











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Los foraminíferos bentónicos, organismos microscópicos ampliamente distribuidos en los fondos oceánicos alrededor del mundo, destacan por su abundancia y diversidad (Jones, 2014; Saraswati, 2021). Su amplia dispersión en diferentes ambientes ha permitido comprender su comportamiento en relación con la disponibilidad de oxígeno disuelto y de nutrientes, la influencia de las corrientes oceánicas y su distribución en función de la profundidad, entre otros aspectos (Kaiho, 1994; Jorissen, 1999; Sen Gupta, 2003; Jorissen et al., 2007; Koho y Piña-Ochoa, 2012; Murray, 2014). Por lo anterior, los foraminíferos bentónicos son una herramienta comúnmente utilizada en las reconstrucciones paleoambientales (p. ej. Murray et al., 2011; Murray, 2013; Katz et al., 2018; Alegret et al., 2021).
La presencia y abundancia de foraminíferos bentónicos en las secuencias que abarcan desde el Eoceno tardío al Mioceno temprano en el Cinturón Plegado de San Jacinto (CPSJ) en el Caribe colombiano (Figura 1) constituyen la oportunidad de investigar las condiciones fisicoquímicas de los fondos marinos del margen NW de Colombia. Estos ambientes parecen haber sido controlados por la tectónica regional, que incluye periodos de deformación y levantamiento (Duque-Caro, 1975, 1979, 1991), y el aumento en la subsidencia dada por estructuras preexistentes y aporte sedimentario proveniente de los Andes desde el Oligoceno (Mora-Bohórquez et al., 2018, 2020).
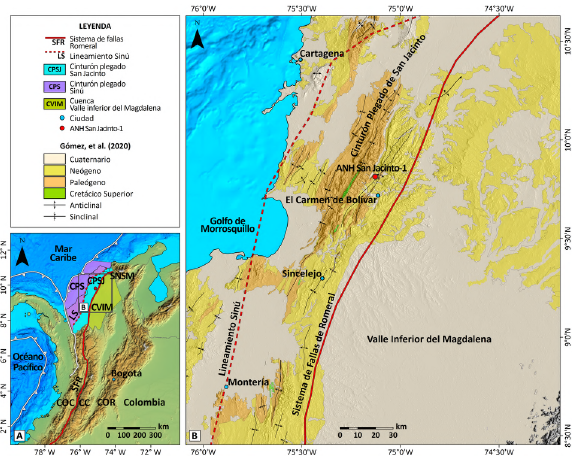
Figura 1 A. Localización del pozo ANH-SAN JACINTO-1 y extensión de las principales cuencas geológicas del Caribe colombiano. B. Mapa geológico del área de estudio basado en Gómez y Montes (2020). Batimetría tomada de GEBCO Compilation Group (2023). Elevación tomada de NASA/METI/AIST/Japan Spacesystems y U.S./Japan ASTER Science Team (2018). CPS: Cinturón Plegado del Sinú. CPSJ: Cinturón Plegado de San Jacinto. CVIM: cuenca del Valle Inferior del Magdalena. SNSM: Sierra Nevada de Santa Marta. COC: Cordillera Occidental. CC: Cordillera Central. COR: Cordillera Oriental. SFR: Sistema de Fallas de Romeral.
La mayoría de los estudios con foraminíferos en el Caribe colombiano para este intervalo de tiempo han sido enfocados en la bioestratigrafía de estos depósitos (Petters y Sarmiento, 1956; Duque-Caro, 1968, 1972; Duque-Caro et al., 1996; Torres et al., 2022; Celis et al., 2023; Duque-Castaño et al., 2023; Plata-Torres et al., 2023), y muy poco se ha explorado su utilidad en la caracterización detallada de los paleoambientes, lo que representa un área de investigación aún por explorar. El conocimiento actual sobre los paleoambientes interpretados en la parte central del CPSJ, en los alrededores del municipio de El Carmen de Bolívar (Figura 1), indica la transición de depósitos marinos poco profundos en el Eoceno tardío hacia depósitos pelágicos de aguas profundas en el Oligoceno y Mioceno temprano (Petters y Sarmiento, 1956; Salazar-Ortiz et al., 2020a). Otros estudios sugieren que, en las regiones adyacentes, los ambientes se desarrollaron en batimetrías menores a 200 metros y en el quiebre de talud durante el Eoceno tardío, y evolucionaron hacia ambientes más profundos en el talud, con batimetrías correspondientes a la zona batial media durante el Oligoceno. Además, se presume la existencia de fondos relativamente anóxicos interrumpidos por eventos episódicos de mayor oxigenación (Duque-Caro, 1971, 1975, 1979; Duque-Caro et al., 1996; Fiorini y Jaramillo, 2007).
El presente estudio tiene como objetivo reconstruir la evolución paleoambiental mediante foraminíferos bentónicos de la sucesión sedimentaria marina que abarca desde el Eoceno superior al Mioceno inferior, registrada por el pozo ANH-SAN JACINTO-1 ubicado en la parte central del CPSJ (Figura 1). Se presenta una reconstrucción en términos de oxigenación de fondo y de contenido de nutrientes, y se relacionan temporalmente sus cambios con los procesos geológicos regionales reportados para la CPSJ y/o globales en el Paleógeno tardío.
Marco geológico
El noroccidente colombiano está localizado en la zona de convergencia entre las placas de Suramérica y Caribe (Toto y Kellogg, 1992; Flinch, 2003). La convergencia de estas placas durante el Cenozoico dio lugar a la formación de diferentes provincias geológicas, entre ellas el Cinturón Plegado de San Jacinto (CPSJ; Duque-Caro, 1979; Flinch, 2003). El CPSJ es una cuenca antearco de rumbo N20°E, desarrollada sobre un basamento oceánico del Cretácico superior, sobre el que se acumularon sedimentos marinos, transicionales y continentales desde el Campaniano hasta el Cuaternario (Duque-Caro etal., 1996; Guzmán-Ospitia, 2007; Silva-Arias et al., 2016; Mora-Bohórquez et al., 2017, 2018; Manco-Garcés et al., 2020; Salazar-Ortiz et al., 2020b; Giraldo-Villegas et al., 2023; Celis et al., 2023; 2024).
El CPSJ limita al sur con las rocas oceánicas del norte de la Cordillera Occidental; al occidente se encuentra el Cinturón Plegado del Sinú, limitado por el lineamiento de Sinú; y al oriente está en contacto con la cuenca del Valle Inferior del Magdalena (VIM), por medio del Sistema de Fallas de Romeral (Duque-Caro, 1979; Guzmán-Ospitia et al., 2004; Barrero et al., 2007; Mora-Bohórquez et al., 2017, 2018, 2020; ver Figura 1). Desde el Eoceno tardío al reciente, la sedimentación en el CPSJ y el VIM se dio en una cuenca antearco sin magmatismo relacionada con un proceso de subducción plana (Mora-Bohórquez et al., 2017). Las unidades del Eoceno superior al Mioceno inferior se sedimentaron en ambientes marinos someros y profundos, en una cuenca subsidente, y con tasas de sedimentación que fueron incrementando durante el Oligoceno tardío y el Mioceno temprano (Mora-Bohórquez et al., 2018).
En el centro del CPSJ, entre el pozo ANH-SAN JACINTO-1 y los alrededores del municipio de El Carmen de Bolívar (Figuras 1 y 2), las rocas sedimentarias del Eoceno superior a Oligoceno inferior están representadas por la Formación San Jacinto, conformada por arenitas, conglomerados y lodolitas, con abundantes foraminíferos bentónicos, sedimentados en ambientes que abarcan desde zonas costeras hasta el quiebre del talud (Petters y Sarmiento, 1956; Duque-Caro, 1967; Duque-Caro et al., 1996; Salazar-Ortiz et al., 2020a; Celis et al., 2024). Los depósitos del Oligoceno inferior al Mioceno inferior hacen parte de la Formación El Carmen (Duque-Caro et al., 1996; Guzmán-Ospitia et al., 2004; Guzmán-Ospitia, 2007; Salazar-Ortiz et al., 2020a). Esta unidad está compuesta por arcillolitas grises y negras, acumuladas en una cuenca marginal, restringida y anóxica (Duque-Caro et al., 1996; Fiorini y Jaramillo, 2007).
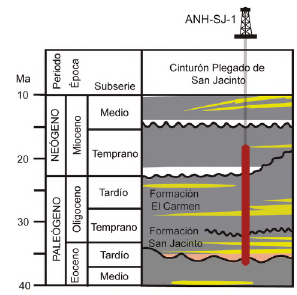
Figura 2 Cronoestratigrafía del Eoceno tardío al Mioceno temprano en la CPSJ (Mora-Bohórquez et al., 2018) y la posición estratigráfica del pozo ANH-SAN JACINTO-1 (ANH-SJ-1) basado en la edad propuesta por Arias-Villegas et al. (2023). Geocronología según Gradstein et al. (2020).
Características yedad del pozo ANH-SAN JACINTO-1
El pozo ANH-SAN JACINTO-1 es un pozo corazonado perforado por la Agencia Nacional de Hidrocarburos (ANH) al norte del municipio de El Carmen de Bolívar, en el Caribe colombiano (9°48'5.47'' N, 75°7'33.95'' W; Figura 1). Este pozo alcanzó una profundidad de aproximadamente 524 m. Sin embargo, debido al buzamiento de los estratos (entre 40° y 60°), el espesor corregido y calculado por Arias-Villegas et al. (2023) es de 342 m. El pozo se divide en tres conjuntos litológicos (UniCaldas, ANH, 2020; Arias-Villegas et al., 2023). Desde la base a los 342 m hasta aproximadamente los 300 m de profundidad, la litología consta de lodolitas calcáreas con restos de moluscos, laminación plana paralela y calizas (packestone y grainstone esqueletales). Localmente, el intervalo presenta sublitoarenitas y litoarenitas masivas y con laminación plana paralela y areniscas bioclásticas. Desde 300 m a 80 m se observan lodolitas masivas de color negro con bioturbación (Chondrites y Taenidium). Entre 80 m y el techo del pozo se identificaron lodolitas calcáreas negras masivas y arcillolitas limosas y calcáreas pardo-amarillentas con laminación paralela, ocasionalmente bioclásticas y bioturbadas (Chondrites y Palaeophycos).
El núcleo fue datado de base a techo mediante nanofósiles calcáreos por Arias-Villegas et al. (2023). Según estos autores, el pozo abarca desde el Eoceno tardío (Priaboniano) hasta el Mioceno temprano (Aquitaniano), y cubre las biozonas NP19-20 (CNE19) a NN2 (CNM1) de Martini (1971), aproximadamente entre 35,2 a 22,5 Ma en la escala geocronológica de Gradstein et al. (2020); Figura 3. De acuerdo con ese modelo de edad, el pozo presenta algunas discontinuidades (Arias-Villegas et al., 2023). En el Eoceno superior se reconoce un hiato de ~0,8 Ma a los 270 m de profundidad. En el Oligoceno inferior, la ausencia de la biozona de nanofósiles calcáreos CNO3 en la profundidad de 176 m sugiere una discordancia de 2 Ma. Una última discontinuidad fue propuesta hacia los 12,6 m, la cual pone en contacto el Oligoceno superior y el Mioceno inferior. Dicha discordancia comprendería un hiato de ~4,4 Ma y fue definida por la ausencia de las biozonas de nanofósiles calcáreos NP25 y NN1. Las muestras en este estudio abarcan las mismas profundidades del trabajo de Arias-Villegas et al. (2023).
Metodología
Las muestras utilizadas en este estudio provienen de lodolitas separadas cada ~6 m a lo largo del pozo, que abarcan los tres conjuntos litológicos. Un total de 60 muestras fueron analizadas en este estudio para foraminíferos bentónicos.
La preparación de las muestras se desarrolló mediante el método de Thomas y Murney (1985), para rocas de grano fino (lodolitas) y una dureza relativamente baja. Se utilizaron aproximadamente 30 g de roca por muestra. La muestra estuvo en remojo en una solución compuesta de agua y peróxido de hidrógeno (~5%), para luego ser lavada en húmedo a través de un tamiz con apertura de malla de 63 μm, y posteriormente secada en un horno (40°C, 24 h). El sedimento seco fue tamizado y dividido en las fracciones >63 μm y >125 μm. Para este estudio se extrajeron todos los foraminíferos bentónicos de la fracción >125 μm. La identificación taxonómica de los foraminíferos planctónicos será abordada en una próxima publicación. Los foraminíferos bentónicos de la fracción inferior fueron excluidos del análisis al tener una mayor susceptibilidad al transporte y redepósito (Lohmann, 1978; Lutze y Coulbourn, 1984), fenómeno que ya ha sido documentado en las sucesiones cenozoicas del CPSJ (p. ej., Bürgl, 1965; Fiorini y Jaramillo, 2007; Duque-Castaño et al., 2023; Ospina-Muñoz et al., 2023; Vallejo-Hincapié et al., 2023). Igualmente, al excluir la fracción más fina, se evita incertidumbres en la identificación taxonómica de ejemplares bentónicos en estado juvenil (Schröder-Adams, 1986; Kurbjeweit et al., 2000; Heinz et al., 2008; Schönfeld et al., 2012; Weinkauf y Milker, 2018).
La identificación de los ejemplares bentónicos se realizó en un estereomicroscopio binocular NIKON SMZ 1500®. Para este estudio, se clasificaron los foraminíferos bentónicos calcáreos hasta el nivel de género, debido a su alta diversidad y gran variabilidad morfológica. Esta aproximación facilita el análisis comparativo a nivel de morfogrupos (la lista de los géneros se encuentra en la Tabla Suplementaria 1). En contraste, los foraminíferos aglutinados, con menor diversidad y morfologías más distintivas, fueron identificados hasta el nivel específico (Tabla Suplementaria 2). Las clasificaciones taxonómicas siguen las propuestas de Van Morkhoven et al. (1986), Bolli et al. (1994), Jones (1994), Kaminski y Gradstein (2005), Holbourn et al. (2013) y Torres et al. (2022). Las especies consideradas como redepositadas del Paleoceno y el Eoceno inferior a medio fueron excluidas de las interpretaciones paleoambientales (Bolli et al., 1994; Kaminski y Gradstein, 2005; Torres et al., 2022).
Se calculó la riqueza y diversidad de especies a través de los índices de riqueza (S) y Shannon (H') (Shannon, 1948; Magurran, 1988), por medio del software PAST versión 4.12 (Hammer et al., 2001). Se estimó la relación entre foraminíferos planctónicos (no clasificados en este estudio) y bentónicos (P/B) utilizando la fórmula P/B= número de especímenes planctónicos/número de especímenes bentónicos (calcáreos y aglutinados). Además, se evaluaron las condiciones cualitativas de oxígeno de fondo a través de la agrupación de los foraminíferos calcáreos en tres niveles de oxigenación (Tabla Suplementaria 3): óxico, subóxico y disóxico. Esto, basado en el conocimiento actual de diferentes grupos cosmopolitas, para lo que se sigue las propuestas de Kaiho (1994), Sen Gupta (2003) y Kranner et al. (2022).
Se agruparon los foraminíferos bentónicos calcáreos y aglutinados en morfogrupos, relacionando la morfología de la concha con sus preferencias paleoecológicas (Tabla 1 y Tablas Suplementarias 1 y 2). En el caso de las formas calcáreas, se estableció un total de 14 morfogrupos fueron establecidos siguiendo las propuestas de Koutsoukos y Hart (1990), Cetean et al. (2011) y Borges-Quadros et al. (2015); mientras que para los aglutinados se identificaron 4 morfogrupos y se siguieron las propuestas de Nagy et al. (1995), Van Den Akker et al. (2000) y Kaminski y Gradstein (2005).
Tabla 1 Morfogrupos de foraminíferos aglutinados y calcáreos, forma de la conchilla y microhábitat, ambientes y principales géneros y grupos. Modificado de Koutsoukos y Hart (1990), Van Den Akker et al. (2000) y Kaminski y Gradstein (2005).
| Morfogrupos aglutinados | Forma de la conchilla | Posición de vida | Ambiente | Géneros principales |
|---|---|---|---|---|
| M1 | Tubular | Epifauna erecta | Batial y abisal | Bathysiphon, Nothia, |
| Psammosiphonella | ||||
| M2a | Globular Troco redondeado | Infauna somera | Batial y abisal | Psammosphaera |
| M2b | y estreptospiral, trocoespiral planoconvexa | Epifauna superficial | Plataforma a aguas marinas profundas | Budashevaella, Trochammina |
| M2c | Quilla alargada | Epifauna superficial | Plataforma a marino marginal | Spiroplectammina |
| M3a | Planispiral aplanada y estreptospiral | Epifauna superficial | Lagoon a abisal | Ammodiscus, Glomospira, |
| M3b | Aplanado irregular | Epifauna superficial | Batial superior a abisal | Ammolagena |
| M4a | Planispiral redondeada | Epifauna superficial y/o infauna somera | Plataforma interna a batial superior | Haplophragmoides, |
| M4b | Subcilíndrico alargado | Infauna profunda | Plataforma interna a batial superior | Karrerulina, Tritaxia |
| Cónico alargado | Infauna profunda | Plataforma interna a batial superior | Ammobaculites, Reophax | |
| Morfogrupo calcáreos | Forma de la conchilla | Posición de vida | Ambiente | Grupos principales |
| CHA1 | Plano-convexo, bajo/alto trocoespiral ancho | Epifaunal | Nerítico a batial medio | Gavelinélidos |
| CHA2 | Cóncavo-convexo, bajo trocoespiral ancho | Epifaunal | Nerítico a batial medio | Bagínidos |
| CHA3 | Inflado, periferia biconvexa ampliamente redondeada | Epifaunal/infaunal somero | Nerítico externo a batial | Alabamínidos y noniónidos |
| CHA4 | Cónico, trocoespiral bajo | Epifaunal | Nerítico externo a batial medio | Epistomárido, cibícidos |
| CHA5 | Lenticular, periferia trocoespiral baja subaguda/carenada | Epifaunal | Nerítico externo a batial | Osanguláridos, eponídidos y planulínidos |
| CHA6 | Lenticular, planispiral periferia subaguda/ carenada | Epifaunal/infaunal somero | Nerítico a batial medio | Vaginulínidos enrrollados |
| CHB1 | Globular/ovada a alargada/fusiforme bilocular/multilocular | Epifaunal/infaunal | Nerítico a batial medio | Chilostomélidos, pleurostomélidos, lágenos |
| CHB2 | Anchas a palmeadas,comprimidas planispirales a | Epifaunal/infaunal | Nerítico externo a batial | Vaginulínidos desenrollados |
| CHB3 | Alargada, recta a arqueada uniseriada o planispiral-uniseriada | Epifaunal/infaunal | Nerítico a batial medio | Nodosáridos |
| CHB4 | Cónico redondeadoalargado triserialbiserial- uniserial | Infaunal | Nerítico a batial medio | Bolivínidos, buliminélidos, fursenkoínidos, |
| CPA | Fusiforme, comprimida ancha | Epifaunal | Nerítico a batial | Espiroloculínidos |
| CPB | Quinqueloculino alargado a ovado | Epifaunal/infaunal somero | Nerítico interior a medio | Hauerínidos |
Resultados
A lo largo del pozo la preservación de los foraminíferos bentónicos en general es moderada a buena. Sin embargo, en algunas muestras se observa evidencia de deformación, recristalización, abrasión y disolución de algunos ejemplares. Todas las muestras fueron productivas. No obstante, las cantidades de foraminíferos fueron variables, oscilaron entre 3 individuos en la profundidad 89,3 m hasta 442 individuos a los 317,1 m (Figura 3).
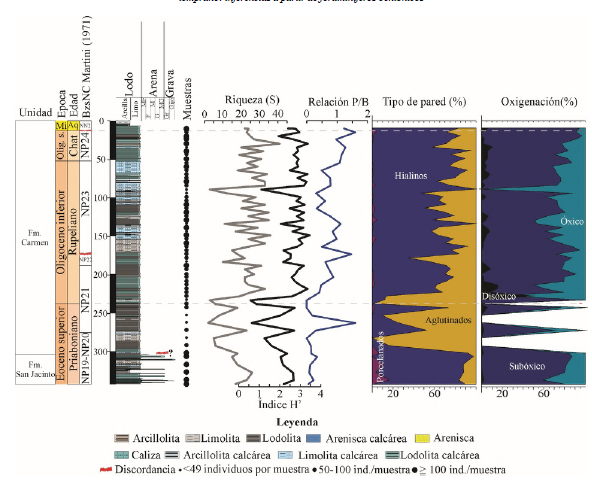
Figura 3 Registros de foraminíferos bentónicos del pozo ANH-SAN JACINTO-1. De izquierda a derecha: Unidades estratigráficas, edad y biozonas de nanofósiles calcáreos (NC) de acuerdo con Arias-Villegas et al. (2023), columna estratigráfica (MF: muy fina, F: fina, M: media, G: gruesa, MG: muy gruesa, Gr: grava). Posición de las muestras analizadas para este estudio con sus abundancias. Riqueza (S), diversidad índice (H'). Relación planctónicos/bentónicos (P/B), distribución de los foraminíferos bentónicos según el tipo de pared y estimación de la oxigenación de fondo a lo largo del pozo. La posible discordancia entre las formaciones San Jacinto y El Carmen está acompañada de un signo de interrogación.
En total se contabilizaron 90 géneros de foraminíferos bentónicos calcáreos (Tabla Suplementaria 1) y 53 géneros de foraminíferos bentónicos aglutinados; estos últimos representados en 105 especies (Tabla Suplementaria 2). La riqueza (S) es predominantemente baja en el Eoceno superior, con un promedio de 5, y alcanza un valor máximo de 18 en la parte superior del Eoceno superior. En los depósitos del Oligoceno y Mioceno, los valores promedios son de 9 (Figura 3). La diversidad entre la base y alrededor de 250 m es fluctuante, pero, desde 250 m y hasta el tope del pozo, los valores se mantienen en promedio superiores a 3, con valores máximos de 4 (Figura 3).
La relación P/B muestra valores mayoritariamente bajos, menores a 0,4, en el Eoceno superior, con un pico elevado de 1,6 a los 267 m. Posteriormente, a finales del Eoceno y comienzos del Oligoceno, los valores disminuyen casi a 0 entre 253 y 223 m. Durante el Oligoceno, la relación incrementa, con valores promedio por encima de 0,4 y alcanza un máximo de 1,5 a los 173,5 m. En el Mioceno, se observan nuevamente valores máximos que llegan 1,6 a los 15 m (Figura 3).
En cuanto a los morfogrupos calcáreos (Figuras 3 y 4), los hialinos (morfogrupos CH) son más constantes a lo largo del pozo. Los morfogrupos calcáreos CHA1 se caracterizan por Anomalinoides spp., Gyroidina spp. y Hansenisca spp., que presentan formas planoconvexas y trocoespirales anchas. El morfogrupo CHA4 está compuesto por Cibicides spp., Cibicidina spp., Cibicidoides spp., Falsoplanulina spp., Helicostegina spp., Heterolepa spp., Lobatula spp. y Nuttallides spp., que exhiben formas cónicas y trocoespirales de baja altura. Y el morfogrupo CHB4 incluye Bolivina spp., Bulimina spp., Cassidulina spp., Epistominella spp., Evolvocassidulina spp., Fursenkoina spp., Globobulimina spp., Praeglobobulimina spp., Protoglobobuliminas spp., Siphogenerina spp., Siphogenerinoides spp., Siphouvigerina spp., Transversigerina spp. y Uvigerina spp., que presentan formas cónicas redondeadas-alargadas triseriales, biseriales y uniseriales. Estos morfogrupos muestran una distribución consistente a lo largo del pozo, con una notable disminución en sus abundancias en la parte superior del Eoceno superior (Figura 4). CHB4 destaca como el más abundante, con un promedio del 24% y máximos del 60% en los depósitos del Mioceno; Uvigerina spp. es el género más abundante de este grupo con máximos del 60% (Figura 5). Por otro lado, CHA1 y CHA4 alcanzan sus máximas abundancias durante el Oligoceno, donde registra un 33% a una profundidad de 89,3 m y un 42% a 134 m, respectivamente (Figura 4). Los géneros Anomalinoides spp. (CHA1), con valores hasta del 33%, y Cibicidoides spp. (CHA4), con máximos del 42%, son los principales exponentes de estos morfogrupos (Figuras 5 y 6).
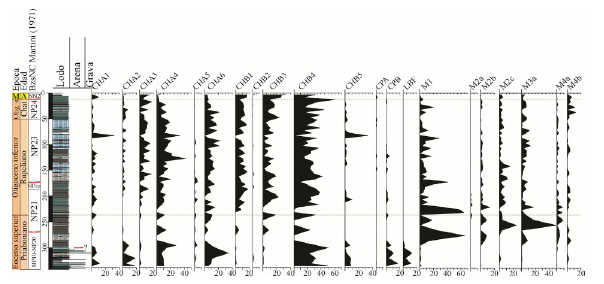
Figura 4 Distribución de los morfogrupos de foraminíferos bentónicos calcáreos y aglutinados en el pozo ANH-SAN JACINTO-1.
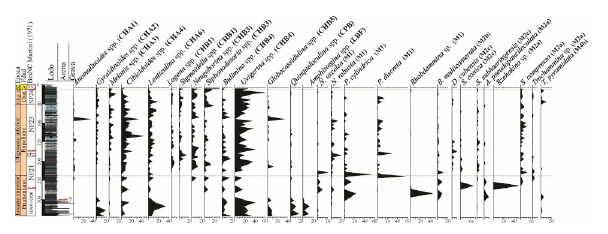
Figura 5 Distribución de los géneros de foraminíferos bentónicos calcáreos y especies de foraminíferos aglutinados más representativos a lo largo del pozo ANH-SAN JACINTO-1.
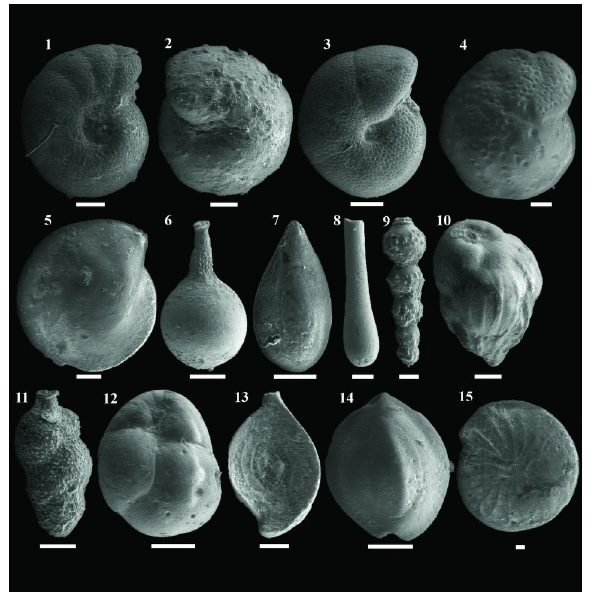
Figura 6 Imágenes de microscopio electrónico de barrido (MEB) de algunas especies que hacen parte de los morfogrupos de foraminíferos bentónicos calcáreos en el pozo ANH-SAN JACINTO-1. 1- Anomalinoides alazanensis (morfogrupo CHA1, vista umbilical), 2- Gyroidinoides dissimilis (CHA2, vista umbilical), 3- Melonispompillioides (CHA3, vista umbilical), 4- Cibicidoides mundulus (CHA4, vista umbilical), 5- Lenticulina melvilli (CHA6, vista umbilical), 6- Lagena sp. (CHB1), 7- Sigmoidella seguenzana (CHB1), 8- Neugeborina longiscata (CHB3), 9- Siphonodosaria jacksonensis (CHB3), 10- Bulimina tuxpamensis (CHB4), 11- Uvigerina rustica (CHB4), 12- Globocassidulina subglobosa (CHB5), 13- Spiroloculina communis (CPA), 14-Quinqueloculina striata (CPB). 15- Amphistegina haueriana (LBF, vista umbilical). La barra blanca equivale a 100 μm.
Los morfogrupos CHA2, que incluyen Cibicorbis spp., Gyroidinoides spp., Hanzawaia spp., Valvulineria spp., se caracterizan por presentar formas cóncavoconvexas y trocoespiral baja y ancha. Por otro lado, CHA6 está compuesto por Amphycorina spp., Astacolus spp., Lenticulina spp., Neolenticulina spp., Planularia spp., Saracenaria spp., Siphonina spp., Spincterules spp., Vaginulinopsis spp., con geometrías lenticulares y planispirales de periferia subaguda/carenada. El morfogrupo CPB, relacionado con foraminíferos bentónicos calcáreos porcelanáceos (CP), tipo quinqueloculina, como Miliolinella spp., Pyrgo spp., Quinqueloculina spp., Spirosigmoilina spp., Triloculinella spp., exhibe formas alargadas a ovadas. Finalmente, LBF (large benthic foraminífera, LBF) como Amphistegina spp. y Operculina spp., destacan en la primera parte del Eoceno superior, entre 342 y 305 m. Estos morfogrupos muestran su máxima abundancia entre los 342 y 305 m, en la parte inferior del Eoceno superior. En este intervalo específico, CHA2 y CHA6 alcanzan sus máximos de abundancia con un 19% a los 327,3 m y un 35% a los 317,1 m respectivamente. Las especies más abundantes de estos morfogrupos son Gyroidinoides spp. (CHA2), con máximos de 18,5%, y Lenticulina spp., con máximos del 33%, mientras que los LBF solo están presentes en este rango estratigráfico y su principal exponente es Amphistegina spp. con abundancias del 13% (Figuras 5 y 6).
Otros morfogrupos presentan mayores abundancias desde inicios del Oligoceno. El morfogrupo CHA3, compuesto por Buccella spp., Melonis spp., Nonion spp., Oridorsalis spp., Osangularia spp., Pullenia spp., y Sphaeroidina spp., caracterizado por formas infladas con periferia biconvexa ampliamente redondeadas, alcanza valores de hasta un 18% a profundidades de 163,7 m y 24 m. El máximo representante es Melonis spp. con abundancias hasta del 14% en el Mioceno (Figuras 5 y 6).
Por otro lado, CHB1, que incluye Chillostomella spp., Ellipsoglandulina spp., Favulina spp., Fissurina spp., Geminiella spp., Glandulina spp., Guttulina spp., Hyalinonetrion spp., Lagena spp., Lagenosolenia spp., Mucronina spp., Oolina spp., Parafissurina spp., Plectofrondicularia spp., Pleurostomella spp., Procerolagena spp., Pyrulina spp., Reussoolina spp. y Sigmoidella spp., se caracteriza por geometrías globulares, ovadas a alargadas y fusiformes biloculares a multiloculares. Este morfogrupo alcanza valores de 18% a los 193,3 m y 16% en el Mioceno. Las máximas abundancias se logran gracias a la presencia de Sigmoidella spp. con máximas hasta de 14% (Figuras 5 y 6).
Por su parte, CHB3, que comprende Bolivina spp., Bulimina spp., Cassidulina spp., Epistominella spp., Evolvocassidulina spp., Fursenkoina spp., Globobulimina spp., Praeglobobulimina spp., Protoglobobuliminas spp., Siphogenerina spp., Siphogenerinoides spp., Siphouvigerina spp., Transversigerina spp. y Uvigerina spp., presenta formas alargadas, rectas a arqueadas uniseriales o planispiral-uniseriada. Este morfogrupo alcanza su máximo de 33% a los 10 m en los depósitos del Mioceno; Siphonodosaria spp. es la especie más abundante a lo largo del pozo con máximos del 30% (Figuras 5 y 6).
Los foraminíferos aglutinados y sus 4 morfogrupos están presentes a lo largo del pozo ANH-SAN JACINTO-1 (Figuras 3 y 5). El morfogrupo M1 (Bathysiphon sp., B. microrhaphidus, Botellina labyrinthica, Nothia sp., N. excelsa, N. robusta, Psammosiphonella spp., P. cylindrica, P. discreta, Rhabdammina sp., R. linearis, y Rizhammina sp.), que comprende principalmente formas tabulares, es el más abundante (Figuras 5 y 7). Sus valores máximos superiores al 50% de la abundancia total se observan entre 282 m y 233 m, con algunos picos notorios en 179 m y 94 m (Figuras 4 y 5).
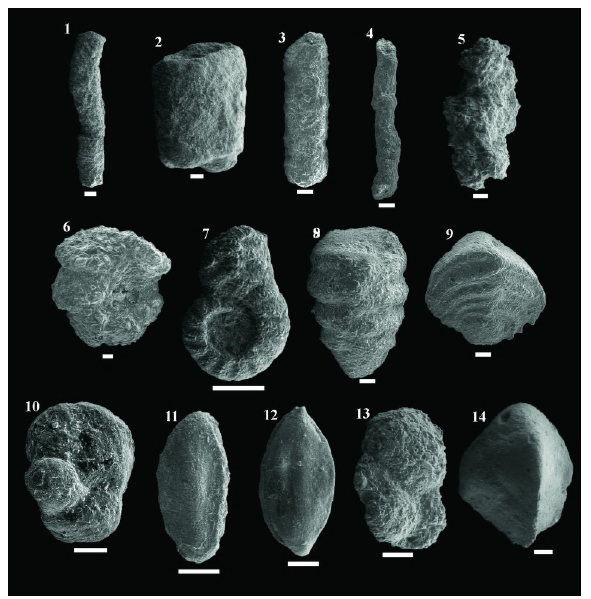
Figura 7 Imágenes de MEB de las especies más comunes de foraminíferos bentónicos aglutinados en el pozo ANH-SAN JACINTO-1. 1- Nothia excelsa (morfogrupo M1), 2- Nothia robusta (M1), 3- Psammosiphonella cylindrica (M1), 4- P. discreta (M1), 5- Rhabdammina sp. (M1), 6- Budashevaella multicamerata (M2b), 7- Duquepsammina cubensis (M2c), 8- Sahulia conica (M2c), 9- Spiroplectinella subhaeringensis (M2c), 10- Ammosphaeroidina pseudopauciloculata (M3a), 11- Rzehakina sp. (M3a), 12- Spirosigmoilinella compressa (M3a), 13- Trochammina sp. (M3a), 14- Tritaxia pyramidata (M4b). La barra blanca equivale a 100 μm.
El morfogrupo M2 (Figuras 5 y 7) se subdivide en 3 subgrupos distintos. M2a representado por Psammosphaera irregularis; M2b que incluye Budashevaella sp., B. multicamerata, B. trinitatensis, Conotrochammina voeringensis, Cribrostomoides subglobosus, C. subglobosus, Lituotuba lituiformis, Recurvoides sp. y R. lamella; y, por otro lado, M2C que comprende Duquepsammina cubensis, Bigenerina nodosaria, Clavulina sp., Goesella cylindrica, Sahulia conica, Spiroplectammina sp., S. navarroana, S. spectabilis, Spiroplectinella sp., S. subhaeringensis, Textularia sp., T. agglutinans, T. isidroensis, Textulariella simplex y Vulvulina pennatula. Estos subgrupos exhiben formas que van desde globulares a trocospirales redondeados, estreptospirales, planoconvexas y quillados elongados. En cuanto a la distribución, M2a alcanza un máximo del 2,5% a una profundidad de 223 m, M2b muestra picos de hasta el 13,3% a 228 m, mientras que M2c es el más abundante del morfogrupo M2, con sus niveles más altos (25%) registrados a los 263 m (Figuras 4 y 5).
El morfogrupo M3 está exclusivamente compuesto por representantes del subgrupo M3a que incluye Ammodiscus sp., A. latus, Ammomarginulina aubertae, Ammosphaeroidina pseudopauciloculata, Eggerella bradyi, Glomospira sp., G. charoides, Paratrochamminoides sp., Rzehakina sp., Spirosigmoilinella sp., S. compressa, Trochamminoides sp., T. variolarius, T. altiformis, Trochamminopsis parvus. Estas formas se caracterizan por ser planispirales aplanadas y estreptosirales. El morfogrupo M3 alcanza su máxima abundancia del 50% a los 263 m (Figuras 4 y 5).
El morfogrupo M4 se divide en 2 subgrupos distintos. M4a está compuesto por Ammoscalaria pseudospiralis, Cyclammina cancellata, Haplophragmoides sp., H. emaciatia, H. excavatus, H. kirki, H. stomatus, H. suborbicularis, H. walteri, Reticulophragmium sp., R. acutidorsatum y R. rotundidorsatum, caracterizadas por formas redondeadas planispirales. Estas formas alcanzan sus valores máximos (>8% de la abundancia total) entre las profundidades 253 m y 228 m (Figuras 4 y 5). Por otro lado, M4b incluye formas elongadas subcilíndricas y elongadas cónicas como Ammobaculites sp., A. agglutinans, A. calcareous, Dorothia goesi, Hormosina velazcoensis, Karreriella bradyi, K. novangliae, K. siphonella, Karrerotextularia crassisepta, Karrerulina conversa, K. horrida, Martinotiella sp., M. communis, Multifidella nodulosa, Nodosinum gaussicum, Pseudonodosinella nodulosa, Reophax sp., R. duplex, R. pilulifer, R. subfusiformis, Subreophax scalaris, Tritaxia barakai, Tritaxia pyramidata y Tritaxis conica. Este subgrupo muestra sus formas con valores más altos hacia el techo del pozo (entre 44 a 15 m), y alcanza máximos de 10% de abundancia a 44 metros de profundidad.
A nivel de oxigenación, los géneros subóxicos y óxicos son más constantes a lo largo del núcleo. En promedio son más abundantes los subóxicos con valores superiores al 60%. Los géneros anóxicos son más constantes y comunes a partir del Oligoceno, y al inicio del Oligoceno alcanzan valores máximos próximos al 60% (Figura 3).
Discusión
Inferencias paleoambientales del pozo ANH-SAN JACINTO-1 en los depósitos del Eoceno superior
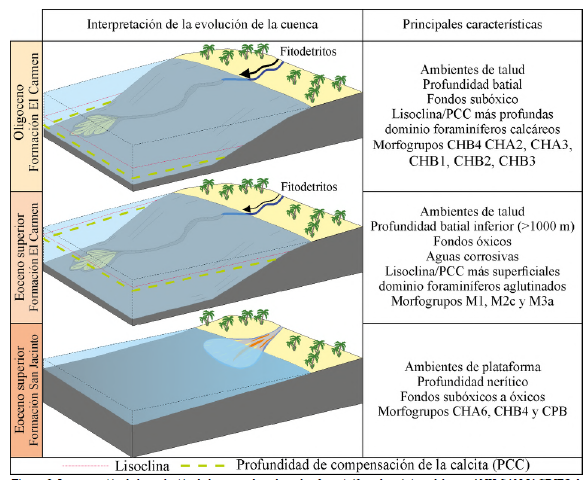
En el Eoceno superior, los foraminíferos bentónicos sugieren 2 condiciones paleoambientales diferentes (Figuras 3 y 8). Desde 342 m hasta aproximadamente los 305 m, predominan los foraminíferos de pared calcárea, representados principalmente por los morfogrupos CHA6, CHB4 y CPB; este último compuesto mayoritariamente por el género Quinqueloculina spp. (Figuras 4 y 5). Estos morfogrupos están asociados con condiciones subóxicas a bien oxigenadas y profundidades en el espectro nerítico (Parada-Ruffinatti, 1996; Katz et al., 2003; Murray, 2006, 2014; Myhre et al., 2017). En este intervalo, la relación P/B muestra valores bajos, lo que se asocia con batimetrías inferiores a 200 m (Tapia et al., 2022). La presencia del morfogrupo LBF (Operculina spp. y Amphistegina spp.) indica igualmente ambientes relativamente someros (neríticos), en aguas tropicales, oligotróficas y de subóxicas a bien oxigenadas (Langer y Hottinger, 2000; Roozpeykar y Moghaddam, 2016; Oron et al., 2018).
Entre el final del Eoceno superior y el inicio del Oligoceno inferior (305 y 230 m) se observan fluctuaciones en la riqueza (S) y diversidad (H'), con un rápido incremento y predominio de los foraminíferos de pared aglutinada, especialmente de especies de los morfogrupos M1, M2c y M3a (Figuras 4, 5 y 7). El Morfogrupo M1 es el más abundante en este intervalo, que consiste en foraminíferos aglutinados epifaunales que se alimentan por suspensión y viven adheridos al sustrato (Murray et al., 2011; Kaminski y Ortiz, 2014). Se relacionan, además, con ambientes en los que existen corrientes de fondo suaves y aguas corrosivas (Kaminski y Gradstein, 2005) y con profundidades superiores a 1000 m entre batial inferior a abisal (Figura 8), con un suministro constante de nutrientes en suspensión (Schröder-Adams, 1986).
Las variaciones en la distribución y composición de los foraminíferos bentónicos podrían indicar cambios batimétricos en la cuenca. A 305 m de profundidad, se observa un cambio litológico donde se estableció el límite entre las formaciones San Jacinto y El Carmen (Arias-Villegas et al., 2023). Esta transición se manifiesta en el cambio de areniscas calcáreas con gradación normal, ocasionalmente intercaladas con calizas, relacionadas con procesos turbidíticos. Estos depósitos suprayacidos por estratos de lodolitas y lodolitas calcáreas presentan laminación plana paralela y bioturbación, que junto con las asociaciones de foraminíferos bentónicos podría asociarse a sistemas marinos profundos.
En la parte central del CPSJ, Duque-Caro et al. (1996) identifican una discordancia entre estas formaciones, con un marcado cambio litológico entre niveles de areniscas bioturbadas de la Formación San Jacinto y estratos de lodolitas de la Formación El Carmen. Salazar-Ortiz et al., 2020 describen un cambio de ambientes someros, desde shoreface a foreshore con deltas dominados por mareas en la Formación San Jacinto, a ambientes desarrollados en profundidades batiales de la Formación El Carmen. Este cambio es interpretado como una transgresión marina que invadió progresivamente el CPSJ desde el norte y el oeste (Mora-Bohórquez et al., 2018; Salazar-Ortiz et al., 2020).
Este aumento en el nivel relativo del mar durante el Eoceno tardío podría estar asociado a un incremento en la subsidencia de las cuencas del Caribe colombiano, tal como fue propuesto por Duque-Caro, 1979 y Mora-Bohórquez et al. (2018). Las variaciones en la composición de los foraminíferos bentónicos, con una disminución de especies calcáreas y un predominio de especies aglutinadas, junto con los cambios litológicos en el pozo, reflejan este cambio paleoambiental. Igualmente, la relación P/B en este intervalo muestra un pico alto, lo que podría indicar ambientes más profundos.
La profundización del CPSJ pudo provocar que los hábitats se ubicaran por debajo de la lisoclina y/o por debajo de la profundidad de compensación de la calcita (PCC). Durante el Eoceno medio y tardío, el enfriamiento global incrementó la cantidad de CO2 disuelto en el océano, lo que aumentó la acidificación en el océano e hizo que la lisoclina y la PCC fueran más someras (Zachos et al., 2001; Rea y Lyle, 2005; Armstrong-McKay et al., 2016). Estas condiciones habrían favorecido la proliferación de foraminíferos aglutinados sobre los calcáreos. Además, el enfriamiento global produjo durante el Eoceno tardío el desarrollo de corrientes corrosivas que se extendieron globalmente (Miller, 1983; Kaminski y Ortiz, 2014; Bordiga et al., 2015). Aunque aún se desconoce cómo la circulación oceánica global influenció el norte de Colombia durante el Paleógeno, es muy posible que la abundancia de foraminíferos bentónicos aglutinados en los depósitos del Eoceno superior se deba a la influencia de estas corrientes. Otra posibilidad, es que estos períodos estuvieran caracterizados por un gran aporte de materia orgánica, lo que habría provocado un pH bajo, y, en consecuencia, una menor disponibilidad de carbonato cálcico disuelto en el océano (Broecker y Peng, 1982; Morse y Mackenzie, 1990). No obstante, el origen de estos sistemas que aportaron altas cantidades de materia orgánica aún debe ser investigado en los registros marinos del Paleógeno del Caribe colombiano.
Inferencias paleoambientales del pozo ANH-SAN JACINTO-1 en los depósitos del Oligoceno-Mioceno inferior
El inicio del Oligoceno se caracteriza por una disminución de foraminíferos aglutinados y un aumento en los foraminíferos de pared calcárea, aunque con pocos ejemplares porcelanáceos (Figura 3). Esta transición puede explicarse por cambios en la profundidad de la lisoclina y/o la PCC. Los foraminíferos bentónicos aglutinados son comunes en áreas con alta disolución de carbonatos, o zonas por debajo de la lisoclina y la PCC, incluso en regiones con alto flujo orgánico, en donde la materia orgánica se puede oxidar y causar disolución de la interfaz agua-sedimento (Murray, 2006; Schröder-Adams y van Rooyen, 2011). Durante la transición del Eoceno al Oligoceno, la PCC experimentó una profundización significativa y repentina a nivel global, lo cual alteró la química oceánica (Rea y Lyle, 2005). Este cambio favoreció la conservación de los foraminíferos de pared calcárea, mientras que las formas aglutinadas disminuyeron. En el Atlántico Norte, se observó que los foraminíferos aglutinados, especialmente los de los morfogrupos M1, M2 y M3, se vieron afectados y disminuyeron en el límite Eoceno/Oligoceno debido a la profundización de la PCC (Kaminski y Ortiz, 2014). Además, el inicio del casquete glaciar Antártico provocó niveles del mar más bajos (Miller et al., 2020), lo que favoreció una PCC más profunda debido a la exposición de las plataformas calcáreas. Esto permitió que se acumulara más carbonato en el océano, lo que favoreció la proliferación de foraminíferos de pared calcárea (Katz et al., 2005). El incremento de la relación P/B en el pozo a partir del Oligoceno (Figura 3) podría respaldar esta interpretación, ya que indica no solo ambientes profundos con una mayor concentración de foraminíferos planctónicos, sino también una mejor preservación de los microfósiles calcáreos (Figura 8). Los nanofósiles calcáreos en el estudio de Arias-Villegas et al. (2023) muestran que estos microfósiles están mejor preservados a partir del Oligoceno, lo que podría reforzar esta interpretación.
No obstante, aunque no se descarta la posibilidad de que un cambio en la profundidad de la cuenca haya podido contribuir a una mejor preservación del carbonato, no se han encontrado evidencias de cambios batimétricos durante este periodo. En realidad, se considera que la cuenca alcanzó un estado de relativa estabilidad tectónica entre finales del Eoceno tardío y el Oligoceno temprano (Mora-Bohórquez et al., 2018). Otra posibilidad para explicar este cambio en la composición de los foraminíferos bentónicos puede estar relacionado a una mejora en la disponibilidad de nutrientes, en donde los foraminíferos calcáreos pueden proliferar y competir con los aglutinados por recursos. Esta competencia puede resultar en una disminución de los foraminíferos aglutinados, especialmente si los calcáreos son más eficientes en la utilización de los nutrientes disponibles (Gooday, 2003; Fiorini y Jaramillo, 2007; Kaminski y Ortiz, 2014).
Desde el inicio del Oligoceno la diversidad aumenta respecto al Eoceno, y se mantiene casi constante hasta el tope del pozo (Figuras 3 y 4). El predominio del morfogrupo CHB4, compuesto por organismos detritívoros, indica ambientes adaptados a un microhábitat infaunal en sedimentos blandos, en zonas donde el fondo es relativamente estable y hay abundantes nutrientes, principalmente detritos orgánicos y bacterias (Koutsoukos y Hart, 1990). De este grupo, algunos géneros oportunistas como Bolivina spp., Bulimina spp., Globobulimina spp., y Uvigerina spp. pueden surgir para aprovechar la disponibilidad de alimento y favorecen un incremento en la diversidad (Lutze y Coulbourn, 1984; Jorissen, 1999; Jorissen et al., 1995, 2007; Gooday, 2003). Desde el comienzo del Oligoceno, y hasta el tope del pozo en el Mioceno inferior, se reconoce el incremento de los morfogrupos CHA2, CHA3, CHB1, CHB2, CHB3, principalmente relacionados con un influjo constante de nutrientes y ambientes mayoritariamente subóxicos a anóxicos (Koutsoukos y Hart, 1990), lo que es soportado por un incremento de los taxones subóxicos como Bulimina spp., Falsoplanulina spp., Gyroidina spp., Gyroidinoides spp. Hansenisca spp. Lenticulina spp. y Uvigerina spp., y anóxicos como Globobulimina spp. y Bolivina spp. (Figura 3). Este cambio en las biocomunidades no se asocia con ninguna variación litológica evidente en la observación macroscópica.
En este intervalo, aunque el morfogrupo M1 sigue siendo el más abundante, las proporciones de los morfogrupos M2c y M3a se asemejan a las de M1. La constancia del morfogrupo M1 probablemente indica condiciones batiales estables. La presencia de los morfogrupos M2c (detritívoros epifaunales, con enrollamiento trocoespiral o formas esféricas que pueden extender sus pseudópodos hasta la superficie del sedimento, incluso, comprenden formas quilladas con modos de enrollamiento mixtos como Spiroplectammina y Spiroplectinella) y M3a (formas aplanados, estreptospirales aplanadas y trocoespirales epifaunales como los trocamínidos) sugiere condiciones mesotróficas, caracterizadas por una alta penetración de oxígeno, un flujo constante de carbono orgánico y una máxima diferenciación de microhábitats, lo que aumenta la diversidad (Frontalini et al., 2014).
El constante aporte de nutrientes fomentó el consumo de oxígeno e hizo que el fondo sedimentario se tornara subóxico. Estas observaciones confirman las interpretaciones de Duque-Caro et al., 1996 y Fiorini y Jaramillo (2007), quienes han propuesto para las secuencias del Oligoceno en la parte central del CPSJ ambientes pobres en oxígeno y con alto aporte de nutrientes. Es posible que el aporte de nutrientes se deba al incremento de fitodetritos, asociados a los sistemas fluvio-deltáicos desarrollados al sur de la cuenca (Mora-Bohórquez et al., 2018; Celis et al., 2023; Pardo-Trujillo et al., 2023). Particularmente el morfogrupo M2c (p. ej., D. cubensis, S. conica) se relaciona con alto aporte de material siliciclástico (Kaminski y Gradstein, 2005; Murray et al., 2011; Kaminski y Ortiz, 2014), por lo que su constancia en el pozo puede estar relacionado con el origen continental de los fitodetritos (Figura 8).
Hacia la parte superior del Oligoceno y en el Mioceno se registra un importante incremento del morfogrupo aglutinado M4b (T. pyramidata) y el calcáreo CHB3 (Siphonodosaria spp.) y CHB4 (Uvigerina spp.). De acuerdo con Kaminski y Gradstein (2005), Kaminski y Ortiz (2014) y Koutsoukos y Hart (1990), estos morfogrupos se asocian a taxones infaunales profundos, característicos de ambientes pobres en oxígeno y regiones con eutrofismo, asociados a altos niveles de flujo orgánico. Una explicación para este elevado aporte de nutrientes puede estar en las altas tasas de sedimentación y el incremento del aporte de sedimentos hacia las cuencas que se desarrollaron a finales del Oligoceno y comienzos del Mioceno temprano (Mora-Bohórquez et al., 2018).
Implicaciones paleogeográficas del pozo ANH-SAN JACINTO-1
Las características paleoambientales reveladas por los foraminíferos bentónicos del pozo ANH-SAN JACINTO-1 coinciden con la propuesta de ambientes de depósito de Petters y Sarmiento (1956) en la región de El Carmen de Bolívar. Para ellos, la biozona Bulimina jacksonensis del Eoceno superior contiene diferentes géneros de los denominados grandes foraminíferos bentónicos (morfogrupo LBF), entre ellos Amphistegina spp., que es abundante en la base del pozo. Petters y Sarmiento (1956) indican que el Eoceno superior se asocia con "depósitos arenosos de aguas poco profundas", lo que podría concordar con los ambientes más superficiales, neríticos, observados entre la base del pozo a 342 m y aproximadamente los 305 m de profundidad.
En contraste, las biozonas del Oligoceno y Mioceno más inferior, posiblemente las biozonas Cibicidoides perlucidus y Globigerina dissimilis, indican ambientes que abarcan desde la plataforma hasta el talud. Según estos autores, en la región de El Carmen de Bolívar se observa un predominio de formas pelágicas, asociaciones en el rango nerítico externo a batial, lo que sugiere una cuenca profunda. Es importante destacar que Petters y Sarmiento (1956) no observan el cambio entre las formas calcáreas y aglutinadas en el Eoceno superior que se identificaron en el pozo ANH-SAN JACINTO-1.
Por otra parte, Fiorini y Jaramillo (2007) también estudiaron los foraminíferos bentónicos de una sucesión sedimentaria de la Formación El Carmen en la región de El Carmen de Bolívar. Sus resultados indican asociaciones batiales entre la zona batial superior a medio, especialmente con fondos reducidos en oxígeno y ambientes con alto aporte de nutrientes. Aunque el trabajo de estos autores no proporciona un marco temporal que permita comparar sucesiones sedimentarias directamente, sus observaciones coinciden a nivel general con nuestros hallazgos en los niveles del Oligoceno y Mioceno inferior del pozo ANH-SAN JACINTO-1. No obstante, a diferencia de las asociaciones en el pozo, que muestran un predominio de formas calcáreas en el Oligoceno y Mioceno, las muestras de Fiorini y Jaramillo (2007) presentan una mayor constancia y dominancia de formas aglutinadas, lo que sugiere periodos con una menor disponibilidad de carbonato cálcico disuelto, y ambientes ricos en materia orgánica y con pH bajo. El contraste en las abundancias de foraminíferos calcáreos y aglutinados entre el trabajo de Fiorini y Jaramillo (2007) y nuestros resultados tal vez puede deberse al dinamismo oceanográfico que caracterizó la cuenca durante su sedimentación. Sin embargo, la falta de resolución temporal en el estudio de Fiorini y Jaramillo (2007) limita la posibilidad de realizar comparaciones más precisas.
Conclusiones
El estudio de los foraminíferos bentónicos del pozo ANH-SAN JACINTO-1 revela dos paleoambientes distintos durante el Eoceno tardío, que se relacionan con el cambio litológico entre las formaciones San Jacinto y El Carmen. El primero se caracteriza por el dominio de los foraminíferos bentónicos de pared calcárea (principalmente los morfogrupos CHA6, CHB4 y CPB), relacionados a ambientes de plataforma en profundidades neríticas. En el segundo, dominan los foraminíferos aglutinados de ambientes de talud. La disminución de foraminíferos calcáreos se interpreta como resultado de cambios batimétricos que pudieron hacer que el fondo sedimentario se posicionara por debajo de la lisoclina y/o de la PCC. Durante el Oligoceno, vuelven a ser más abundantes los foraminíferos de pared calcárea (p. ej. CHA2, CHA3, CHB1, CHB2, CHB3), lo cual indica posiblemente una profundización en la PCC y/o lisoclina. Además, es notable un incremento en la diversidad de los foraminíferos bentónicos y en la abundancia de especies subóxicas. Los morfogrupos aglutinados M2c y M3a indican un constante influjo orgánico, mientras que el dominio del morfogrupo calcáreo CHB4 sugiere un alto aporte de nutrientes y bajos niveles de oxígeno en el fondo marino. Hacia la parte alta del Oligoceno superior y el Mioceno inferior, el incremento de especímenes del morfogrupo aglutinado M4b y de los morfogrupos calcáreos CHB3 y CHB4 indica anoxia y un aumento en el influjo de nutrientes en los fondos marinos.
Los cambios observados en las comunidades bentónicas a lo largo del pozo ANH-SAN JACINTO-1 pueden correlacionarse con eventos de magnitud global. Durante el Eoceno tardío, el enfriamiento global provocó un descenso en la profundidad de la lisoclina y la PCC, que afectó a las comunidades bentónicas calcáreas. En este período, predominaron los foraminíferos aglutinados, especialmente el morfogrupo M1, asociado con aguas frías y corrosivas. En contraste, durante el Oligoceno temprano, se produjo una profundización de la lisoclina y/o la PCC a nivel global, probablemente debido a la disminución del nivel del mar tras el establecimiento de un casquete de hielo permanente en la Antártida. Esta disminución del nivel del mar aumentó la exposición de las plataformas, elevó la concentración de carbonato de calcio en el océano y favoreció el desarrollo de foraminíferos bentónicos calcáreos, como se observa en el pozo desde el Oligoceno temprano.

