










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.31 no.1 Bogotá Jan./June 2005
Diversidad de tres gremios de hormigas en cafetales de sombra,
de sol y bosques de Risaralda
Diversity of three ant guilds in shade and sun coffee plantations and forests of Risaralda
LEONARDO RIVERA1, IMGE ARMBRECHT1-2
1 Autor para correspondencia: Universidad del Valle, Departamento de Biología, Ciudad Universitaria Meléndez Calle 13 No 100-00, tel: 321 21 00 ext. 2570. A. A. 25360, Cali, Colombia. E-mail: endimion_x@yahoo.com
2 University of Michigan, School of Natural Resources and Environment. Daña Building, 430 East University, Ann Arbor, MI, 48109-1115. E-mail: inge@univalle.edu.co
Resumen. Durante 2001-2002 se realizó un estudio de la mirmecofauna (Hymenoptera: Formicidae) en cafetales del municipio de Apía (Risaralda), para evaluar el impacto que las técnicas modernas de cultivo de café tienen sobre la biodiversidad asociada. Se seleccionaron 12 fincas clasificadas en tres sistemas de manejo de sombra de café (tratamientos) y un control compuesto por parches de bosque. Se ubicaron 20 parcelas de 1 m2 por finca en las cuales se realizó colecta manual de hormigas en hojarasca y con mini-wiñkler además de muestreo de hormigas en el cafeto más cercano a cada parcela. Por último se midieron variables bióticas y abióticas en el área. Las hormigas registradas se agruparon en gremios seleccionados según la clasificación de Delabie et al. para Sur América. Se registraron 22.983 individuos pertenecientes a 115 morfoespecies donde los cafetales orgánicos de sombra poligenérica demostraron contener la riqueza mayor. Las hormigas se agruparon en tres gremios: crípticas de hojarasca, abundantes y de cafetos. Las especies crípticas fueron las más sensibles a los cambios en el manejo, mostrándose muy reducidas en cafetales de sol, mientras que las hormigas abundantes y de cafeto no fueron afectadas en la misma manera. Se encontró que la presencia de este gremio está afectada positivamente por la cobertura vegetal y negativamente por la temperatura del suelo. Se concluye que un manejo agrícola ambientalmente amigable como es el cafetal orgánico de sombra diversa, se constituye en una matriz favorable para la biodiversidad de hormigas propias de parches de bosque en el paisaje rural cafetero.
Palabras clave: Manejo del café. Mirmecofauna. Wínkler. Agroecosistema. Conservación.
Summary. During 2001-2002, a study on the myrmecofauna (Hymenoptera: Formicidae) in coffee plantations was conducted int the Apía municipality (Risaralda) in order to evalúate the impact of modern agricultural practices on the associated biodiversity. TWelve coffee farms were selected and classified into three shade managements (treatments) and a control consisting of forest patches. TWenty Im2 -plots were located on each farm and at each plot ants were collected manually and with a mini-Winkler from the leaf litter. Ants were also sampled from the nearest coffee bush. Finally, biotic and abiotic variables in the área were measured. The ants were grouped into guilds according to the classification of Delabie et al. for Soulh América. A total of 22.983 individuáis belonging to 115 morphospecies was recorded. Polygeneric shaded coffee (organic managed coffee) had the greatest species richness. Ants were grouped into three guilds: cryptic litter-dwelling, abundant and coffee bush. The cryptic guild was the most sensitive to changes in coffe management, being very reduced in sun plantations, while abundant ants and ants on coffee bushes were not affected in the same way. The presence of the cryptic guild was positively affected by vegetation cover and negatively by soil temperature. We concluded that an environmentally friendly agricultural management, like organic coffee with diverse shade, is a matrix that favors the diversity of ants from forest patches in the rural coffee landscape.
Key words: Coffee management. Myrmecofauna. Winkler. Agroecosystem. Conservation.
Introducción
La elevada tasa con la que se expande la frontera agrícola y la intensificación de la agricultura, ha reducido de forma significativa los hábitats naturales disponibles para organismos silvestres (Paoletti et al. 1992; Swift et al. 1996); aún así es posible hallar una cantidad apreciable de diversidad biológica en pequeños parches de bosque y sistemas de agricultura tradicional aledaños, los cuales proporcionan refugio a los organismos propios del bosque.
Sistemas agroforestales como aquellos constituidos por los cafetales tradicionales, han sido foco de atención científica dada su elevada biodiversidad y su complejidad florística (Perfecto et al. 1996). Este sistema, con diversas capas de sombra y árboles frutales, ha sido reemplazado paulatinamente por varias formas moderadas de manejo que comienzan con una regularizado)! y monotonización del suelo, la eliminación de muchas de las intercosechas, continuando con la eliminación por entero de la sombra. El principal propósito del cambio de manejo descrito es hacer frente tanto a problemas fitosanitarios como buscar un incremento en los rendimientos (lbarra-riuñez 1990). Estos cambios, que se realizan en las plantaciones y en las prácticas agrícolas, no tienen en cuenta que la utilización intensiva de plaguicidas en las modernas plantaciones de café sin sombra o monocultivos, incide de forma directa e indiscriminada sobre la fauna del agroecosistema cafetero (Perfecto y Armbrecht 2003). De la misma forma, al disminuir la complejidad estructural del cultivo cambia el microclima de los cafetales, disminuyendo los nichos ecológicos disponibles para muchos grupos de animales (macro y microfauna) (Moguel y Toledo 1999).
Se ha documentado que las modificaciones de los cafetales han producido cambios significativos tanto en términos de la biodiversidad planeada (ej: las cosechas específicamente planeadas por los granjeros) como de la biodiversidad asociada,incluyendo insectos, otras plantas y animales que están asociadas con las cosechas y el habitat que ellas crean (Perfecto et al. 1996). La fauna asociada es un componente muy importante, no solo en los sistemas forestales sino en los agrícolas, incluyendo los cafetales, por tanto es preocupante saber que con la reducción de la sombra también se reduce las fauna potencialmente benéfica para el agricultor (Beer et al. 1998).
Estudios en Costa Rica y México han encontrado que la diversidad de la comunidad de hormigas asociadas a cafetales disminuye conforme aumenta la intensidad del manejo del cultivo (Perfecto y Vandenneer 1994; Perfecto et al. 1996; 1997; Armbrecht y Perfecto 2003). Diversos trabajos demuestran que las hormigas (Hymenoptera: Formicidae) son un grupo sensible que pueden ser usadas como bioindicadores en ambientes terrestres. Las hormigas presentan una distribución geográfica amplia, son localmente abundantes, funcionalmente importantes en todos los niveles tróficos, fáciles de muestrear y separar en morfoespecíes. Además son susceptibles a cambios climáticos y también permiten una clasificación en grupos funcionales debido a su diversidad, de tal manera que pueden relacionarse con la de otros componentes bióticos del área estudiada. Por otro lado, cuando la estructura de la comunidad de hormigas cambia por disturbios ésta puede reflejar cambios en otros grupos de invertebrados (Majer 1993; Andersen 1997; Osborn eí al. 1998).
En este estudio se pretende evaluar el impacto producido por el cambio en el manejo de los cafetales tradicionales y la eliminación de la sombra arbórea, sobre el comportamiento ecológico de tres gremios de hormigas.
Materiales y Métodos
Área de estudio
El municipio de Apía, departamento de Risaralda, es por tradición un pueblo cafetero que ocupa el lugar 68 a nivel nacional y el octavo en el departamento de Risaralda de acuerdo con el escalafón de producción anual de café. La reciente constitución de la Sociedad Agropecuaria Orgánica Tatatná, ha hecho de este municipio uno de los más pujantes en cuanto a cultivo y exportación de café. Sus actividades antrópicas en la microcuenca del río Apía se centran principalmente en la exportación de una caficultura que combina manejos tradicionales con manejos convencionales (tecnificados) del cultivo del café (Boletín Sociedad Agropecuaria Tatamá 2002). Además, es posible encontrar algunos parches de bosque (relictos) que permiten hacer comparaciones entre la rnirmecofauna (fauna de hormigas) presente en estos y en los cafetales.
Tratamientos
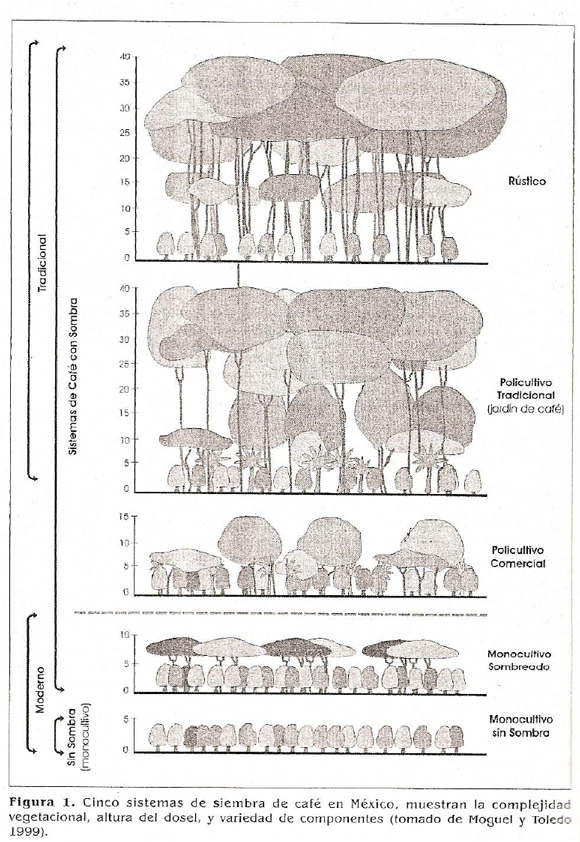
Se seleccionaron tomando como base conceptual los cinco sistemas de manejo del café utilizado en México por Moguel y Toledo (1999) (Fig.1), que tienen en cuenta el nivel de manejo, complejidad vegetal y estructural.
Se ubicaron 12 fincas de la zona clasificadas en cuatro manejos (descripción detallada en Armbrecht, en prensa), correspondientes a los tres cultivos representados en la parte inferior de la figura 1, además del bosque, que no aparece en dicho esquema. Los manejos fueron los siguientes en orden de intensificación creciente: (1) Bosque (Bos), consistente de tres parches de bosque cercanos al estudio y que constituyeron el control positivo asumiendo que representan la vegetación natural de la zona; (2) Cafetal orgánico con sombra poligenérica (PS), es decir, con árboles de sombra de diferentes géneros (tres fincas); (3) Cafetal con sombra monogenérica (MS), cafetales con un solo género de árbol de sombra: Inga spp. ó Cordia alliodora (tres fincas) y (4) Cafetal sin sombra de árboles o monocultivo de sol (Sol). En total fueron tres tratamientos (tres fincas por tratamiento) y un control (tres bosques).
En cada una de las fincas se eligió al azar un transecto sobre el cual se ubicaron 20 unidades de muestreo, diez en temporada seca (octubre-noviembre 2001) y diez en temporada de lluvia (enero-febrero 2002), realizándose un total de 240 unidades de muestreo (UM).
Muestreo
Cada UM consistió de una parcela de un metro cuadrado (utilizando un cuadrante de piola sintética o naylon de 1 x 1 m), en la que se realizó el siguiente procedimiento; (1) captura manual de hormigas forrajeando en el área: 20 minutos de colecta y registro de número de obreras; (2) examen en todas las ramitas, troncos y hojarasca para detección de nidos; (3) cernido de toda la hojarasca presente en la UM; (4) toma de temperatura en la superficie de la hojarasca mediante termómetro de suelo; (5) medición del perfil de hojarasca introduciendo una regla, hasta la tierra con lo cual se obtenía la medida del espesor en cm (promedio de 5 puntos tomados al azar); (6) medición de la cobertura del dosel mediante densiómetro esférico de vegetación (Forestry supplies, Inc, modelo C); (7) toma de pH del suelo a 10 cm de profundidad utilizando un Soil Tester Kelway (Kel Instrumente Co., Inc); (8) humedad del suelo a 10 cm de profundidad medida registrada (con el instrumento del punto 7).
El material de hojarasca cernido (ver procedimiento 3) se dispuso en sacos mini-Winkler durante 48 h para la extracción de las hormigas presentes en la hojarasca colectada de acuerdo con el protocolo de ALL (Ante of Leaf Litter, Agosti et al. 2000). Esto suma 11.520 horas de extracción en las 240 UM. Posteriormente se examinó el arbusto de café más cercano a la parcela UM y se registraron las hormigas encontradas durante 15 minutos de muestreo visual. Además, se revisó en corteza, follaje y restos de soca en busca de individuos o nidos. Si no se detectaba actividad en ese tiempo, se agitaba el cafeto y se observaba de nuevo. El esfuerzo de muestreo directo se calculó en un total 120 h repartidas en 30 h para cada tratamiento en las 12 fincas seleccionadas.
Trabajo de laboratorio
Las hormigas se preservaron en viales individuales con alcohol al 70% y se rotularon. Con estas se montó una colección de referencia determinándolas hasta el nivel taxonómico de género con las claves de tlólldobler y Wilson (1990). Bolton (1994). A nivel de especie se utilizaron algunas claves disponibles y la asesoría del taxónomo Fernando Fernández del Instituto Alexander von Humboldt (Villa de Leyva, Boyacá). Se ratificó la identidad de las especies de hormigas por comparación con la colección de referencia del Museo de Entomología de la Universidad del Valle (Cali). Otras especies se identificaron por varios taxónomos en el exterior.
Análisis de datos
Se registró la información en una base de datos elaborada con el programa EXCEL 97 (2002) donde se introdujeron las siguientes variables para cada muestra de hormigas: nombre de la finca donde se colectó, determinación taxonómica, número de individuos de cada morfoespecie por unidad muestral y total de hormigas por parcela. En los análisis se tuvieron en cuenta tanto el número de obreras de cada especie de hormiga colectada (abundancia), como la incidencia de las mismas en cada tratamiento.
Trabajo con los gremios
Selección. Para estudiar la ecología global de la comunidad se requiere de la identificación de grupos funcionales que trasciendan los límites taxonómicos y biogeográficos en respuesta al estrés y a la perturbación (Andersen 2000), por esto se agruparon las especies de hormigas en tres grupos funcionales con base parcialmente en la clasificación que hacen Delabie et al. (2000) de las hormigas de hojarasca en selva lluviosa de Sur América. Esta clasificación está basada en características de forrajeo, alimentación y anidamiento de la mirmecofauna de la zona. Con la base de datos elaborada para todas las fincas se seleccionaron las especies de cada gremio quedando agrupadas de la siguiente forma: especies de hormigas en las plantas de café, especies de hormigas crípticas de hojarasca, y especies de hormigas abundantes (dominantes) de hojarasca.
Las especies de cada gremio se seleccionaron principalmente según su identidad, su abundancia total, las observaciones registradas durante y después de la captura (sitios de anidamiento y forrajeo). Posteriormente se tuvieron en cuenta las descripciones de comportamiento que hacen tlólldobler y Wilson (1990).
Comportamiento de los gremios en los tratamientos. Se analizó la sensibilidad de los gremios generando diferentes curvas de acumulación de especies usando el programa ESTIMATES 6 (Colwell 1997). Basándose en datos aleatorizados, las curvas de acumulación permiten comparar el crecimiento de las especies por tratamiento y son un esquema del número acumulativo de especies descubiertas S (n), dentro de un área definida, como función del esfuerzo esperado para encontrar éstas (Colwell y Coddington 1994). Se utilizaron únicamente estimadores no paramétricos basados en la incidencia (presencia o ausencia) de las especies. Las hormigas son sociales y por lo tanto se hallan fuertemente agregadas, lo que hace preferible el empleo de este tipo de datos más que los de abundancia. El análisis con hormigas de cafetos no se realizó en los bosques puesto que el sotobosque de éstos no está compuesto de cafetos.
índices de diversidad. Dos índices fueron calculados con el programa SPDIVERS. BAS (Ludwing y Keynolds 1988): el índice de diversidad de Shannon (IT) el cual es el grado promedio de incertidumbre para predecir a qué especies pertenece un individuo cogido al azar de una colección de S especies y n individuos, y el de Simpson [Vi que indica la probabilidad de que dos individuos extraídos al azar de una población pertenezcan a la misma especie. Estos índices se calcularon para cada gremio en cada finca para poner a prueba la hipótesis de que existe diferencia en los índices de los gremios. El gremio cafetos no fue incluido para bosque puesto que las plantas del sotobosque no eran comparables con los cafetos.
Variables bióticas y abióticas. Se realizó un análisis de correlación con el programa STATISTCA 6 entre cada una de las variables bióticas y abióticas tomadas en cada UM y la riqueza de especies (número de especies) de cada uno de los tres gremios seleccionados.
Comparación de los tratamientos. Para comprobar si había diferencias entre fincas o entre tratamientos con el gremio crípticas de hojarasca se usaron pruebas de Kruskal-Wallis para datos no paramétricos.
Resultados
Se registraron 22.983 individuos de hormigas, pertenecientes a 115 morfoespecies, las cuales fueron agrupadas en 44 géneros de ocho subfamilias existentes en el neotrópico (Bolton 1994), 56 pertenecientes a la subfamilia Myrmicinae, 27 dentro de Ponerinae, 14 en Formicinae, ocho en Dolichoderinae, seis en Pseudomyr-mecinae, tres dentro de Ecitoninae y una sola especie clasificada dentro de la subfamilia Cerapachyinae. Una especie de hormiga cazadora (Ponerinae) perteneciente al género Simopelta resultó nueva para la ciencia y fue encontrada en uno de los cultivos poligenéricos de manejo orgánico (finca la Playita, Gustavo Zabala com. pers. octubre 2003). Adicionalmente, de acuerdo con el taxónomo John Lattke otra especie de hormiga cazadora, Gnampto-genys, probablemente es también nueva para la ciencia y proviene de uno de los bosques. Las especies más abundantes fueron las myrmicinas con 63,6% del total de capturas, y las formicinas con el 22%. A nivel de géneros, Pheidole, Hypoponera, Pachycondyla, Solenopsis, y Gnampto-genys estuvieron mejor representadas con 19, 8, 8, 7 y 7 especies respectivamente. En total, la abundancia de hormigas halladas en la hojarasca fue mayor que aquella en cafetos (16.504 y 6.479 individuos, respectivamente).
En términos de riqueza los cultivos poligenéricos albergaron el porcentaje mayor de hormigas en el estudio (Pig. 2). Comparando estos resultados con los de los bosques, donde se registraron el 61,5% de las especies, se observa que los cafetales de sombra poligenérica representaron el mejor acumulado de mirmecofauna del estudio. Un total de 27 especies estuvieron presentes (compartidas) en los tres tratamientos de manejo de cafetales y en los bosques. Los cafetales de sombra monogenerica y los de sombra poligenérica con 47 especies compartidas, fueron los tratamientos con más semejanzas en cuanto a composición. En términos de hojarasca, los bosques presentaron el mayor número de especies exclusivas (Tabla 1), seguidos por los cafetales de sombra, y el valor más bajo fue para ios cafetales de sol. A nivel de estratos verticales, 41 especies de hormigas se colectaron al menos una vez en hojarasca y 34 especies una vez en cafetos (hasta 2 m de altura).
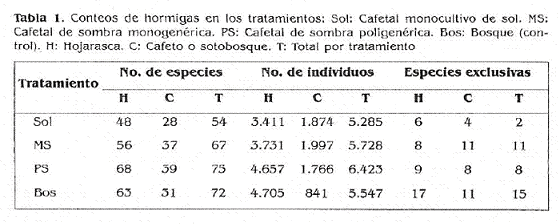
El cultivo pol¡genérico es el tratamiento con mayor número de especies (68) y con la mayor abundancia de individuos (6.423), el tratamiento Sol con 54 especies comparte el mayor número de sus especies con los demás tratamientos por tener solo dos especies excusivas. Algunas especies se observan como típicas de un tipo de habitat. Por ejemplo, Tetramorium simillimum, Wasmannia awopunctata, Atta cephalotes y Gnamptogenys gr striatüla son muy frecuentes en los Soles. Lahídus coecus, Odontomachus erytrocephalus y Gnamptogenys ty fueron obtenidas en mayor proporción en cultivos monogenéricos. En cuanto a los cafetales de sombra poligenérica, se encuentran diferentes especies que son abundantes en estos tratamientos, pero que son compartidas con los bosques tales como Octostruma cf síenocarpa, O. balzani, Pseudomyrmex oculatus, P. tenuissimus y Eciton burchelli; mientras que otras como Paratrechina steinheili y Gnampfogenys minuta fueron compartidas con los cafetales de sombra monogenérica. En los bosques se presentó el mayor número de especies típicas siendo notorias por su alta abundancia las especies Crematogaster sp. 3 y Gnamptogenys bisulca.
Análisis de los gremios
Un total de 51 individuos se agruparon como especies de hormigas crípticas de hojarasca (se presentan generalmente en densidades bajas), 22 abundantes de hojarasca (aquellas en cantidades mayores o iguales a 110 individuos) y 24 como especies de hormigas en las plantas de café. Las especies de hojarasca presentaron altas diferencias en su abundancia, el gremio de las especies abundantes exhiben niveles altos en casi todos los tratamientos, mientras que las especies del grupo crípticas de hojarasca son relativamente pocas por finca.
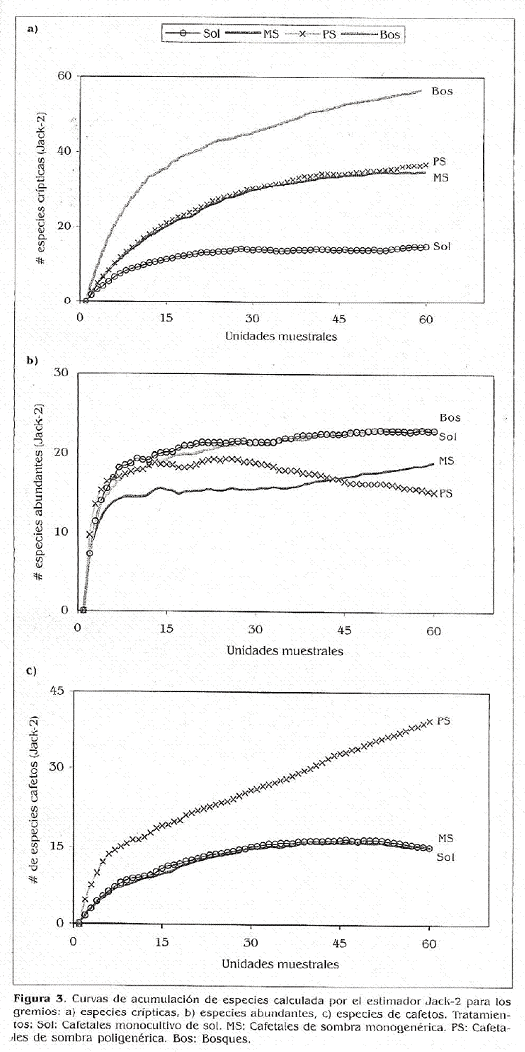
Acumulación de especies. Se trabajó con el indicador Jacknife 2, el cual se basa en el número de únicos y duplicados y en el número de sitios muestreados, mostrando una alta tendencia a estabilizarse en cuanto al incremento de especies. Este estimador fue el que presentó curvas más asintóticas en todos ios tratamientos.
El gremio de las especies crípticas muestra tendencia a decrecer en los tratamientos sol y cafetal de sombra monogenérica (MS), mientras que se incrementa en los tratamientos con sombra poligenérica (PS), alcanzando su valor máximo en los bosques (Fig. 3a). Las hormigas que se clasificaron en el conjunto de las abundantes se ven acumuladas tanto en las fincas de café sin sombra (Sol) como en los bosques (Fig. 3b); así mismo, la gráfica no manifiesta diferencias entre los tratamientos MS y PS. Las hormigas que habitan los cafetos tienen una fuerte tendencia a ser más diversas en los tratamientos de sombra poligenérica (Fig. 3c).
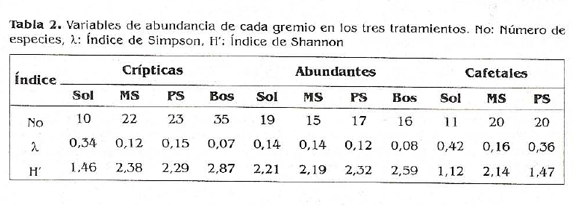
Comparación de la diversidad de los gremios entre fincas. Según el índice de diversidad de Shannon (H'), el gremio crípticas de hojarasca en el tratamiento control Bosque, es el más diverso, y el grupo de hormigas de cafetales en los cultivos de sol el menos diverso (Tabla 2). La respuesta del índice de Simpson en el gremio crípticas fue muy alta para los bosques pues entre más bajo el valor de la probabilidad, más diverso es el habitat.
Correlación con variables bióticas y abióticas. El gremio de hormigas crípticas de hojarasca es significativamente afectado por dos de las variables tomadas, cobertura de dosel y temperatura del suelo (Fig. 4). Los datos correspondientes a las variables en cada finca representan la media de todas la unidades de muestreo realizadas en el transecto (20 UM).
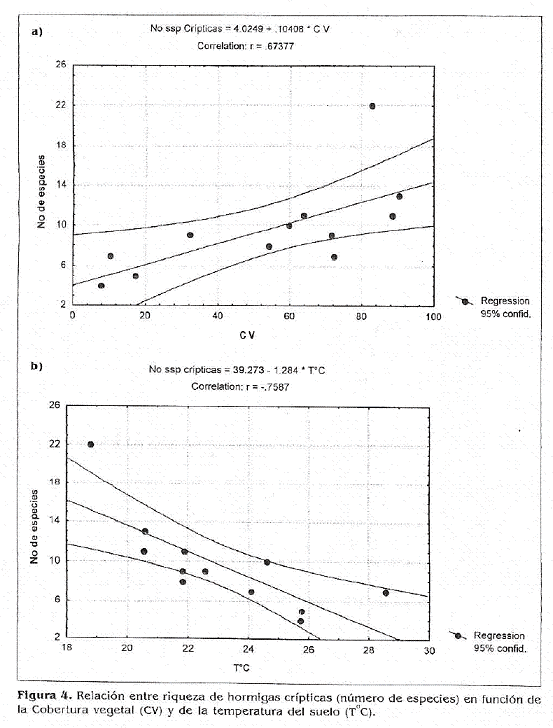
Cobertura del dosel. Esta variable presentó una relación positiva con la riqueza de hormigas crípticas de hojarasca (r= 0,67377; g.l 10; p< 0,02) (Fig. 4a). Los porcentajes más altos de cobertura se registraron en cafetales de sombra poligenérica y bosqties, y progresivamente fue más bajo en los monocultivos (Soles) (Tabla 3).
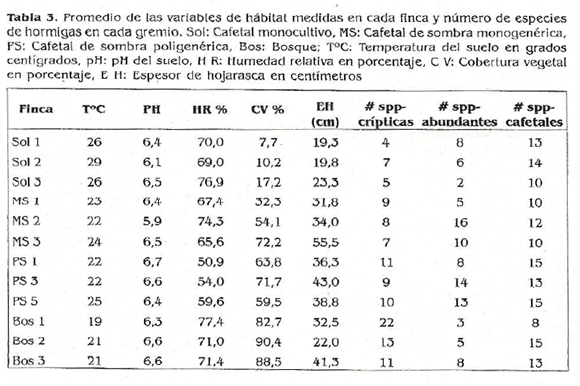
Temperatura del suelo. Se encontró una relación significativa negativa entre esta variable y la riqueza (r = -0,7587; g.l 10; p<< 0,01) (Fig. 4b). La temperatura más baja va desde 17°C registrada en Monteverde (Bosl), hasta la más alta de 36°C en los cafetales sin sombra La Estrella y La María (Sol 2 y Sol 3).
Comparación de las especies crípticas entre fincas y tratamientos. El análisis Kruskal Wallis mostró diferencias significativas entre las diferentes fincas (Sol 1, Sol 2, Sol 3,..., Bos 3) en cuanto al número de especies (riqueza) (II = 51,67; g.l. 11; p< 0,001); y diferencias altamente significativas al ser analizada la abundancia de individuos del gremio (H = 41,09; g.l. 11; p< 0,001). Igualmente, los tratamientos (es decir, los manejos: Sol, MS, PS y Bos) presentaron diferencias significativas tanto en términos de la riqueza (Kruskal Wallis, H = 2,69; g.l. 3; p< 0,00001) como de la abundancia (Kruskal Wallis, H = 18,27; g.l. 3; p< 0,00001).
Discusión
Las 115 especies de hormigas, registradas en este estudio, representan aproximadamente el 15% de las 750 especies señaladas para el país. Este porcentaje, sumado al hallazgo de dos especies nuevas para la ciencia, evidencian la importancia biológica del paisEije cafetero colombiano como fuente de fauna asociada de artrópodos.
Trabajar con riqueza (número de especies) es más apropiado que la sola abundancia para evaluar la sensibilidad de los tratamientos (Qotelli y Colwell 2001). Al analizar la diversidad en términos de riqueza, obtenida en los tratamientos, se observa que los cafetales de sombra poligenérica concentran una mayor cantidad de especies que los parches de bosque muestreados. Estos resultados difieren de lo que se espera ocurra con las hormigas ya que éstas tienden a disminuir en diversidad a medida que aumenta la intensidad del manejo (Perfecto y Vanderrneer 1994). En otras palabras, debería haber una más alta diversidad en bosques que en cualquier
cafetal. Se proponen tres explicaciones para este resultado: (1) Los estratos muestreados (suelo hasta 2 m) no hicieron justicia a los bosques, en donde parte de su fauna puede estar concentrada en capas altas de dosel, y por tanto no fueron detectadas en este estudio. Si el objetivo hubiera sido un inventario exhaustivo de cada habitat habría sido necesario abarcar la mayor heterogeneidad posible del sitio (Ramos et al. 2002), pero debido a que el presente estudio trataba de comparar los cuatro habitáis con exactamente el mismo esfuerzo de muestreo, cabe la posibilidad de submuestreo en el dosel y subcapas arbustivas de los bosques. (2) Los bosques seleccionados no son tan diversos como se esperaba, debido a que son parches pequeños de bosque secundario relativamente joven, que anteriormente fueron cafetales y que son preservados porque significan una fuente natural de agua. (3) Los cafetales de sombra, especialmente los de sombra poligenérica, tienen tanta diversidad como el bosque. Los datos de este estudio sugieren fuertemente que éste puede ser el caso, y, aunque también la evidencia muestra que los bosques preservan especies únicas o exclusivas mucho más que cualquiera de los cafetales, este estudio reitera una vez más la inmensa importancia que tienen cafetales manejados con sombra diversa para la conservación de la diversidad biológica. Una matriz de alta calidad como los cafetales poligenéricos de manejo orgánico puede convertirse en un vehículo que favorece la dinámica de metapoblaciones (colonización o recolonización) entre los relictos de bosque que existen en la zona cafetera Colombiana.
La identidad de las especies evidencia también la importancia de los bosques. Los parches de bosque muestreados contribuyeron con el mayor número de especies exclusivas, 17 en hojarasca y 11 en sotobosque (Tabla 1). Aunque no presentaron una riqueza alta como se esperaba, hay que tener en cuenta que, para propósitos de conservación, se debe integrar información sobre calidad o vulnerabilidad biológica, que podría ser más importante que cantidad (Ramos et al. 2002). En este aspecto, siguen siendo un banco de especies que posiblemente no se adapten a otro tipo de condiciones como las presentes en los cultivos sin sombra y por tanto la conservación de los bosques debe estar en la primera prioridad y ser concomitante con un apropiado manejo de la matriz agrícola.
Se consideró que los gremios seleccionados son los que mejor representan los estratos horizontales y verticales del lugar muestreado, pero hay que tener en cuenta que esta clasificación está hecha para selva lluviosa (Delabie eí al. 2000), en la cual es posible que existan más microhábitats disponibles y por lo tanto, se pueden hacer más tipos de agrupaciones; aun así, la clasificación se puede ajustar a hormigas de toda la región neotropical.
Hay dos variables que presentan un alto porcentaje de correlación. La variable abiótica temperatura del suelo es la que mejor explica la riqueza de especies, presentándose una relación inversa, donde a medida que aumenta la temperatura del sitio disminuye el número de especies capturadas (esto solo teniendo en cuenta las especies que se logran capturar con Winkler y que además son diurnas). La variable biótica cobertura vegetal presenta también un buen porcentaje de relación con la riqueza de especies, lo que sugiere que a mayor densidad arbórea en el habitat se espera encontrar más especies crípticas de hormigas. El microhábitat que genera la presencia y diversidad de árboles de sombra está por tanto favoreciendo el establecimiento de hormigas crípticas de hojarasca. El índice de diversidad Simpson, al ser el más bajo en los bosques para el gremio de crípticas analizado, está indicando mayor diversidad (alta equitatibilidad y riqueza) de estos grupos en relación á los demás tratamientos.
Este estudio analizó el impacto ecológico y ambiental causado por las nuevas técnicas de siembra de café usando los ensamblajes de hormigas y algunos de sus gremios. Se espera que estas relaciones sirvan corno herramientas en futuros planes de conservación que fomenten la presencia de sombra en las plantaciones de café, sin afectar la producción, ya que el nivel de sombra puede mantenerse hasta en un 60% sin reducir la productividad del cafetal (Rice y Drenning 2003). La presencia de sombra arbórea no sólo hace la diferencia de calidad de habitat más significativa en términos de la ecología y la economía de los sistemas de café (Beer 1987) sino que es también un factor clave en el mantenimiento del equilibrio de la tierra (Beer et al. 1998) y de la diversidad y abundancia de organismos del suelo (Nestel et ai. 1993). Además, estos sistemas de cultivo pueden servir como modelo para concientizar a las nuevas generaciones de agricultores a pensar en pos del futuro de la tierra y de todos sus seres vivos. Prácticas agrícolas ambientalmente amigables se pueden implementar sin sacrificar la producción como se ha demostrado en esta misma región de Risaralda (Orgánica Tatamá, Francisco Herrera, com. pers. Nov. 2002). Al evaluar la adopción de prácticas menos intensivas, como las encontradas en cafetales tradicionales se pueden obtener mejores beneficios tanto para la tierra, como para el agricultor y la fauna asociada.
Conclusiones y Recomendaciones
• Los cafetales de sombra poligenérica contienen mayor riqueza faunística de hormigas. Esta riqueza decrece con la intensidad de los tratamientos y la disminución de la sombra, pero en general los resultados muestran que todos los cafetales de sombra son importantes para preservar la biodiversidad de hormigas: representan una matriz de alta calidad para contribuir a preservar especies de bosques o de vegetación natural. Aunque los parches de bosque de esta región presentaron una menor acumulación de especies con respecto a los cafetales de sombra poligenérica, el papel que juegan éstos como reservorio es de vital importancia para las especies exclusivas de hormigas que no se adaptan a altas perturbaciones agrícolas y para las especies crípticas de hojarasca.
• El análisis de las variables bióticas y abióticas medidas en las unidades muéstrales sugieren que tanto la temperatura como la cobertura vegetal, brindan una importante información en cuanto a la riqueza y abundancia de hormigas en los tratamientos, pero no todas las variables tomadas explican la presencia de especies y de individuos al momento de realizar el muestreo. Los tratamientos donde se obtuvieron promedios más bajos de cobertura vegetal y temperaturas más altas, fueron los más perturbados y menos diversos. Los tratamientos más ricos y abundantes en mirmecofauna, poseen altos niveles de cobertura y temperaturas de suelo más bajas en relación con los menos ricos y abundantes.
• Este estudio encontró que algunos gremios dentro del taxón Formicidae responden ecológicamente de formas opuestas a la modernización o intensificación del cultivo de café. Hay un grado de sensibilidad de los gremios de especies crípticas de hojarasca y las de cafetales, pero las especies abundantes no mostraron preferencia marcada por ningún tipo de tratamiento. Es más, algunas especies de hormigas se pueden ver beneficiadas con la transformación a cafetal de sol, abriendo la posibilidad de surgimiento de plagas por su asociación con homópteros.
• Para planes de conservación, es importante realizar estudios que permitan visualizar la importancia de las formas de cultivo amigables con la naturaleza. Estos estudios son importantes no sólo en el café sino también en diferentes cultivos que pueden estar sustentando la fauna de organismos tales como las hormigas. De otra manera, la conservación de la biota quedaría restringida a las áreas de reserva, las cuales cada vez son más escasas y vulnerables. Se sugiere, por tanto, implementar políticas para la conservación de áreas naturales de bosque pero complementada al mismo tiempo con un adecuado manejo de la matriz agrícola que usualmente rodea estos relictos naturales.
Agradecimientos
Los autores agradecen a Gustavo Álvarez, Elizabeth Jiménez, Gloria Vargas, quienes ayudaron con la realización de labores de campo y laboratorio. A Jack Longino, John Lattke, Fernando Fernández, James Rugler, James Trager, Tánia Arias, y Phillip Ward por la identificación de hormigas a nivel de especie. Este estudio fue financiado por el Programa de Ciencias de Medio Ambiente y del Habitat de Colciencias, código 1106-13-1693, la Universidad del Valle (Cali), Land Institute (Kansas) y la Universidad de Michigan (Ann Arbor, EEUU).
Literatura citada
AGOSTI, D.; MAJER, J. D.; ALONSO, L. E.; SCMULTS, T. R. 2000. Ants: Standard methods for measuring and monitoring biodiversity: Preface. Smithsonian Institution. Washington, D. C. [ Links ]
ANDERSEN, A. 1997. Using ants bioindicators: Multiscale issues ¡n ant community ecology. Conservation Ecology 1 (1): 8. [ Links ]
ANDERSEN, A. N. 2000. Global Ecology of rain forest ants, functional groups ¡n relation to environmental stress and disturbance. p. 25-34. En: D. Agosti, J. D. Majer, I. E. Alonso, & T. R. Shults (eds.). Ants: Standard methods for measuring and monitoring biodiversity: Smithsonian Institution. Washington, D. C. [ Links ]
ARMBRECHT, I.; PERFECTO I. 2003. Litter-twig dwelling ant species richness and predation potential within a forest fragment and neighboring coffee plantations of contrasting habitat quality in México. Agriculture, Ecosystems and Environment 97: 107-115. [ Links ]
BEER, J. 1987. Advantages, disadvantages and desirable characteristics of shade trees for coffee, cocoa and tea. Agroforestry Systems 5: 3-13. [ Links ]
BEER, J.; MUSCHELER, R.; KASS, D.; SOMARRIBA, E. 1998. Shade management in coffee and cacao plantations. Agroforestry Systems 38: 139-164. [ Links ]
BOLETÍN DE LA SOCIEDAD AGROPECUARIA TATAMÁ. 2002. E.mail: cafeapia@pereira.multi.net.co [ Links ]
BOLTON, B. 1994. Identification guide to the ant genera of the word. Harvard University press. Cambridge, Massachusetts. 222 p. [ Links ]
COLWELL, R. K. 1997. User's guide to EstimateS 6. Statistical estimation of species richness and shared species from samples. http://viceroy.eeb.uconn.edu/estimates. [ Links ]
COLWELL, R. R.; CODDINGTON, J. A. 1994. Estimating terrestrial biodiversity through extrapolaron. Philosophical Transactions of the Royal Society of London (serie B). 345: 101. [ Links ]
DELABLE, J. H.; AGOSTI, D.; DO NASCIMENTO, 1. C. 2000. Litter ant communities of Brazilian Atlantic rain forest región. Chapter 1. p. 1-17. En: D. Agosti, J. D. Majer, L. Alonso fie T. Schultz (eds.). Sampling Ground-dwelling ants: case studies from the world's rain forests. Curtin University School of Environmental Biology Bulletin. No. 18. Perth, Australia. [ Links ]
GOTELLI, N. J.; COLWELL, R. K. 2001. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters. 4:379-391. [ Links ]
MOLLDOBLER, B.; WILSON, E. O. 1990. The ants. Harvard University Press. Cambridge, Massachussets. 732 p. [ Links ]
IBARRA-NÚÑEZ, G. 1990. Los artrópodos asociados a cafetos en un cafetal mixto del Soconusco, Chiapas, México. Folia entomológica Mexicana 79: 207-231. [ Links ]
LUDWING, J. A.; REYTIOLDS, J. 1988. Statistical ecology: a primer methods and computing. Wiiey, new York. [ Links ]
MAJER, J. D. 1993. Comparison of the arboreal ant mosaic in Ghana, Brazil, Papua riew Guinea and Australia -its structure and influence on arthropod diversity. p. 115-141. En: J. LaSalle & I. D. Gauld (eds.). Hymenoptera and Biodiversity, CAB international. Wallingford, United Kingdom. [ Links ]
MOGUEL, R; TOLEDO, V. M. 1999. Biodiversity conservation in traditional systems of México. Conservation Biology 13: 11-21. [ Links ] [ Links ]
OSBORN, F; GOITIA. W.; CABRERA, M.; JAFFÉ, K. 1998. Ants, plants and butterflies as diversity indicators: Companson between strata at six forest sites in Venezuela. Fauna and Environment 1: 59-64. [ Links ]
PAOLETTI, M. G.; PIMENTEL, D.; STINNER, B. R.; STINNER, D. 1992. Agroecosystem biodiversity: matching production and conservation biology. Agriculture, Ecosystems and Environment 40: 3-23. [ Links ]
PERFECTO, I.; VANDERMEER. J. 1994. Unders-tanding biodiversity loss in agroecosystems: Reduction of ant diversity resulting from transformation of the coffee ecosystem in Costa Rica. Entomological trends in agricultural science. 2: 7-13. [ Links ]
PERFECTO, I.; ARMBRECHT, I. 2003. Tecnological change and biodiversity in the coffee agroecosystem of Mothern Latin América (Chapter 6). p. 159-194. En: J. Vandermeer (ed.). Tropical agroecosystems CRC Press LLC. Boca Ratón, Florida. [ Links ]
PERFECTO, I.; RICE, R. A.; GREENBERG, R.; VAN DER VOORT, M. E. 1996. Shade coffee: a disappearing refuge for biodiversity. Bioscience 46: 598-608. [ Links ]
PERFECTO, I.; VANDERMEER, J.; HANSON, P; CARTIN, V. 1997. Arthropod biodiversity loss and the transformation of a tropical agroecosystem. Biodiversity and Conservation 6: 935-945. [ Links ]
RAMOS-SUÁREZ. M. P; MORALES, H.; RUIZ-MONTOYA, L.; SOTO-PINTO, L.; ROJAS-FERNÁNDEZ, P. 2002. ¿Se mantiene la diversidad de hormigas con el cambio de bosque mesófilo a cafetales?, p. 16-30. En: A. Monro & M. C. Peña (eds.). Actas del Simposio café y Biodiversidad. Revista Protección Vegetal, Año 12 (2) ISBN 056501816. Edición Especial. San Salvador. [ Links ]
RICE, A.; DRENNING, J. 2003. Manual de café bajo sombra. Smithsonian Migratory Bird center (bird friendly). National Zoological Park, Washington, D.C. www.si.edu/srnbc. 62 p. [ Links ]
SWIFT, M.; VANDERMEER, J.; RAMAKRISHNAN, P. S.; ANDERSON, C; ONG, K.; MAWKINS, B. A. 1996. Biodiversity and agroecosystem function. p 261-298. En: H.A. Mooney, J. H. Cushman, E. Medina, O. E. Sala & E. D. Schulze (eds.). Functional Roles of Biodiversity: a global perspective. John Wiley &r Sons Ltd. [ Links ]
Recibido: Nov. 23 / 2003 Aceptado: Mar. 27/2004

