










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488On-line version ISSN 2665-4385
Rev. Colomb. Entomol. vol.33 no.2 Bogotá June/Dec. 2007
Sección Agrícola
Resistencia de especies de Manihot a Mononychellus tanajoa (Acariformes), Aleurotrachelus socialis y Phenacoccus herreni (Hemiptera)
Resistance of Manihot species to Mononychellus tanajoa (Acariformes), Aleurotrachelus socialis, and Phenacoccus herreni (Hemiptera)
MARITZA BURBANO M. 1, ARTURO CARABALÍ M.2, JAMES MONTOYA L.3, ANTHONY C. BELLOTTI4
1 Estudiante de Biología Universidad del Valle. merry2309@yahoo.com
2 Asistente de Investigación, Entomología de yuca, CIAT. a_carabali@yahoo.com
3 Profesor Departamento de Biología Universidad del valle. jamesmon@univalle.edu.co
4 Líder Proyecto Entomología de yuca, CIAT, A.A. 6713, Cali, Colombia. a_bellotti@cgiar.org (Autor para correspondencia).
Resumen: Las especies silvestres del género Manihot ofrecen una fuente potencial de genes de resistencia para el control de la mayoría de insectos plaga de yuca (Manihot esculenta). Recientemente, esta característica ha sido aprovechada para el desarrollo de variedades resistentes a plagas y enfermedades a través de cruces inter-específicos con especies silvestres de Manihot. Con el objetivo de buscar nuevas fuentes de resistencia en especies de Manihot, se evaluaron y compararon dos especies silvestres brasileras, Manihot flabellifolia y M. peruviana, frente a genotipos comerciales de M. esculenta, para determinar su potencial de resistencia a tres de las principales plagas de yuca en las Américas, Africa y Asia: el ácaro Mononychellus tanajoa, el piojo harinoso (Phenacoccus herreni) y la mosca blanca (Aleurotrachelus socialis). La investigación se realizó en CIAT, Palmira, bajo condiciones controladas (25±5 ºC, 60±10 HR, 12L/D) utilizando una metodología de tamizados y bioensayos asistida por escalas de infestación y daño. Los resultados mostraron que los genotipos de M. flabellifolia (444-002) y M. peruviana (417-003 y 417-005) exhibieron altos niveles de resistencia frente a M. tanajoa y A socialis, mientras, el genotipo de M. flabellifolia (444-002) y M. peruviana (417-003) presentaron niveles intermedios de resistencia a P. herreni. Se discute la importancia de estas fuentes de resistencia a la luz de los programas de conservación de la diversidad y de la resistencia de planta hospedera como un componente del manejo integrado de plagas.
Palabras clave: Yuca. Mosca blanca. Resistencia planta hospedera.
Abstract: Wild species of the genus Manihot offer a valuable and potential source of resistance genes for the control of the majority of insect pests of cassava (Manihot esculenta). Recently, this characteristic has being exploited to develop strains resistant to pests and diseases through inter-specific crosses with wild Manihot species. In order to find new sources of resistance in Manihot species, two wild Brazilian species, M. flabellifolia and M. peruviana, were evaluated and compared to commercial genotypes of M. esculenta to determine potential resistance against three of the major cassava pests in the Americas, Africa and Asia: the mite Mononychellus tanajoa, the mealybug (Phenacoccus herreni) and the whitefly (Aleurotrachelus socialis). The research was carried out at CIAT, Palmira, under controlled conditions (25±5 ºC, 60±10 RH, 12L/D) using screening and bioassay methodologies assisted by controlling infestation and damage levels. The results showed that the genotypes M. flabellifolia (444-002) and M. peruviana (417-003 and 417-005) exhibited highest levels of resistance against M. tanajoa and A. socialis, while the genotypes M. flabellifolia (444-002) and M. peruviana (417-003) showed middle levels of resistance against P. herreni. The importance of these sources of resistance is discussed in light of the diversity conservation and host plant resistance programs and a component of integrated pest management.
Key words: Cassava. Whitefly. Host plant resistance.
Introducción
Manihot Miller, Euphorbiaceae, es un género nativo del Neotrópico que ocupa un amplio rango de hábitats, desde el sur de Arizona hasta Argentina (Rogers y Appan 1973). Sus especies son perennes y varían desde arbustos acaulescentes hasta árboles de 10 a 12 metros de altura. La mayoría presenta raíces tuberosas y algunas acumulan grandes cantidades de almidón, como la yuca (Manihot esculenta Crantz), uno de los cultivos tropicales más importantes del mundo, la cual es producida, principalmente, por pequeños agricultores como fuente alimenticia e industrial (Cock 1985). M. esculenta es tolerante a la sequía y a suelos degradados, además de tener gran flexibilidad respecto a la plantación y la cosecha que le confieren alta adaptabilidad a una gran variedad de ecosistemas tropicales donde, es un elemento básico en la dieta de alrededor de 500 millones de personas (FAO/FIDA 2000). Lamentablemente, las características agronómicas de este cultivo favorecen, la presencia de artrópodos plaga que causan grandes pérdidas como consecuencia de daños directos e indirectos (Bellotti et al. 1994). En la actualidad tres especies son los artrópodos que más afectan y limitan el aprovechamiento de este cultivo: el ácaro verde de la yuca (AVY), Mononychellus tanajoa (Bondar, 1938), especie que causa pérdidas en el rendimiento de raíces entre el 13-80% en las Américas y África (Skovgard et al. 1993; Bellotti et al. 1999); el piojo harinoso, Phenacoccus herreni (Cox-Willians, 1981), en altas poblaciones puede causar pérdidas en el rendimiento hasta del 80% en el norte de Sur América y Norte-Este del Brasil (Vargas y Bellotti 1981) y la mosca blanca, Aleurotrachellus socialis (Bondar, 1923), presente en Ecuador, Venezuela, Brasil y en Colombia, donde ocasiona pérdidas hasta del 79% (CIAT 1986). Para contrarrestar los daños ocasionados por estas plagas, se utilizan generalmente plaguicidas químicos (Smith 1989), pero su uso indiscriminado ha generado efectos negativos en los insectos benéficos, ha propiciado el desarrollo de resistencia a los insecticidas y, en general, ha producido altos niveles de contaminación ambiental (Smith 1989; Panda y Khush 1995). Adicionalmente, su empleo es una opción costosa para el pequeño productor (Arias y Guerrero 2000). Ante estos hechos, se hace necesaria la búsqueda de alternativas de manejo que permitan mantener las poblaciones plagas por debajo del nivel de daño económico. El manejo integrado de plagas (MIP), combina resistencia vegetal a plagas con métodos de control, biológico, cultural y químico. La resistencia de planta hospedera (RPH) es la base del Manejo Integrado de Plagas (MIP), por su compatibilidad con otros métodos de control, bajo costo y compatibilidad con el ambiente (Bellotti et al. 1993).
Las especies silvestres de Manihot son conocidas “fuentes” de genes de resistencia a prácticamente todas las plagas y enfermedades de yuca, como también, al deterioro fisiológico post-cosecha (PPD). No obstante su valor, han sido poco utilizadas en el mejoramiento de yuca, con el agravante que dichas especies están amenazadas por extinción, debido a cambios del medio y destrucción de sus hábitats naturales (CIAT 2004).
Con el objetivo de buscar nuevas fuentes de resistencia, en este estudio se evaluaron y compararon dos especies silvestres brasileras (M. flabellifolia y M. peruviana) frente a genotipos comerciales de Manihot esculenta, para determinar sus potenciales de resistencia a tres plagas claves de yuca en las Américas, África y Asia: el ácaro Mononychellus tanajoa, el piojo harinoso Phenacoccus herreni y la mosca blanca Aleurotrachellus socialis.
Materiales y Métodos
Plantas y Artrópodos. En el programa de entomología de yuca (CIAT) se propagaron 20 plantas de los genotipos CMC- 40, MECU-72 de M. esculenta Crantz; MFLA 444-002 de M. flabellifolia y MPER 417-0003, MPER 417-005 de M. peruviana (Mueller) y se transplantaron posteriormente a bolsas plásticas y materas. Cinco plantas de cada genotipo, de 40 días de edad, se introdujeron individualmente en jaulas de tul y madera de 0,5 m x 0,5 m x 1m.
Los especimenes de mosca blanca, A. socialis, y del piojo harinoso, P. herreni, se recolectaron de colonias independientes establecidas en CIAT. La colonia de ácaros M. tanajoa fue reestablecida siguiendo la metodología propuesta por Lenis et al. (1993).
Evaluación de resistencia. Para la identificación de los niveles de resistencia presentes en genotipos y especies silvestres de Manihot se utilizaron infestaciones artificiales de especimenes de cada una de las plagas. Las evaluaciones se realizaron bajo condiciones controladas de temperatura, humedad relativa y fotoperiodo (25±5 ºC, 60±10 HR, 12L/D).
La infestación con el acaro M. tanajoa se realizó sobre cinco plantas de cinco genotipos, M. esculenta (CMC-40; MEcu-72), M. flabellifolia (MFLA 444-002) y M. peruviana (MPER 417-003; 417-005), las cuales se introdujeron individualmente en jaulas de tul y madera de (0.5m x 0.5m x 1m), cada una con tres repeticiones y dispuestas al azar. La infestación se realizó sobre las hojas superiores de cada genotipo y especie silvestre a evaluar, colocando dos lóbulos de hojas infestadas con 200 ácaros por cinco días. Al finalizar este periodo las hojas viejas se retiraron. Las evaluaciones de infestación y daño se realizaron 10 días después de la infestación y se continuaron cada cinco días durante cuatro semanas.
La infestación de genotipos y especies silvestres con piojo harinoso (P. herreni) se realizó utilizando huevos dispuestos en ovisacos provenientes de la colonia de P. herreni. Para una mejor colonización los ovisacos se colocaron sobre la axila de la hoja con la ayuda de un punzón. Los huevos del piojo harinoso contienen una sustancia pegajosa que facilita su adherencia a las plantas. La primera evaluación se realizó a los 10 días y se continuaron las evaluaciones cada 10 días durante dos meses. Así mismo, la infestación se realizó en jaulas individuales con cinco plantas de cinco genotipos, cada una con tres repeticiones y dispuestas al azar.
La infestación con Aleurotrachellus socialis en cada genotipo y especie silvestre se realizó con 200 adultos de A. socialis recién emergidos. Hojas de M. esculenta, con aproximadamente 500 pupas (se distinguen por tener los ojos rojos), se tomaron de la colonia de mosca blanca y colocadas en una caja-jaula de madera de 30 cm x 30 cm x 30 cm, forrada con tul negro en tres de sus caras y en la superior con vidrio. Las hojas permanecieron en la caja durante 12 horas, después de las cuales se retiraron los adultos emergidos y con la ayuda de un aspirador manual de pipeta se colocaron en las hojas más jóvenes de las plantas a evaluar. Las evaluaciones se realizaron cinco días después de la primera infestación y cada 10 días durante dos meses.
Evaluación de infestación y daño. Las evaluaciones para las tres especies se realizaron por observación directa. Este tipo de evaluación puede ser adaptable a muchas asociaciones insecto-planta, a la vez que permite examinar un gran número de plantas en corto tiempo. Este procedimiento se basa en escalas que califican el grado de infestación y, por otra parte, el grado de daño que ha sufrido la planta. La escala de infestación, define cualitativamente la cantidad aproximada de insectos que se encuentran en la planta en un momento dado y en qué parte de ésta se concentran. La escala de daño permite definir o categorizar en alto, intermedio o susceptible el nivel de resistencia manifestado por el deterioro de las plantas. Esta escala está generalmente comprendida entre uno y seis.
Para ácaros y piojos harinosos también se realizó una división de la planta en tres estratos, alto (cogollo y hojas nuevas), medio (hojas medias) y bajo (hojas viejas) y se observó la distribución de los diferentes estados de la plaga en las diferentes partes de la planta, estos datos se consignaron teniendo en cuenta las escalas de daño y grados de infestación.
Para evaluar el grado de infestación y daño para las plantas infestadas con moscas blancas se siguió el método desarrollado por Arias (1995), mediante el cual se estima un porcentaje del número de formas adultas, huevos, ninfas y pupas presente en cada uno de las tres repeticiones de los genotipos evaluados en los tres estratos de la planta. De cada parte se examina una hoja al azar en cada muestreo por observación directa. De estas tres observaciones, posteriormente, se calcula un promedio que corresponde al dato obtenido para cada muestreo.
En general, para la evaluación del daño y población que ocasionan las especies M. tanajoa, P. herreni y A. socialis sobre los genotipos de M. esculenta y especies silvestres de Manihotfueron empleadas las escalas de población y daño definidas por Bellotti y Kawano (1983) para ácaros, Castillo (1985) para piojos harinosos y Arias (1995) para A. socialis, las cuales usan rangos de uno a seis, donde uno indica ausencia de población y no daño; mientras seis indica muy alta población plaga y daño severo (Tablas 1, 2 y 3).
Análisis estadístico. Los valores promedios de daño e infestación, se sometieron a un análisis de normalidad (Kolmorov-Smirnov) y de homogeneidad de varianzas (Levene). En aquellos casos en que los datos no fueron homogéneos se realizó transformación de raíz cuadrada y se llevó a cabo un análisis de varianza de una vía para determinar si existían diferencias significativas para cada variable. Cuando se encontraron diferencias significativas se utilizó la prueba de comparación múltiple de Tukey.
Resultados y Discusión
Mononychellus tanajoa. El análisis comparativo mostró que existen diferencias significativas en el nivel de infestación y daño de M. tanajoa entre los genotipos evaluados (F11.132 = 29,86, F11.132 = 66,72; P<0.05). En el análisis de la variable infestación, el genotipo de M. flabellifolia (444-002) difiere de los genotipos de M. esculenta, pero no difiere de uno de los dos genotipos de M. peruviana MPER 417-005 (Tukey P<0.05). No se presentaron diferencias significativas entre los dos genotipos de M. peruviana, pero estos si difirieron al ser comparados con los de M. esculenta. Dentro de los genotipos de M. esculenta estudiados no se presentaron diferencias significativas.
La infestación de M. tanajoa para los genotipos de M. esculenta (CMC-40 y MECU-72) fue alta (cinco y seis), mostrando mucha susceptibilidad a la infestación por el ácaro (Fig. 1). Los tres genotipos silvestres MFLA (444-002) de M. flabellifolia y MPER (417-0003, MPER 417-005 de M. peruviana presentaron valores de infestación entre uno y tres lo cual sugiere que son hospederos de menor preferencia por este ácaro, siendo M. flabellifolia (MFLA 444-002) el hospedero menos conveniente.
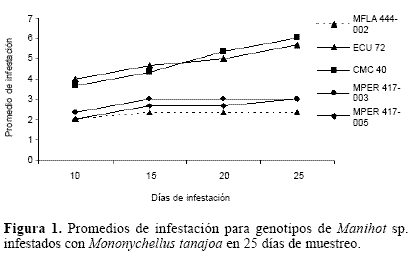
Por otro lado, se presentaron diferencias significativas para la variable daño debidas en gran parte al genotipo de M. flabellifolia (MFLA 444-002) el cual difiere de todos los genotipos de M. esculenta (CMC-40, MECU-72), pero fue similar a los de M. peruviana (MPER 417-003 y MPER 417- 005) (Tukey P<0.05). Estos últimos no presentaron diferencias significativas entre ellos, pero si, cuando fueron comparados con los de M. esculenta. Así mismo, entre los genotipos de M. esculenta no se presentaron diferencias significativas.
Los resultados mostraron que todos los genotipos de M. esculenta fueron altamente susceptibles ya que presentaron valores entre cinco y seis en la escala de daño, mientras que el genotipo MFLA 444-002 presentó el más bajo nivel de daño (2.33), seguido de los genotipos de M. peruviana MPER 417- 003 y MPER 417-005 con valores de tres, sugiriendo menor susceptibilidad de estos tres genotipos con respecto a los niveles de daño (Fig. 2).
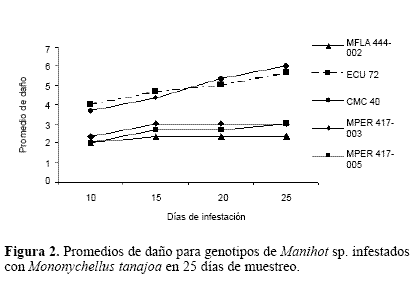
Los resultados en la variable infestación fueron consistentes con el daño de M. tanajoa. Sin embargo, aunque los datos para la variable daño en los genotipos silvestres los clasificaron como resistentes, los promedios de infestación dejan ver que más que resistencia lo que hay son niveles de tolerancia.
Phenacoccus herreni. Hubo diferencias significativas entre los genotipos evaluados para P. herreni en cuanto al nivel de infestación y daño (F11.204 = 8,24 y F11.204 = 2,83; P < 0,005). Para la variable infestación se encontró que MPER 417-003 tiene diferencias significativas con todos los genotipos evaluados a excepción de MFLA 444-002 (Tukey P < 0.05). Nuevamente existen diferencias entre los dos genotipos de M. peruviana estudiados.
Los resultados de la variable infestación para P. herreni conformaron dos grupos (Fig. 3).Un grupo conformado por los genotipos MPER 417-003, MFLA 444-002 con bajos niveles de infestación (dos y tres) lo cual los clasifica como resistentes. En el otro grupo, los genotipos presentaron altos grados de infestación (susceptibles). En contraste con el otro genotipo de M. peruviana, el genotipo MPER 417-005, presentó un alto nivel de infestación (> 3) que lo identifica como susceptible.
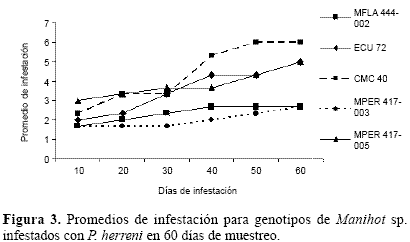
La variable daño para los distintos genotipos de Manihot infestados con P. herreni, permiten concluir que al igual que para ácaros, todos los genotipos de M. esculenta son hospederos convenientes para el piojo harinoso (altamente susceptibles), mientras que el genotipo MPER 417-003 presentó los más bajos niveles de daño, sugiriendo ser un hospedero no conveniente (resistentes) para P. herreni (Fig. 4). No obstante, es importante destacar que MPER 417-005, a pesar de ser un genotipo de la misma especie que el anteriormente mencionado (M. peruviana), exhibió niveles relativamente altos de daño, indicando que pueden existir variaciones intra e ínter específicas. Estos resultados sugieren, evaluar un mayor número de genotipos de cada especie.
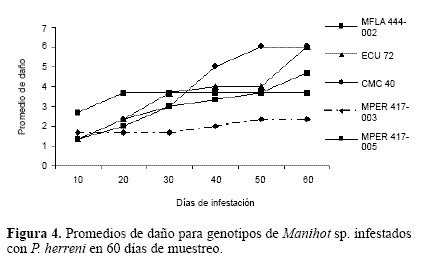
El genotipo MFLA 444-002 para esta plaga presentó un grado intermedio de daño (3,66). La diferencia expresada entre el grado de daño y el de infestación cataloga a este genotipo como altamente susceptible, pues muestra que con poca infestación el daño es considerable. En síntesis, los resultados para el piojo harinoso P. herreni, mostraron que los genotipos MFLA 444-002 y MPER 417-003 presentan moderados niveles de daño, pero altos niveles de infestación. Estos valores sugieren altos niveles de tolerancia y se recomiendan posteriores evaluaciones.
Aleurotrachellus socialis. El análisis comparativo mostró que existen diferencias significativas entre los genotipos con respecto a las variables, infestación y daño (F7.136 = 12.3, F7.136 = 19.54; P < 0,005). Para la variable infestación no se encontraron diferencias significativas entre los genotipos silvestres, pero si al ser comparados con el genotipo MECU- 72 y CMC-40 (Tukey P < 0,05).
Las diferencias en los valores de infestación formaron para esta variable tres grupos (Fig. 5). Uno conformado por los tres genotipos silvestres MPER 417-003, MPER 417-005 y MFLA 444-002, con un promedio de infestación en la escala de uno que corresponde a un nivel alto de resistencia. Un segundo grupo correspondiente a MECU-72 con un promedio de infestación (tres y cuatro) que lo califica en un nivel medio de resistencia y, finalmente, un tercero conformado por CMC- 40 con un alto nivel de susceptibilidad.
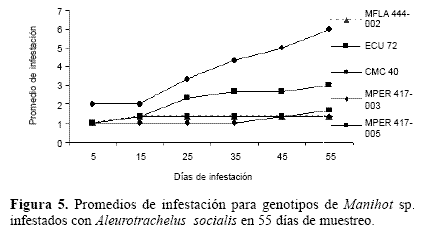
Los datos el grado de infestación para A. socialis, sugiere que para la mosca blanca los genotipos silvestres presentan niveles de resistencia. MECU-72 presentó niveles de tolerancia, pues poseen resistencia al nivel de daño pero un nivel medio para infestación y un porcentaje relativamente alto de colonización de A. socialis. Estos resultados concuerdan con lo reportado por Bellotti y Vargas (1986), los cuales proponen a MECU-72 como un genotipo tolerante a A. socialis. El genotipo CMC-40 presentó altos niveles de susceptibilidad. Este resultado concuerda con lo esperado para CMC-40, genotipo sobre el cual se encuentra la colonia de A. socialis establecida en CIAT.
El análisis de postanova para la variable daño mostró que entre los tres genotipos silvestres (MFLA 444-002, MPER 417- 003 y MPER 417-005), no se presentan diferencias significativas, pero sí entre ellos y los genotipos de M. esculenta. Así mismo, MFLA 444-002 difirió significativamente de CMC- 40. Los dos genotipos de M. peruviana fueron similares entre sí, pero si difieren de CMC- 40. El genotipo MECU-72 no difirió de ninguno de los genotipos silvestres, pero sí de CMC 40 (Tukey P<0.05).
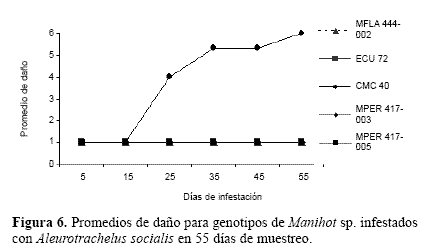
Los valores de daño para los distintos genotipos de M. esculenta y especies de Manihot infestados con A. socialisconformaron dos grupos para esta variable (Fig. 6). Uno formado por las especies silvestres (MPER 417-003, MPER 417-005 y MFLA 444-002) y ECU 72 con un daño en escala (< 2) que corresponde a niveles importantes de resistencia a la mosca blanca. Un segundo grupo formado por el genotipo CMC 40 con valores de daño (cinco y seis) valor que lo clasifica como un hospedero altamente susceptible a A. socialis.
Conclusiones
En términos generales, se presentaron diferencias entre los genotipos de Manihot esculenta y sus parientes silvestres evaluados en este estudio, con respecto a la resistencia a las tres plagas estudiadas. Estas diferencias podrían explicarse filogenéticamente por el concepto de acervos genéticos. Roa (1997) encontró que tanto por métodos de diferenciación morfológica como molecular, se evidencia una estrecha cercanía entre las poblaciones silvestres brasileras de M. esculenta ssp. flabellifolia, M. esculenta ssp. peruviana y los cultivares de M. esculenta. Así mismo, propuso que estas dos especies poseen una mayor proporción de diversidad genética con respecto a las poblaciones de M. esculenta. Esta diversidad genética podría proveerle a las especies silvestres una mayor capacidad de defensa contra las plagas de cultivo lo cual sería consecuente con lo encontrado en el presente estudio.
Desde el punto de vista genético, la resistencia que se presenta en los genotipos de yuca y sus parientes silvestres es probablemente heredada multigenéticamente y de tipo horizontal o de campo. Investigaciones recientes indican que no hay inmunidad, excepto para Spaceloma manihoticola. Existe resistencia a la mayoría de los insectos y enfermedades a niveles bajos e intermedios (Bellotti y Kawano 1983) en los cultivares existentes.
El germoplasma es evaluado para determinar resistencia a diferentes plagas y para identificar y seleccionar poblaciones resistentes. Posiblemente altos niveles de resistencia no pueden encontrarse en una sola variedad. Si diferentes genes de efecto aditivo están involucrados, cruces entre genotipos pueden incrementar el nivel de resistencia. Una vez que la resistencia ha sido identificada para diferentes plagas, los genotipos pueden ser cruzados, para un aumento de resistencia si es necesaria. Adicionalmente, los niveles de resistencia encontrados en las especies silvestres M. flabellifolia y M. peruviana, posibilitaran en un futuro cercano, con la ayuda de las herramientas moleculares, combinar genes de resistencia a M. tanajoa, P. herreni y A. socialis dentro de variedades comerciales.
Literatura citada
ARIAS, B. 1995. Estudio sobre el comportamiento de la "Mosca Blanca" Aleurotrachelus socialis Bondar (Homoptera: Aleyrodidae) en diferentes clones de yuca Manihot esculenta Crantz. Tesis de Maestría. Universidad Nacional de Colombia, Facultad de Agronomía. Palmira-Colombia. 181 p. [ Links ]
ARIAS, B.; GUERRERO, J. M. 2000. Control de plagas de la yuca (Manihot esculenta Crantz) por resistencia varietal. Sociedad Colombiana de Entomología SOCOLEN. Memorias XXVII Congreso Medellín - Colombia, Julio 26-28, pp. 243-259. [ Links ]
BELLOTTI, A. C.; KAWANO, K. 1983. Mejoramiento para resistencia varietal en el cultivo de la yuca. En: Domínguez, C. Yuca: Investigación, producción y utilización. CIAT, Cali, Colombia. 171- 193 p. [ Links ]
BELLOTTI, A. C.; VARGAS, O. 1986. Mosca Blanca del cultivo de yuca: Biología y Control. Cali, Colombia. Centro Internacional de Agricultura Tropical (CIAT). Serie: 04SC-04.05. 34 p. [ Links ]
BELLOTTI, A. C.; REYES, J. A.; VARELA, A. M. 1993. Observaciones de los piojos harinosos de la yuca en las Américas; su biología, ecología y enemigos naturales, pp. 313-337. En: Domínguez, C. Yuca: Investigación, producción y utilización. CIAT, Cali, Colombia. [ Links ]
BELLOTTI, A. C.;BRAUN, A. R.; RIAS, B.; CASTILLO, J. A.; GUERRERO, J. M. 1994. Origin and management of neotropical cassava arthropod pests. African Crop Science Journal 2(4): 407- 417. [ Links ]
BELLOTTI, A. C. SMITH, L.; LAPOINTE, S. L. 1999. Recent advances in cassava pest management. Annual Review of Entomology 44: 343-370. [ Links ]
CASTILLO, J. A. 1985. Informe de las investigaciones realizadas por el CIAT entre 1976 y 1985 sobre Phenacoccus gossypii y P. herreni dos especies de piojos harinosos plagas en los cultivos de yuca, proyectos futuros. Centro Internacional de Agricultura Tropical (CIAT) Cali, Colombia.18 p. [ Links ]
CIAT, 1986. Mosca blanca en el cultivo de la yuca: biología y control. Serie 045C-0405. Audiotutorial. Centro Internacional de Agricultura Tropical, Cali, Colombia. 34 p. [ Links ]
CIAT, 2004. Annual report: Integrated pest and disease management in major agoecosystems. CIAT, Cali, Colombia. 12-95 p. [ Links ]
COCK, J. H. 1985. Cassava; New potential for a neglected crop. Boulder: Westview Press, 191 p. [ Links ]
FAO y FIDA. (Fondo Internacional de Desarrollo Agrícola/ Organización de las Naciones Unidas para la Agricultura y la Alimentación) 2000. La economía mundial de la yuca: hechos, tendencias y perspectivas. Italia 59 p. [ Links ]
LENIS, J. I.; BRAUN, R.; MESA, N.; DUQUE, M. 1993. Uso de escalas para la estimación de poblaciones de ácaros Tetranychidae en cultivos de yuca. p 139-172. En: Braun, R. Bases fundamentales para investigación sobre los ácaros plagas y sus enemigos naturales en el Ecuador. CIAT, Cali-Colombia. [ Links ]
PANDA, N.; KHUSH, G. S. 1995. Host plant resistance to insects. International Rice Research Institute, Philippines. 431 p. [ Links ]
ROA, A. C. 1997. Estimación de la diversidad genética en Manihot sp. mediante marcadores moleculares. Tesis de magíster en Ciencias, Biología. Universidad del Valle, Cali, Colombia. 112 p. [ Links ]
ROGERS, D. J.; APPAN, S. G. 1973. Manihot, Manihotoides (Euphorbiaceae), Flora Neotropica, Monograph 13. New York, Hafner Press. [ Links ]
SKOVGARD, H.; TOMKIEVWICZ, J.; NACHMAN, G.; MUNSTER - SWENDSEN, M. 1993. The effect of the cassava green mite Mononychellus tanajoa on the growth and yield of cassava Manihot esculenta in a seasonally dry area in Kenya. Experimental Applied Acaraology 17: 41-58. [ Links ]
SMITH, C. M. 1989. Plant resistance to insects: A fundamental approach. John Wiley and Suns, New York. 286 p. [ Links ]
VARGAS, O.; BELLOTTI, A.C. 1981. Pérdidas en rendimiento causadas por moscas blancas en el cultivo de la yuca. Revista Colombiana de Entomología 7: 13-20. [ Links ]
Recibido: 16-ene-2006 - Aceptado: 7-ago-2007


{kind=link}
{kind=link}
{kind=link}