intrínsecos (intrinsic clusters) dispuestos en los ya mencionados patrones de bandas, por marcación de colaterales axónicos, los cuales presentan conectividad recíproca, como lo evidencia la doble marcación. Por otra parte, el transporte a través de axones principales formó agregados asociativos (associational clusters) igualmente dispuestos en forma de bandas marcadas, alternadas con bandas no marcadas (30).
Los resultados permitieron mostrar que el tamaño de las bandas marcadas anterógrada o retrógradamente tiende a ser igual; de la misma manera, las bandas de proyección, formadas por los axones principales, tienden a tener el mismo tamaño que las bandas intrínsecas, formadas por colaterales axónicos (aproximadamente 300 µm). Los investigadores están de acuerdo en considerar que estas estructuras, caracterizadas por convergencia, divergencia y reciprocidad en su conectividad local, constituirían el sustrato anatómico básico de los procesos complejos ejecutados por estas áreas corticales.
Concepto de módulo cortical
No solamente la descripción sino sobre todo la conceptualización del modelo de columnas o módulos corticales recibió un impulso definitivo a partir del trabajo del neurobiólogo húngaro Janos Szentagothai (1969, 1975 y 1983). Este investigador, basándose en los trabajos fisiológicos de las décadas de los cincuenta y de los sesenta del siglo pasado, especialmente los de Mountcastle, en corteza somatosensorial de gatos, y los de Hubel y Wiesel, en corteza visual de primates, introdujo el concepto de principio de arquitectura modular, para establecer un sustrato anatómico a estos fenómenos que habían sido descritos fisiológicamente (31).
En particular, Szentagothai propone un modelo tridimensional en el cual los módulos o columnas, tanto en las cortezas sensoriales como en las de asociación, estarían definidos a partir de tres componentes básicos. En primer lugar, por las proyecciones aferentes hacia el módulo, así como por las proyecciones eferentes que del módulo se dirigen a otras regiones de la corteza cerebral. En segundo lugar, por los componentes neuronales del módulo, considerados funcional y morfológicamente como pertenecientes a dos grandes categorías: neuronas excitatorias e inhibitorias. En tercer lugar, por el módulo cortical, que puede ser descrito en términos de su circuitería interna —vale decir el patrón de conectividad intrínseco que se establece entre los componentes celulares, mencionados anteriormente— (31).
Szentagothai señala que en la corteza de asociación de los mamíferos un módulo, cuyo diámetro es de 300 µm en promedio, estaría compuesto por aproximadamente cuatro mil neuronas, de las cuales, en términos generales, el 70%-80% serían neuronas piramidales, es decir, neuronas de proyección. Los axones de estas células pueden dirigirse a otras áreas de la corteza cerebral del mismo hemisferio o pueden cursar a través del cuerpo calloso hacia regiones homólogas 'en espejo', de la corteza del hemisferio contralateral. El autor estima que cada módulo podría recibir y a su vez proyectar hacia por lo menos otros cincuenta módulos respectivamente. Finalmente, considera que en la neocorteza cerebral humana existirían al menos tres millones de estas estructuras.
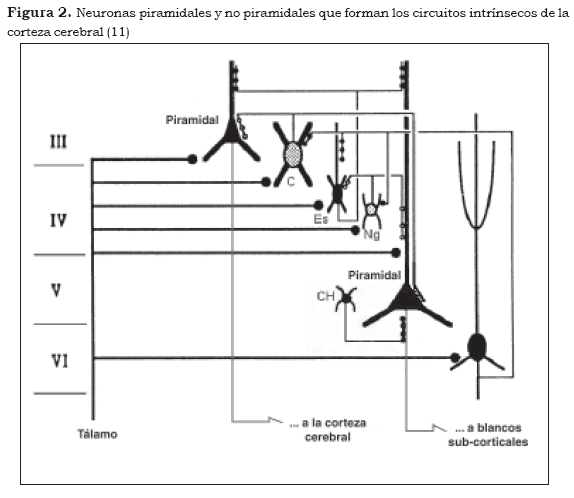
En la figura anterior se representan tanto los aferentes talámicos como dos de los sistemas de eferentes desde la corteza: córtico-cortical y córtico-subcortical. Por otra parte, Eccles (31) se ha aproximado al concepto de modularidad de la corteza cerebral, desde el punto de vista de su contribución a la operatividad de la misma. Considera que los módulos son los ";… elementos anatómicos básicos en el diseño funcional del córtex". En esta dirección, el autor señala que la función de la corteza cerebral depende críticamente de los patrones de conectividad de los módulos corticales; no obstante, tales patrones se refieren no solamente a la dimensión espacial, sino también a la temporal, en la medida en que la conectividad variaría ";momento a momento", en función del flujo de la información aferente (31).
En el contexto del problema mentecerebro, Eccles (32) hace observar que un correlato anatómico de la operación modular de toda la corteza cerebral lo constituye la organización en agregados discretos (bundles) de las dendritas apicales, de las neuronas piramidales, las cuales tienen un curso intracortical ascendente hacia la lámina I. El autor denomina a estos agrupamientos dendrones y los considera microcircuitos estructurales de la corteza cerebral. Estas estructuras, postula, constituirían el punto de interacción entre los eventos neurales y procesos mentales unitarios, a los cuales por contrapartida denomina psicones. Dicha interacción sería análoga a la que se ha propuesto para los campos de probabilidad de la mecánica cuántica, los cuales pueden ejercer efectos en micrositios, a pesar de carecer de energía y de masa (33).
Un dendrón estaría constituido por aproximadamente cien dendritas apicales, de igual número de células piramidales, localizadas en diferentes láminas corticales; la mitad provendría de piramidales de la lámina V, mientras que el resto se reclutaría de piramidales localizadas en las láminas II y III. El diámetro de esta estructura sería de 50 ìm en promedio, lo cual representa sólo una fracción del diámetro atribuido a las columnas corticales, en trabajos experimentales, previamente mencionados.
Al menos desde el punto de vista de sus dimensiones, como se verá más adelante, la estructura propuesta por Eccles parece más compatible con el concepto de mini o microcolumna, descrita por otros autores. Finalmente, el estimado total de estas unidades en la corteza cerebral humana es de cuarenta millones, cifra que ampliamente podría dar cuenta de toda la riqueza de la experiencia mental humana (31), (32).
Mountcastle (10),(11), como ya se ha señalado, fue el pionero en la descripción de los módulos corticales en las áreas sensoriales primarias, por medio del uso de métodos fisiológicos. Pero, además, a este autor se debe la formalización de la teoría de la organización columnar de la corteza cerebral, de acuerdo con la cual la ";unidad básica de la operación cortical es la mini-columna", una estructura de orientación vertical, constituida por grupos de entre ochenta y cien neuronas, distribuidas a lo largo de todas las láminas corticales, excepto la primera por su carencia relativa de células (10),(11).
Cada minicolumna contendría todos los tipos de células de la corteza, en proporciones diferentes, dependiendo del área y de la función. Desde el punto de vista funcional, estas estructuras constituyen ";unidades de procesamiento complejo", cuya función se describe como una operación de transformación de los imputs en outputs, mediante cadenas de procesamiento intrínseco. Por otra parte, las conexiones intracorticales largas pondrían en relación columnas con propiedades similares en el terreno funcional.
Por su parte, las columnas estarían conformadas por grupos de minicolumnas unidas por conexiones horizontales cortas, que comparten el mismo imput, además de un cierto número de propiedades fisiológicas. El número de minicolumnas en el interior de una columna podría ser variable, entre cincuenta y ochenta, en función de las diferencias en los espacios entre ellas. El diámetro de una columna estaría en el rango de 300 a 500 µm, con relativamente poca variación entre especies que difieren varios órdenes de magnitud en el tamaño global del cerebro.
Jones (12) señala que la hipótesis de una organización columnar, principalmente en las cortezas sensoriales, requiere un patrón de conectividad intrínseco, caracterizado por su orientación vertical y por la distribución del imput hacia regiones supra e infragranulares, desde las neuronas de la lámina IV, donde terminan las proyecciones procedentes de los núcleos talámicos. De igual manera, el autor observa que la cuestión de la dimensión de las presuntas columnas corticales es crítica, pues se trataría de determinar una estructura lo suficientemente pequeña, como para que efectivamente sea una unidad elemental de procesamiento, independiente de las particularidades del área cortical donde se encuentre localizada.
Quizá el pionero de esta idea halla sido Creutzfeldt (1977), quien señala que la estructura de la corteza cerebral es homogénea en toda su extensión, con variaciones solamente en la procedencia de sus aferencias y en el destino de sus proyecciones eferentes (2). Jones (12) indica que los referidos patrones de conectividad constituirían la clave final en la determinación de las columnas, sólo sí es posible demostrar que efectivamente existe un ";patrón repetitivo de conectividad intrínseca".
En esta dirección, Somogyi et al. (34) señalan que es posible establecer un patrón estándar de conectividad intrínseca (al que denominan circuito cortical básico, o circuito canónico) en la corteza cerebral sobre la base de la conectividad sináptica y de las características moleculares de esas mismas sinapsis. Se trata de un dispositivo de conexiones entre tipos de células diferentes, mediados por sinapsis excitatorias o inhibitorias, el cual se repite iterativamente en cada una de las láminas corticales, en función de las presiones evolutivas por el aumento en la capacidad de procesamiento.
Los investigadores proponen una caracterización de la corteza cerebral desde esta circuitería básica, en la cual destacan que:
El córtex cerebral consiste de 1. - Una amplia población de neuronas principales (piramidales), conectadas recíprocamente con el tálamo y entre sí, vía axones principales y colaterales, los cuales liberan aminoácidos excitatorios (principalmente glutamato) y 2.- Por una población más reducida de neuronas inhibitorias, principalmente GABAérgicas, que conforman circuitos locales, con las neuronas piramidales y entre sí. (34)
Buxhoevedeen y Casanova (5) han señalado que el concepto de columnas celulares permite establecer una correlación más directa entre la función y la estructura de la corteza cerebral de mamíferos. Estos autores destacan que las minicolumnas constituyen una especie de matriz básica (template) de la organización cortical, que constituye una organización completa en sí misma. Este hecho concede la ventaja de la modularidad: una columna puede establecer interacciones con unidades similares adyacentes o distantes, que aumentan de esta manera la complejidad de los niveles de procesamiento de la información.
No obstante lo anterior, los investigadores creen que es erróneo considerar a las minicolumnas como unidades repetitivas tipo clones; por el contrario, aunque homogéneas en su diseño básico, definido por proyecciones de entrada (inputs), proyecciones de salida (outputs) y por los patrones de circuitería interna, las minicolumnas pueden presentar, sin embargo, variaciones considerables dependiendo, entre otros factores, de su localización en la corteza, así como de su conectividad extrínseca. Por otra parte, los autores piensan que el análisis de las variaciones en la organización de las minicolumnas constituiría una clave en la identificación de los mecanismos fisiopatológicos en trastornos como el autismo, la esquizofrenia o el síndrome de Down (5).
Columnas corticales, desarrollo y evolución
Una de las líneas de argumentación más interesantes y más sólidas que sustentan la hipótesis de la organización modular en columnas verticales de la corteza cerebral proviene de las investigaciones del desarrollo temprano de la misma, durante el período de la llamada corticogenia. En particular, Rakic (35), (36),(37) ha propuesto la hipótesis de la unidad radial, en la cual se describe la secuencia de eventos en el desarrollo cortical que luego conducirá a la organización en columnas en la corteza cerebral madura. Se parte de señalar que en primates, incluidos humanos, la formación de la corteza cerebral tiene lugar durante la primera mitad del desarrollo embrionario.
Este fenómeno se inicia en la zona proliferativa de la región periventricular del cerebro embrionario, donde los primordios de las neuronas corticales (neuroblastos) se multiplican mediante ciclos sucesivos de división mitótica. Al final de este proceso, después de la última división celular, las neuronas corticales 'migran' en dirección a la superficie pial y conforman una organización descrita como inside-out. En este patrón, las neuronas más recientes sobrepasan a las ya establecidas en la placa cortical, formando una ";columna radial", la cual constituye la unidad fundamental de la corteza en desarrollo (35).
Las vías a lo largo de las cuales se produce la migración celular descrita están constituidas por una estructura radial de células gliales (scalfold), la cual tiene sólo una existencia transitoria durante la fase migratoria. La hipótesis de la unidad radial señala que en el plano horizontal la posición de las células en la corteza madura dependerá de la posición relativa de las células precursoras, en la zona proliferativa periventricular, la cual constituye de esta manera una suerte de protomapa de la corteza en formación (35). Por otra parte, la posición de las células dentro de una columna vertical estaría dada por el momento en el cual fueron generadas. De esta manera, todas las células que están localizadas dentro de la misma columna comparten el mismo lugar de origen, migran por el mismo conjunto de células gliales radiales, pero han sido generadas durante diferentes momentos.
Desde este punto de vista, el tamaño de la superficie cortical estaría determinado por el número de las columnas radiales, formadas a su vez por el número de divisiones mitóticas (simétricas) de las células progenitoras, antes del inicio de la migración celular. Mientras que las variaciones y el espesor de la corteza dependerían del número de células dentro de cada columna, determinado a su vez por el número de divisiones asimétricas de las células progenitoras, después del inicio de la migración celular (35),(37).
Coherente con lo anterior, en mamíferos la evolución de la corteza cerebral parece seguir una tendencia hacia el aumento de la superficie, cuyo incremento se estima hasta en mil veces; mientras que el espesor cortical permanece relativamente estable. De esta manera, la expansión de la corteza se produciría en función de ";… cambios en la cinética proliferativa de las células fundadoras en la zona ventricular" (37), lo cual produce como resultado el aumento de las columnas corticales, sin cambios significativos en el espesor cortical. En este sentido, Korman y Rakic (38) han descrito cambios en la duración del ciclo celular y, consecuentemente, en el número de divisiones mitóticas entre especies de roedores y primates, que muestran un aumento hasta de cinco veces, en el caso de los primates.
La estructura de las columnas corticales
Desde las aproximaciones iniciales de Llorente de Nó (5),(11), la estructura de los módulos o columnas corticales se ha descrito a partir de tres elementos básicos. En primer lugar, los componentes neuronales, que constituyen la base celular de esta estructura; en segundo lugar, el conjunto de proyecciones que entran y salen del módulo, constituyendo su conectividad extrínseca, y, en tercer lugar, las conexiones internas que conforman los llamados circuitos intrínsecos.
Los componentes neuronales de las columnas corticales, a pesar de su crecido número, se pueden clasificar en dos tipos principales de acuerdo con su morfología externa y con su función en el procesamiento de la información (4). Hacia finales del siglo XIX, Golgi propuso establecer una diferenciación básica entre neuronas cuyos axones abandonan la corteza cerebral, para terminar en regiones relativamente distantes de su origen, a las cuales denominó neuronas tipo I, y neuronas cuyos axones de corta trayectoria se ramifican localmente, para las cuales propuso la denominación de neuronas tipo II (39).
De esta manera, sobre la base de las características del axón, las células del tipo I de Golgi corresponden a neuronas de proyección, mientras que las células del tipo II, a interneuronas, las cuales conforman circuitos locales generalmente inhibitorios, cuya morfología y localización en la corteza varía en función de las conexiones específicas que establecen (11),(34),(40).
Por otra parte, Le Vay (1973) introdujo otro importante criterio de clasificación de las neuronas corticales, basado en la presencia de espinas sinápticas en la membrana celular. Desde esta perspectiva, las neuronas de la corteza se dividen en dos grandes grupos: espinosas y lisas (4). Dentro del primer grupo se incluyen la totalidad de las neuronas piramidales, así como algunos tipos de interneuronas de la lámina IV, las cuales constituyen los blancos primarios para las proyecciones talámicas en las cortezas sensoriales. Por otra parte, el grupo de las neuronas lisas o poco espinosas está conformado por todas las demás interneuronas, localizadas en diferentes zonas del espesor cortical (5), (41).
Las neuronas de proyección constituyen el 70%-80% de todas las neuronas de la corteza cerebral de los mamíferos (4),(42) y su morfología se ajusta, en términos generales, al modelo de las células piramidales (Fedelman y Peters, 1984). Estas células se encuentran presentes en toda la corteza cerebral, a excepción de la lámina I, pero son particularmente evidentes, en las láminas II-III y V-VI, donde se originan los principales sistemas eferentes de la corteza cerebral (11).
Las células piramidales, aunque constituyen el tipo de neurona más abundante de la corteza cerebral de mamíferos, presentan un fenotipo relativamente homogéneo, y las variaciones se observan principalmente en el tamaño del soma y en la extensión de sus procesos, características que constituyen los criterios básicos para su clasificación. Por otra parte, se ha establecido claramente que la mayoría de las neuronas de proyección utilizan neurotrasmisores aminoacídicos, principalmente glutamato y en menor medida aspartato, cuya acción en la membrana postsináptica es despolarizante, es decir, produce un efecto general excitatorio (43),(44),(45).
Las neuronas del segundo tipo, aunque menos numerosas que las anteriores, presentan, sin embargo, una considerable variación en su morfología, al punto que se han descrito alrededor de quince tipos distintos (40),(41). Esta variabilidad ha dado lugar a la utilización de diversas denominaciones para caracterizarlas, como células estrelladas, células granulares o neuronas no piramidales (46); pero prevalece la denominación de interneuronas, para indicar su función preponderante en el establecimiento de circuitos locales entre neuronas adyacentes. De igual manera, se ha podido establecer que la gran mayoría de estas células utiliza el ácido gama amino butírico (GABA) como neurotransmisor, cuya acción sobre la membrana postsináptica es de hiperpolarización, lo que causa un efecto general inhibitorio (47),(48).
Los criterios para clasificar las interneuronas incluyen principalmente los aspectos morfológicos clásicos, especialmente el tipo de ramificación de los axones (46); los criterios asociados con patrones característicos de respuesta eléctrica (49), (50), (51); así como características bioquímicas y moleculares específicas, como expresión de neuropéptidos, moléculas de la superficie celular o proteínas específicas (52). En esta última dirección, un desarrollo muy significativo de la investigación ha estado asociado con la identificación de proteínas atrapadoras de calcio, pertenecientes a la familia hf-hand, las cuales parecen funcionar como búferes fisiológicos para el control del exceso de ca++ citoplasmático, cuyos efectos excitotóxicos han sido documentados ampliamente en la literatura médica (53).
Sin embargo, aparte de su función fisiológica, se ha establecido que distintas proteínas de este tipo —principalmente parvalbúmina, calbindina y calretinina— se expresan de manera diferencial en subpoblaciones de interneuronas corticales, con muy poco grado de superposición, lo cual las convierte en marcadores muy útiles en la identificación de grupos particulares de neuronas no piramidales (42),(52), (54), (55).
En lo relacionado con el segundo elemento definitorio de los módulos corticales, las proyecciones extrínsecas, habría que señalar que las proyecciones que ingresan a los módulos (aferentes), de hecho, definen el carácter y la extensión de las columnas corticales (11),(30). Las proyecciones procedentes de los núcleos sensoriales específicos del tálamo delimitan el territorio de los módulos localizados en las cortezas sensoriales. Estas proyecciones talámicas terminan en láminas intermedias sobre neuronas estrelladas espinosas o sobre piramidales estrelladas, las cuales constituyen las únicas excepciones a la apariencia lisa, característica de las demás interneuronas corticales. Igualmente, algunas fibras talámicas terminan sobre las dendritas basales de las neuronas piramidales de la lámina III (17),(56),(57). Esta organización sináptica constituye el mecanismo básico para la distribución vertical de la información sensorial aferente, junto con otros dispositivos, como la inhibición lateral ejercida por células en cesta y la función de interneuronas, por ejemplo, las de doble bouquet, cuya disposición espacial estabiliza la estructura de las columnas (58),(59),(60).
Los módulos de las cortezas de asociación, por otra parte, están definidos por las proyecciones córticocorticales, que se originan principalmente en los axones de neuronas piramidales de la lámina III. Estos haces de fibras, en general, siguen dos trayectorias: proyecciones dentro del mismo hemisferio, formando vías intracorticales o ipsilaterales o, bien, proyecciones a regiones corticales homólogas del otro hemisferio, para conformar vías contralaterales que cursan a través del cuerpo calloso (21).
Otros aferentes a los módulos corticales proceden de las fibras horizontales de la lámina I, las cuales hacen sinapsis con los 'penachos' terminales de dendritas apicales de las neuronas piramidales. De igual manera, arriban a las columnas colaterales axónicos horizontales, procedentes de neuronas piramidales localizadas en módulos adyacentes (61), (62), así como de interneuronas inhibitorias, particularmente proyecciones horizontales de células en cesta.
Las proyecciones de salida de los módulos corticales, de manera general, se pueden caracterizar como segregados en tres sistemas eferentes. El primero está constituido por proyecciones que se originan principalmente en neuronas piramidales de la lámina V, las cuales tienen como blanco estructuras subcorticales localizadas a la altura del diencéfalo, del tallo cerebral y de la médula espinal. Estas eferentes corticales eventualmente constituirán el gran sistema de salida de la corteza cerebral hacia los efectores musculares, responsables de las respuestas motoras del organismo (63).
El segundo sistema, ya mencionado, se origina en neuronas piramidales de la lámina III y conforma proyecciones córtico-corticales, las cuales constituyen un sistema de asociación que evolutivamente se va haciendo más voluminoso, hasta llegar a constituir un sistema masivo en primates (64). El tercer sistema se origina en neuronas de proyección de la lámina VI, las cuales dirigen eferencias al tálamo y a su vez reciben proyecciones recíprocas de esta estructura y constituyen un circuito córtico-talámico-cortical que regula el ingreso de la información sensorial a la corteza cerebral (21),(65).
Circuitos intrínsecos
El concepto de circuitos intrínsecos de los módulos o columnas corticales se refiere al patrón de conexiones sinápticas que se establece entre los componentes neuronales dentro del dominio del módulo, el cual está definido, como ya se estableció, bien sea por las aferencias talámicas, las proyecciones córticocorticales o por grupos discretos de fibras eferentes que parten de la corteza motora.
Los contactos sinápticos pueden clasificarse en tres tipos generales, desde el punto de vista fisiológico: excitatorios, inhibitorios y modulatorios, de acuerdo con el efecto que ejerzan sobre la membrana postsináptica. Sin embargo, desde el punto de vista morfológico, se han establecido correlaciones entre este efecto funcional y la estructura sináptica. De esta manera, Grey (1959) y Colonier (1968) describen diferencias estructurales que llevan a proponer la denominación de sinapsis simétricas, para las inhibitorias, y sinapsis asimétricas, para las excitatorias, basados en la observación de microscopía electrónica (4). Por otra parte, además del criterio anteriormente señalado, las sinapsis pueden clasificarse en función de los elementos celulares que se relacionan mediante este contacto. Así, se tendrían cuatro tipos básicos de sinapsis: axosomáticas, axodendríticas, axoaxónicas y dendrodendríticas.
Finalmente, como fue mencionado en otro lugar, una característica morfológica diferencial de las células excitatorias corticales, incluidas todas las neuronas de proyección y un grupo de interneuronas de láminas intermedias, lo constituye la presencia de espinas en su superficie celular. Estas estructuras corresponden a protrusiones de la membrana celular, dotadas de una estructura particular, cuya funcionalidad en principio es el aumento y especialización de las superficies de contacto sináptico (39),(66).
El núcleo central en la organización de los módulos corticales está constituido principalmente por el conjunto de neuronas piramidales, que dan origen a los sistemas eferentes de la corteza cerebral (5). Sobre la base de esta consideración es posible describir el patrón de contactos sinápticos dentro de una columna cortical, teniendo como referente básico el eje definido por las neuronas piramidales, alrededor de las cuales se organizan los diferentes contactos sinápticos, siguiendo un patrón constante, en el cual diferentes tipos de interneuronas realizan contactos específicos, en regiones definidas de la topografía celular de la neurona piramidal.
Para ilustrar este patrón de conectividad se considerarán a continuación cinco de los tipos 'canónicos' de contacto sináptico en el interior de un módulo cortical estándar, definidos tanto por la zona en la cual se presentan como por el tipo de interneurona comprometida.
1. Sobre el soma de las neuronas piramidales y sobre sus dendritas proximales se ha descrito la formación de contactos sinápticos múltiples, de tipo simétrico realizado por las células en cesta, especialmente en las láminas III y V (67). El axón de estas neuronas realiza proyecciones horizontales considerablemente extensas, hasta 1.800 µm (1). Estas características morfológicas han dado pie para proponer que las células en cesta realizan una inhibición lateral de los módulos corticales adyacentes y que con ello establecen una mejor definición de la señal procesada en un módulo específico, frente a señales competitivas de los otros módulos (68). Por otra parte, se ha señalado que estas neuronas, junto con interneuronas espinosas de las láminas intermedias, constituyen el principal blanco de las aferencias talámicas excitatorias, en la corteza cerebral (34),(41).
2. En el segmento inicial del axón de las neuronas piramidales de las láminas III y V se ha descrito un tipo particular de sinapsis realizado por las células en Chandelier (34),(59). La morfología de estas neuronas se caracteriza por la presencia de un axón con múltiples terminales verticales cortos, los cuales forman un dispositivo sináptico múltiple denominado cartridges. La estructura de la neurona en Chandelier le permite establecer contacto con varios cientos de células piramidales, pero dentro del dominio de un módulo cortical, pues su arborización no excede las 300-350 µm. Las sinapsis que forman estas neuronas son de tipo simétrico, por lo tanto inhibitorias y gabérgicas. Su ubicación estratégica, en la 'zona de disparo' de los potenciales de acción, ha llevado a considerarlas como el más poderoso control inhibitorio de la actividad excitatoria de las neuronas piramidales (59).
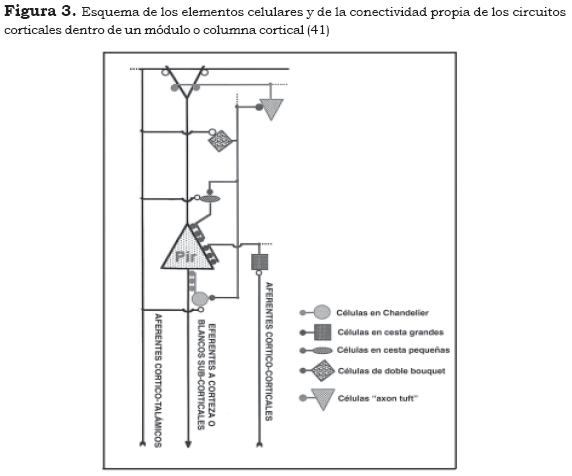
3. En la zona de la dendrita apical, láminas II y III, para el sistema eferente supragranular, se han descrito dos tipos de interneuronas: las células de doble bouquet y las células bipolares. Las primeras se caracterizan por poseer densos agregados axónicos que cursan desde la lámina II a la V (34). Clásicamente se propuso que estas células, por su localización, formarían sinapsis con las dendritas apicales ascendentes; sin embargo, los hallazgos apuntan a señalar que forman sinapsis simétricas sobre tallos y espinas de dendritas basales y ramas oblicuas de la dendrita apical, pero no sobre esta última directamente (54). Se ha propuesto que las células de doble bouquet, por la organización espacial de sus axones, contribuirían a definir funcionalmente unidades modulares dentro de las columnas, produciendo una ";fuerte corriente inhibitoria verticalmente orientada" (11) que tendría efecto sobre piramidales de distintas láminas, pero también sobre otras interneuronas inhibitorias, especialmente sobre células de doble bouquet y Chandelier. Por otra parte, la aferencia excitatoria principal a las neuronas de doble bouquet se origina en proyecciones córtico-corticales (11),(41). Las células bipolares tienen una morfología y una localización similar a las anteriores, razón por la cual algunos autores las clasifican dentro de la misma categoría (1); sin embargo, a diferencia de aquéllas, se caracterizan por tener un plexo axonal menos abundante y más estrecho: menos de 100 µm, el cual contribuye igualmente al mantenimiento de la estructura modular vertical de la corteza cerebral. Sin embargo, estas células conformarían sinapsis asimétricas, no gabérgicas, sobre las espinas dendríticas de las células piramidales (54), es decir, tendrían un efecto de signo contrario en el mismo territorio de las células de doble bouquet (4),(69).
4. En el extremo superior de las neuronas piramidales, correspondiente a la lámina I, se ha descrito un tipo de interneuronas exclusivo de esta lámina, denominado células de Cajal-Retzius. Se trata de células poco numerosas, cuyos procesos se orientan horizontalmente y se distribuyen ampliamente para formar sinapsis inhibitorias, principalmente con las estructuras terminales, bifurcaciones o 'penachos' de las dendritas apicales de las neuronas piramidales (70). Se ha señalado que estas células, las primeras en aparecer durante el desarrollo cortical en mamíferos, son críticas en el desarrollo de la organización laminar de la corteza cerebral, por la secreción de la proteína denominada reelina, la cual estaría asociada con la formación de los estratos corticales (71),(72). Estas células son más evidentes en el cerebro de individuos jóvenes, pero sufren transformaciones muy significativas en el curso del desarrollo, en lo concerniente a la extensión de sus procesos, los cuales terminan por cubrir toda la extensión de la corteza cerebral (8),(9),(70).
5. En el extremo opuesto a las anteriores se ha descrito otro tipo de célula, denominada célula de Martinoti. Los somas de estas neuronas están localizados principalmente en la lámina VI, pero se encuentran en todos los estratos corticales. Sus axones se extienden por todo el espesor de la corteza cerebral para terminar en la lámina I y hacen sinapsis simétricas, inhibitorias sobre los penachos de las dendritas apicales de las neuronas piramidales (40). Al igual que las células de Cajal-Retzius de la lámina I, estas neuronas aparecen tempranamente en el desarrollo, expresan reelina y, probablemente, en interacción con aquéllas, constituyen elementos cruciales en la formación de la matriz laminar de la corteza (41). Se ha señalado que los terminales axónicos de las células de Martinoti entran a la lámina I, acopladas con los terminales o penachos de las dendritas apicales del mismo estrato cortical, conformando de esta manera un nuevo 'amarre', desde 'abajo' de la organización modular vertical (70).
Finalmente, otra característica importante del patrón sináptico intrínseco de las columnas corticales está dado por las conexiones recíprocas que se establecen entre los distintos tipos de interneuronas corticales. Somogyi et al. (34) señalan que un porcentaje pequeño ";pero significativo" de las proyecciones de interneuronas se dirige a otras células del mismo tipo, tanto en la neocorteza como en el hipocampo, donde las células en cesta son la principal fuente de tales proyecciones.
Por otra parte, Fukuda y Kosaka (73) han señalado que las interneuronas que expresan la proteína atrapadora de calcio (parvoalbúmina), es decir, las células en cesta y las células en Chandelier, presentan un doble tipo de interconexión recíproca; por una parte, sinapsis químicas entre terminales axónicos y soma; por otra, presentan sinapsis de tipo eléctrico, mediadas por uniones estrechas (gap junctions) entre dendritas. Esta estructura dual, especialmente las uniones estrechas, podría tener un papel importante en la sincronización de la respuesta eléctrica o actividad oscilatoria, de amplias poblaciones neuronales.
Por su parte, Benes y Beretta (41) señalan que en la corteza cerebral redes de interneuronas inhibitorias, interconectadas recíprocamente, organizarían 'trenes de descarga' de las neuronas piramidales, que resultan en oscilaciones coherentes, las cuales se han propuesto como agrupamientos de características para la formación de representaciones unitarias en la corteza cerebral.
Bibliografía
1. Jones E. History of the cortical cytology. In: Cerebral cortex. New York: Plenum Press; 1984. p. 1-29. [ Links ]
2. Creuttzfeldt OD. Cortex cerebri: performance, structural and functiopnal organization of the cortex. Oxford: Oxford University Press; 1995. [ Links ]
3. Le Brrun Kempler T, Galaburda AM. Principles of cytoarchitectonics. In: Cerebral cortex. New York: Plenum Press; 1984. p. 35-57. [ Links ]
4. Peters A, Jones EG. Classification of cortical neurons. In: Cerebral cortex. New York: Plenum Press; 1984. p. 107-21. [ Links ]
5. Buxhoevedeen DP, Casanova MF. The minicolumn hypothesis in neuroscience. Brain 2002;125:935-51. [ Links ]
6. Glickstein M. Santiago Ramón y Cajal. In: Encyclopedia of life sciences. Macmillan Publishers Ltd., Nature Publishing Group; 2002. [ Links ]
7. Berciano J, Lafarga M, Berciano M. Santiago Ramón y Cajal. Neurología 2001;16(3):118-21. [ Links ]
8. Marín-Padilla M. Cajal-Retzius cells and the development of the neocortex. Trends in Neurosciences 1998;21(2):64-71. [ Links ]
9. Marín-Padilla M. Evolución de la estructura de la neocorteza del mamífero: nueva teoría citoarquitectónica. Revista de Neurología 2001;33:843-53. [ Links ]
10. Mountcastle V. Introducción. Cerebral Cortex 2003 Jan;13:2-4; 1047-3211. [ Links ]
11. Mountcastle V. The columnar organization of the neocortex. Brain 1997;120:701-22. [ Links ]
12. Jones E. Microcolumns in the cerebral cortex. Proc Nat Acad Scien 2000; 97(10):5019-21. [ Links ]
13. Dow BM. Orientation and color columns in monkey visual cortex. Cerebral Cortex 2002;12(10):1005-15. [ Links ]
14. Girman S, Sauve I, Lund R. Receptive field properties of single neurons in rat primary visual cortex. Journal of Neuro Psysiology 1999. [ Links ]
15. Preuss T, Huixin Q, Kaas J. Distinctive compartmental organization of human primary visual cortex. Proc Natl Acad Sci 1999;96:11601-6. [ Links ]
16. Simonds DJ, Land PW. Cortical barrels: maps and plasticity. In: Encyclopedia of life sciences. Nature Publish. [ Links ]
17. Miller KD, Pinto D, Simons DJ. Processing in layer IV of the neocortical circuit: new insights from visual and somatosensory cortex. Currrent Opinion in Neurobiology 2001;11:486-97. [ Links ]
18. Levine MW, Shefner JM. Fundamentals of sensation and perception. Pacific Grove (CA): Cole Publishing Co; 1991. [ Links ]
19. Schreiner CE, Read HL, Sutter ML. Modular organization of frecuency integration in primary auditory cortex. Annual Review of Neurosciences 2000;23:501-29. [ Links ]
20. Read HL, Winer JA, Schreiner. Modular organization of intrinsic connections associated with spectral tuning in cat auditory cortex. Procedeengs of National Academy of Science 2001;98(14):8042-7. [ Links ]
21. Escobar MI, Pimienta H, Cavines VS, Jacobson M, Crandall JE, Kosik KS. Architecture of apical dendrites in the murine neocortex: dual apical dendritic systems. Neuroscience 1986;17(4):975-89. [ Links ]
22. Lev DL, White EL. Organization of pyramidal cell apical dendrites and composition of dendritic clusters in the mouse: emphasis on primary motor cortex. European Journal of Neuroscience 1997; 9(2):280-90. [ Links ]
23. Keller A. Intrinsic synaptic organization of the motor cortex. Cerebral Cortex 1993;3: 430-41. [ Links ]
24. Rizzolatti G, Luppino G. The organization of the cortical motor system: new concepts. Electroencephalography and Clinical Neurophysiology 1998;106:238-96. [ Links ]
25. Sanes JN, Donoghue JP. Plasticity and primary motor cortex. Annual Review of Neuroscience 2000;23:393-415. [ Links ]
26. Goldman PS, Nauta WJ. Columnar distribution of cortico-cortical fibres in the frontal association, limbic and motor cortex of the developing rhesus monkey. Brain Research 1977;122(3):393-413. [ Links ]
27. Bugbee NM, Goldman-Rakic PS. Columnar organization of corticocortical projections in squirrel and rhesus monkeys: similarity of column width in species differing in cortical volume. Journal of Comparative Neurology 1983;220(3):355-64. [ Links ]
28. Levitt JB, Lewis DA, Yoshioka T, Lund JS. Topography of pyramidal neuron intrinsic connections in macaque monkey prefrontal cortex (areas 9 and 46). Journal of Comparative Neurology;338(3):360-76. [ Links ]
29. Kritzer MF, Goldman-Rakic PS. Intrinsic circuit organization of the major layers and sublayers of the dorsolateral prefrontal cortex in the rhesus monkey. Journl of Comparative Neurology 1995;359 (1):131-43. [ Links ]
30. Pucak ML, Levitt JB, Lund JS, Lewis DA. Patterns of intrinsic and associational circuitry in monkey prefrontal cortex. Journal of Comparative Neurology 1996; 376(4): 614-30. [ Links ]
31. Eccles JC. The cerebral neocortex: a theory of its operation. In: Cerebral cortex. New York: Plenum Press; 1984. p. 1-24. [ Links ]
32. Eccles JC. Evolution of consciousness. Proc Natl Acad Sci 1992;89:7320-4. [ Links ]
33. Beck F, Eccles JC. Quantum aspects of brain activity and the role of consciousness. Proc Natl Acad Sci 1992; 89: 11357-61. [ Links ]
34. Somogyi P, Tamás G, Luján R, Bhul EH. Salient features of synaptic organization in the cerebral cortex. Brain Research Reviews 1998;26(2-3): 113-35. [ Links ]
35. Rakic P. Specification of cerebral cortical areas. Science 1988;241(4862):170-6. [ Links ]
36. Rakic P. Radial vs. tangential migration of neuronal clones in the developing cerebral cortex. Proc Nat Acad Sciences 1995; 92:11323-7. [ Links ]
37. Rakic P. Radial unit hypothesis of neocortical expansion. Novartis Foundation Symposium 2000;228:42-52. [ Links ]
38. Kornack DR, Rakic P. Changes in cellcycle kinetics during the development and evolution of primate neocortex. Proc Nat Acad Sciences 1998;95:1242-6. [ Links ]
39. Fedelman ML. Pyramidal neurons. In: Cerebral cortex. New York: Plenum Press; 1984. [ Links ]
40. Gupta A, Wang Y, Markran H. Organizing principles for a diversity of gabaergic interneurons and synapses in the neocortax. Science 2000;287:273-8. [ Links ]
41. Benes FM, Beretta S. Gabaergic interneurons: implications for understanding schizophrenia and bipolar disorder. Neurpshichopharmacology 2001;26 (1):1-27. [ Links ]
42. Hoff R, Glezer II, Condé F, et al. Cellular distribution of the calcium-binding proteins parvalbumin, calbindin, and calretinin in the neocortex of mammals: phylogenetic and developmental patterns. Journal of Chemical Neuroanatomy 1999;16:77-116. [ Links ]
43. Hill E, Kalloniatis M, Tan SS. Glutamate, GABA and precursor amino acids in adult mouse neocortex. Cerebral Cortex 2000; 10(11):1132-42. [ Links ]
44. Wang Y, Gupta A, Markram H. Anatomical and functional differentiation of glutamatergic synaptic innervation in the neocortex. Journal of Physiology 1999; 93:305-17. [ Links ]
45. Conti F, Weinberger R. Shaping Excitation at glutamatergic synapses. Trens in Neurosciences 1999;22:451-8. [ Links ]
46. Fairén A, DeFelipe J, Regidor J. Nonpyramidal neurons: general account. In: Cerebral cortex New York: Plenum Press; 1984. p. 201-53. [ Links ]
47. Bormann J. The ABC of GABA receptors. Trends on Pharmachological Sciences 2000;21(1):16-9. [ Links ]
48. DeFelipe J. Cortical interneurons: from cajal to 2001. Progress in Brain Research 2001; 136:215-38. [ Links ]
49. Caulli B, Audinat E, Lambolez B, et al. Molecular and physiological diversity of cortical nonpyramidal cells. The Journal of Neuroscience 1997;17(10):3894-906. [ Links ]
50. Kawaguchi Y, Kubota Y. Gabaergic cell subtypes and their synaptic connections in rat frontal cortex. Cerebral Cortex 1997;7:476-86. [ Links ]
51. Kawaguchi Y, Kubota Y. Neurochemical features and synaptic connections of large physiologically-ientified gabaergic cells in the rat frontal cortex. Neuroscience 1998;85(3):677-701. [ Links ]
52. De Felipe J. Neocortical neuronal diversity: chemical heterogeneity revelated by colocalization studies of classic neurotransmitters, neuropeptides, calcium - binding proteins, and cell surface molecules. Cerebral Cortex 1993; 3 (4):273-89. [ Links ]
53. Schwaller B. Calcium-binding proteins. In: Encyclopedia of life sciences. Nature; 2001. p. 1-8. [ Links ]
54. DeFelipe J. Types of neurons, synaptic connections and chemical characteristics of cells immunoreactive for CB, PV, CR in the neocortex. Journal of Chemical Neuroanatomy 1997;14:1-19. [ Links ]
55. Raynolds GP, Beasley CL. Gabaergic neuronal subtypes in the human frontal cortex development and deficits in schizophrenia. Journal of Chemical Neuroanatomy 2001;22:95-100. [ Links ]
56. Petersen C, Sakmann B. The excitatory neuronal network at rat layer IV barrel cortex. The Journal of Neuroscience 2000;15:7579-86. [ Links ]
57. Lubke J, Egger V, Sakmann B, Feldmeyer D. Columnar organization of dendrites and axons of single and synaptically coupled excitatory spiny neurons in layer 4 of the rat barrel cortex. Journal of Neurosciences 2000;20(14):5300-11. [ Links ]
58. DeFelipe J, Jones EG. Vertical Organization of Gamma-aminobutyric acidaccumulating Intrinsic neuronal systems in monkey cerebral cortex. Journal of Neuroscience 1985;12:3246-60. [ Links ]
59. DeFelipe J, Endry SH, Hashikawa T, Molinari, Jones EG. A microcolumnar structure of monkey cerebral cortex revelated by inmunocytochemical studies of doble bouquet cell axons. Neuroscience 1990;37:655-73. [ Links ]
60. Krimer LS, Goldman-Rakic PS. Prefrontal microcircuits: membrane properties and excitatory imput of local, medium and wide arbor interneurons. The Journal of Neuroscience 2001;21(11):3788-96. [ Links ]
61. González-Burgos G, Barrionuevo G, Lewis D. Horizontal synaptic connections in monkey prefrontal cortex: an in vivo electrophysiological study. Cerebral Cortex 2000;10:82-92. [ Links ]
62. Lewis D, González-Burgos G. Intrinsic excitatory connections in the prefrontal cortex and the pathophysiology of schizophrenia. Brain Research Bulletin 2000;52(5):309-17. [ Links ]
63. Luppino G, Rizzolatti G. The organization of the frontal motor cortex. News Phys Sciences 2000;15:219-24. [ Links ]
64. Barbas H, Rempel-Clower N. Cortical structure predicts the pattern of corticocortical connections. Cerebral Cortex 1997;7:635-46. [ Links ]
65. Deschênes M, Veinante P, Zhang Z. The organization of corticothalamic projections: reciprocity vs. parity. Brain Research Reviews 1998;28(3):286-308. [ Links ]
66. Kuno M. The synapse: function plasticity and neurotrophism. Oxford: Oxford Science Publications; 1995. [ Links ]
67. Akil M, Lewis DA. Diferential distribution of parvalbumin-immunoreactive pericellular clusters of terminal butons in developing and adult monkey neocortex. Exp Neurol 1992;115:2239-49. [ Links ]
68. Lund JS, Yoshioka T, Levitt JB. Comparison of intrinsic connectivity in different areas of macaque monkey cerebral cortex. Cerebral Cortex 1993;3(2):148-62. [ Links ]
69. Río M del, DeFelipe J. Synaptic connections of calretinin-immunoreactive neurons in the human neocortex. The Journal of Neuroscience 1997; 17( 13):5143-64. [ Links ]
70. Marín-Padilla M. Early ontogenesis of the human cerebral cortex. In: Cerebral cortex. New York: Plenum Press; 1984. [ Links ]
71. Curran T, D'Arcangelo G. Role of reelin in the control of brain development 1. Brain Research Reviews 1998;26:285-94. [ Links ]
72. Pesold C, Impagnatello F, Pisu MG. Reelin is preferentially expressed in neurons synthesizing g-aminobutyric acid in cortex and hippocampus of adult rats. Proc Natl Acad Sci USA 1998;95:3221-6. [ Links ]
73. Fukuda T, Kosaka T. The dual network of gabaergic interneurons linked by both chemical and electrical synapses: a possible infraestructure of the cerebral cortex. Neuroscience Research 2000;38(2):123-30. [ Links ]
74. DeFelipe J. Chandelier cells and epilesy. Brain 1999;122:1807-22. [ Links ]
75. Baimbridge KG, Celio MR, Rogers JH. Calcium-binding proteins in the nervous system. Trends in Neurosciences 1992;15 (8):303-8. [ Links ]

