Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Los bosques neotropicales montanos son considerados puntos calientes de biodiversidad (Myers et al., 2000). Estos ecosistemas son característicos de la zona denominada El PILA, considerada una de las áreas protegidas más importante y extensa de Panamá abarcando 207 000 ha (Brenes et al., 2004). Presenta una alta diversidad de flora y fauna debido a la presencia de nueve de las doce zonas de vida reconocidas para Panamá, y según el mapa de vegetación de Panamá es el área protegida con mayor variación de vegetación natural del país ( Autoridad Nacional del Ambiente [Anam], 2004). Sin embargo, la construcción de proyectos hidroeléctricos y las malas prácticas en el ecoturismo se han convertido en las principales amenazas de pérdida de biodiversidad en áreas adyacentes al PILA (Anam, 2010).
La mayor diversidad y abundancia de briófitos se encuentra en estos bosques montanos (Salazar-Allen, 2001), donde cumplen funciones ecológicas relacionadas con mantener la productividad y acumulación de biomasa, forman parte del ciclo hidrológico (Pócs, 1980; Wolf, 1993a; Ah-Peng et al., 2017) y también son indicadores de condiciones ambientales o zonas de intervención (Holz y Gradstein, 2005; Slack, 2011; Benítez et al., 2015). En Panamá actualmente se registran alrededor de 1252 especies de briofitos, incluyendo 751 de musgos, 488 de hepáticas y 13 de antocerotes (Dauphin et al., 2006; Schäfer-Verwimp, 2014; Dauphin et al., 2015).
Dentro de la gran diversidad de briófitos en los bosques montanos, los briófitos epífilos (aquellos que crecen sobre las hojas) son un grupo importante. Estos junto con líquenes, hongos, algas verdes y cianobacterias, forman un entorno especial denominado filósfera (Coley y Kursar, 1996). Los briófitos epífilos son de importancia porque al igual que las especies que colonizan otros sustratos, contribuyen a retener la humedad en los bosques, mantienen la biodiversidad tropical y participan en procesos fisiológicos del ecosistema (Lücking y Lücking, 1996; Freiberg, 1998). Estas especies presentan variedad de adaptaciones morfológicas a la filósfera (Zhu y So, 2001) y la mayoría pertenecen a las hepáticas de la familia Lejeuneaceae (Gradstein, 1997; Zartman, 2003). Los briófitos epífilos se encuentran limitados por variables microclimáticas, estructura del bosque y características del sustrato donde se desarrollan (Lücking y Lücking, 1996), por ello se han utilizado como indicadores de fragmentación y cambios en el microclima (Malombe et al., 2016). Estas plantas son más vulnerables a perturbaciones que incluyan modificación de hábitat (Pócs, 1996; Zhu y So, 2001), por lo que cambios en condiciones microclimáticas como temperatura, humedad relativa y disponibilidad de luz influyen en la diversidad y establecimiento de estas especies (Coley y Kursar, 1996; Sonnleitner et al., 2009; Kraichak, 2014).
En los bosques montanos todas las investigaciones en diversidad y comunidades de briófitos epífilos se han focalizado en el sotobosque de varias regiones (Winkler, 1966; Lücking, 1997; Pócs, 1996). Además, todas las investigaciones se han restringido a los briófitos epífitos desde la base hasta el dosel (Wolf, 1993b; Holz et al., 2002; Acebey et al., 2003; Holz y Gradstein, 2005). A pesar que varios estudios se han enfocado en briófitos epífilos del dosel en bosques de tierras bajas (Coley y Kursar, 1996; Eggers, 2001; Gradstein y Ilkiu-Borges, 2009), son ausentes los estudios enfocados a los epífilos en el dosel de los bosques montanos. Estudios enfocados en analizar la relación de las comunidades de briófitos epífilos y diferentes hospedadores documentaron que no hay preferencias de las especies por el sustrato, y que cambios en el microclima (e. g. zonas verticales en un árbol hospedero) podrían modificar la estructura y composición de estas comunidades (Marino y Salazar-Allen, 1992; Benavides y Sastre-De Jesús, 2011). Por esta razón el presente trabajo evalúa por primera vez los cambios en la riqueza y composición de las comunidades de briófitos epífilos en las diferentes zonas del estrato arbóreo de Beilschmiedia costaricensis (Mez & Pittier) C.K. Allen (Lauraceae) en el bosque montano del Parque Internacional La Amistad, Tierras Altas, Chiriquí, Panamá.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se realizó al oeste de la República de Panamá, específicamente en un bosque montano a 2218 m de altitud (figura 1), dentro del sendero Panamá Verde en la región Pacífica del Pila-Pacífico (8°53’37.4’’ latitud N-82°38’52.5’’ longitud W). En este sitio se registra una temperatura media anual de 20°C y precipitación media anual de 5486 mm (datos obtenidos de la estación meteorológica del Pila). El dosel del bosque está caracterizado por arboles de más de 30 m de altura como Alnus acuminata y Wercklea woodsonii, mientras que Beilschmiedia costaricensis es característico del subdosel del bosque. El resto de las especies son arbustos del sotobosque pertenecientes a las familias Piperaceae, Melastomataceae, Rubiaceae, Ericaceae, Solanaceae y, además, helechos arborescentes. El hospedero seleccionado en el estudio fue B. costaricensis, ya que era el único hospedero dentro del subdosel que sus hojas estaban cubiertas de briófitos, es uno de los géneros pantropicales ubicado dentro de la familia con gran diversidad (Lauraceae) y presentan un crecimiento arbóreo o arbustivo (Nishida, 1999). En la zona neotropical las especies de este género se distribuyen desde México hasta Brasil y el centro de Chile, desde bosques húmedos premontanos hasta bosques montanos (Nishida, 1999; Monro et al., 2017).
Diseño y recolección de datos
Los árboles hospederos se seleccionaron al azar a ambos lados del sendero, ubicando en total seis árboles. El muestreo se efectuó entre los meses de septiembre de 2015 a julio del 2016. Para la colecta se consideraron tres zonas dentro del gradiente vertical siguiendo el protocolo de Johansson (1974), a 11 m (zona 3, parte interna de la copa del árbol), 13 m (zona 4, parte media de la copa del árbol), y 15 m (zona 5, parte externa de la copa del árbol). A partir de ahora en todo el documento vamos a considerar a la zona1 como A, la zona 2 como B y la zona 3 como C. Por cada árbol con ayuda de una podadora aérea y en cada una de las tres zonas, se recolectaron dos hojas al azar (seis hojas en total por cada árbol). En cada hoja se registró la presencia/ausencia de todos los briófitos epífilos presentes. Las muestras se recolectaron bajo el permiso de colecta SE/P-11-15 otorgado por la Dirección de Áreas Protegidas del Ministerio de Ambiente de Panamá y, luego, fueron depositadas en el laboratorio del herbario (UCH) de la Universidad Autónoma de Chiriquí en Panamá.
Para la identificación de los especímenes se observaron sus características morfológicas usando estereoscopio (Zeiss-Stemi DV-4), microscopio (Zeiss-PrimoStar) y posteriormente diversas claves taxonómicas (Reyes, 1982; Gradstein et al., 2001; Gradstein y da Costa, 2003; da Costa, 2008; Dauphin et al., 2008; Gradstein y Ilkiu-Borges, 2009; Allen, 2002, 2010). La clasificación taxonómica está basada en Goffinet et al. (2009) para los musgos y en Crandall et al. (2009); Pócs, et al. (2014) y Ye et al. (2015) para las hepáticas.
Análisis de datos
La riqueza específica por hoja se comparó mediante estimadores de riqueza no paramétricos (Chao 1, Chao 2, Jack 1 y Jack 2) para cada una de las tres zonas. Los estimadores se calcularon con EstimateS 8.2 (Colwell, 2013). Los cambios en la riqueza de especies entre las diferentes zonas se determinaron con un análisis de varianza de una vía (Anova). Previo al Anova se verificó la normalidad de los datos con la prueba de Shapiro-Wilk (p-valor= 0.3). Se realizó un análisis de escalamiento multidimensional no métrico (NMDS) con los valores de presencia de las especies, para visualizar la similitud de las comunidades epífilas en las tres zonas. Para estos análisis se utilizó la distancia euclídea como medida de similitud. Para determinar los cambios en las comunidades de las tres zonas se realizó un análisis de similitud Anosim (Oporto et al., 2015). El nivel de significancia se determinó con la prueba de Monte Carlo (1000 permutaciones). Todos los análisis se realizaron con el programa R (R Development Core Team, 2008) con el paquete estadístico “vegan” (Oksanen et al., 2016).
RESULTADOS
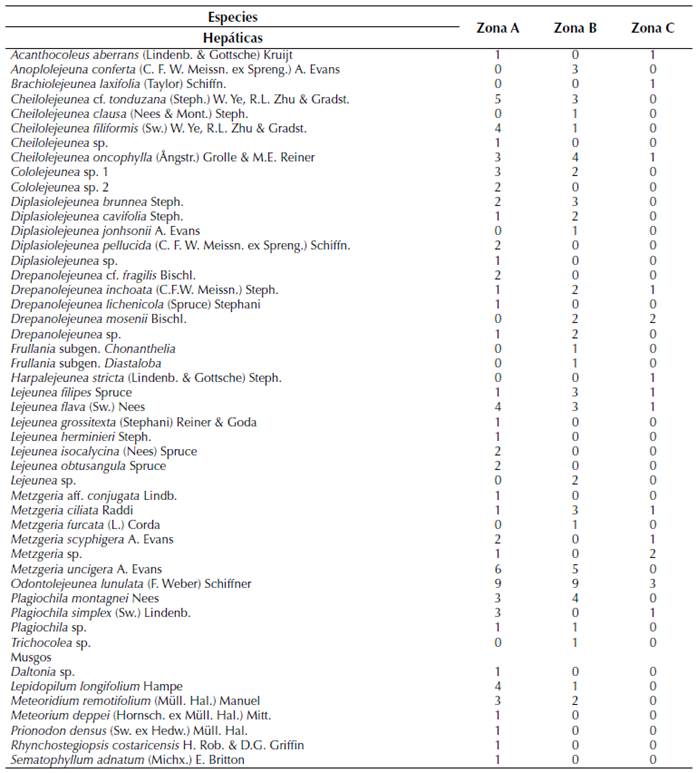
Se registró un total de 48 especies de briófitos epífilos (anexo 1), distribuidas en cinco familias, 19 géneros, 41 especies de hepáticas y, por otra parte, seis familias, siete géneros y siete especies de musgos. En las hepáticas foliosas la familia con mayor diversidad fue Lejeuneaceae, representando el 70 % de las especies, seguido de las hepáticas talosas de la familia Metzgeriaceae con el 19 %. La división Bryophyta representa un 11 % del total de especies, donde se encuentran familias como Meteoriaceae, Pilotrichaceae, Daltoniaceae, Sematophyllaceae, Leucomiaceae y Prionodontaceae.
En función de los estimadores de riqueza la zona A presentó un mayor número de especies y los valores más altos de especies estimadas en comparación con las otras zonas (tabla 1). El diagrama de cajas señaló un mayor número de especies por hoja para la zona A con un máximo de 15 especies, en comparación con las otras zonas (figura 2).
Tabla 1 Estimadores de la riqueza de especies de briófitos epífilos y desviación estándar por cada zona del gradiente vertical en el árbol hospedero. Zona A= 11 m; zona B = 13 m; zona C = 15 m
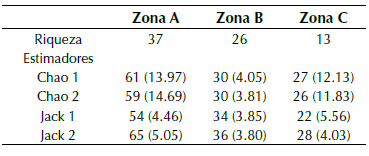
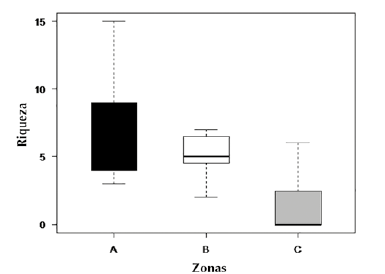
Figura 2 Diagrama de cajas con la riqueza de especies de briófitos epífilos por cada zona del gradiente vertical en el árbol hospedero.
El anterior resultado fue comprobado mediante la prueba Anova, la cual señaló que existen diferencias significativas (F=12.86; p <0.0001) en la riqueza de briófitos epífilos en cada uno de las tres zonas. El NMDS nos permitió visualizar que no hay diferencia en la composición de las comunidades de epífilas de las tres zonas (figura 3). Corroborando este patrón, el análisis de similitud (Anosim) señaló que no hay diferencias significativas en la composición de briófitos de las tres zonas (R= 0.0436; p> 0.05). Además, existe similitud en la composición de briófitos epífilos al comparar las tres zonas (tabla 2).
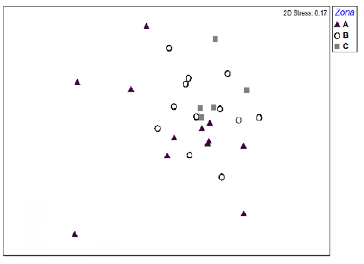
Figura 3 Análisis de escalamiento multidimensional no métrico para la composición de las comunidades por hoja en cada uno de las tres zonas de los árboles hospederos. Zona A (11 m) = triángulos negros; zona B (13 m) = círculos blancos y zona C (15 m) = cuadrados grises.

DISCUSIÓN
Los resultados indicaron que la riqueza de especies de briófitos epífilos cambia a lo largo de las tres zonas del hospedador B. costaricensis. Sin embargo, la composición de las comunidades fue similares en las tres zonas. Se ha documentado que la familia Lejeuneaceae forma el 90 % de las comunidades de hepáticas epífilas en la mayoría de los bosques tropicales (Cornelissen y Ter Steege, 1989; Marino y Salazar-Allen, 1992; Dauphin, 1995; Pócs, 1996; Zhu y So, 2001; Mota de Oliveira et al., 2009; Sierra et al., 2018). De manera similar, en el Sendero Panamá Verde la familia Lejeuneaceae representó el 70 % de la diversidad. Además, registramos que especies de musgos pertenecientes a las familias Brachytheciaceae, Calymperaceae, Daltoniaceae, Leucomiaceae, Meteoriaceae, Pilotrichaceae, Prionodontaceae y Sematophyllaceae están presentes en el sustrato epífilo, aunque varias de estas no se encuentran estrictamente en este hábitat (de Almeida, 2005; Müller y Pócs, 2007; Morales y Moreno, 2010).
La riqueza de especies en B. costaricensis disminuye a medida que se acerca a la parte superior de la copa de los árboles (zona C). En concordancia con lo que de Almeida (2005) y Gradstein y Ilkiu-Borges (2009) documentan sobre que en el sotobosque de los bosques húmedos tropicales de tierras bajas existe una mayor riqueza de especies epífilas en comparación con el dosel, patrón que está relacionado con una mayor humedad y menos intensidad de luz, las cuales son características distintivas de este estrato del bosque. De igual forma, en nuestra área de estudio el mayor número de especies se registró en la parte baja de la copa (zona A) aproximándose a las condiciones que propicia el sotobosque donde existe mayor humedad y menor cantidad de luz en comparación con las zonas B y C.
Las comunidades de briófitos epífilos en B. costaricensis no reflejaron cambios entre las diferentes zonas. En este contexto Sonnleitner et al. (2009) señala que no hay correlación entre las comunidades de briófitos epífilos y las hojas de las plantas hospedadoras, y que factores relacionados con el microclima influyen directamente en estas comunidades. Marino y Salazar-Allen (1992), en un bosque semihúmedo de tierras bajas de la Isla de Barro Colorado, documentaron que el microclima (luz y humedad) limita las comunidades epífilas que crecen sobre los arbustos hospedadores Hybanthus prunifolius (Violaceae) y Psychotria horizontalis (Rubiaceae). Sin embargo, en este caso la elevación y diferencias en condiciones climáticas como la temperatura y humedad ambiental podrían ser factores que inciten variaciones en la composición de las comunidades de briófitos epífilos. Por consiguiente, recomendamos que en futuros estudios de análisis de comunidades de briófitos epífilos se comparen patrones de composición considerando bosques de bajas y altas elevaciones.
Además, las diferentes condiciones microclimáticas de luz, humedad y temperatura entre los estratos se han identificado como factores limitantes en las comunidades de briófitos epífilos (Sonnleitner et al. 2009; Malombe et al., 2016). Por ejemplo, Kraichak (2014) señala que la diversidad beta de briófitos epífilos sigue un patrón en función del microclima. En nuestro caso, B. costaricensis es un árbol que crece cerca y entre el dosel dominado por A. acuminata y W. woodsonii, por lo que se podría esperar que las condiciones de microclima sean similares entre las tres zonas. Futuros análisis considerando mayor distancia entre las zonas en estudios y descripción del microclima por zona, podrían proporcionar mayor información al respecto. En nuestro estudio las comunidades están compuestas principalmente en la zona A por Odontolejeunea lunulata (16 %), Cheilolejeunea oncophylla (11 %) y Drepanolejeunea mosenii (11 %). La zona B por Odontolejeunea lunulata (13 %), Cheilolejeunea oncophylla (6 %), Plagiochila montagnei (6 %) y la zona C por Odontolejeunea lunulata (11 %) y Cheilolejeunea filiformis (5 %). El resto de las especies en cada zona está representado de menos del 5 %.
Por otra parte, otras variables no registradas en este estudio relacionado con los rasgos del hospedador son importantes limitantes en la comunidad de briófitos epífilos. Por ejemplo, varios estudios han documentado que las características de las hojas del hospedador relacionados con la forma, tamaño, textura influyen en la composición de las comunidades de briófitos epífilos (Lücking y Lücking, 1996; Sonnleitner et al., 2009), Así, mismo Müller y Riederer (2005) señalan que las hojas de las plantas vasculares difieren en sus propiedades físicas y químicas. Por ejemplo, las hojas de B. costaricensis son grandes (hasta 17 x 9 m), su superficie es lisa en comparación con los árboles a su alrededor como es el caso de W. woodsonii que es de hoja ancha, pero con gran cantidad de tricomas, mientras que A. acuminata sus hojas tienen la superficie lisa, pero su área foliar es más reducida, lo que convierte a B. costaricensis en un hospedero apto para la colonización, aparte de su ubicación con relación a las otras dos especies dominantes del dosel. También pudimos evidenciar que los árboles de B. costaricensis albergan comunidades de briófitos epífilos accidentales, por ejemplo, briófitos epífitos de la corteza u otros sustratos que lograron establecerse y adaptarse al sustrato de las hojas. En otro contexto, Sierra et al. (2018) indican que la composición de especies en las comunidades epífilas no depende de la secuencia cronológica en el hospedador y proponen que los cambios no siguen una dinámica de sucesión, sino que es un proceso al azar. Como en nuestro caso la distancia entre las zonas era de aproximadamente dos metros, la composición de las comunidades también puede atribuirse a la capacidad de dispersión de los briófitos (Zartman et al., 2015).
CONCLUSIONES
Podemos concluir que la riqueza de especies de briófitos epífilos de B. costaricensis en el bosque montano del Parque Internacional La Amistad, Tierras Altas, Chiriquí, Panamá, fue mayor en las zonas cercanas al sotobosque que corresponde a la zona A, dicha zona presenta mayor humedad y menor incidencia de luz en comparación a las zonas próximas al dosel del bosque (zonas B y C). Sin embargo, la composición de las comunidades de briófitos es similar entre las zonas consideradas, por ello análisis comparativos entre las condiciones microclimáticas y rasgos de la hoja por zona podrían brindar mayor información en los estudios de patrones de distribución de la riqueza y composición de briófitos epífilos en un gradiente vertical.