











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Soybean seed quality is a key factor for successful crop establishment and yield, as it influences germination, initial vigor, and uniformity in plant development (Frandoloso et al., 2015; Delouche, 2021). Furthermore, the sanitary quality of the seed is crucial, since pathogen transmission can lead to productivity losses and make phytosanitary management in the field challenging (Tonello et al., 2019; Weerasekara et al., 2021). Different studies have shown that factors such as the production environment, processing, storage, and harvest time significantly affect seed health and physiological performance (Ávila et al., 2003; Zuffo et al., 2017; Fialho et al., 2023). Thus, the use of seeds of high physical and sanitary quality not only contributes to achieving high yields but also reduces the need for chemical treatments and supports agricultural sustainability (Jaques et al., 2022; Zuffo et al., 2018).
In this sense, climatic conditions during soybean development and maturation directly influence the physiological, sanitary, and commercial quality of the seed. Temperature, rainfall, and relative humidity, among others, influence key parameters such as vigor, germination, and chemical composition, thereby affecting crop establishment potential and the longevity of stored seeds (Chabaniuk et al., 2024; Thuzar et al., 2010; Weerasekara et al., 2021). For example, heat stress and excessive rainfall during reproductive stages can significantly reduce seed quality, increase health risks and decrease crop yield (Ávila et al., 2003; Khan et al., 2011; Naeve and Huerd, 2008). Therefore, understanding and anticipating the impact of climate variations is key to ensuring the sustainable production of high-quality soybeans (Alsajri et al., 2020; Jarecki and Bobrecka-Jamro, 2021). In Colombia, soybean production reached 228 141 tons in 2024, cultivated across 90 695 ha in the Andean, Caribbean, and Orinoquía regions. The department of Meta is the leading producer, with 89 % of the national output, which is primarily used to produce balanced feed for animal consumption (FENALCE, 2024).
The Colombian Orinoquía region is characterized by high annual water availability and low soil fertility (Amézquita et al., 2013). Precipitation is monomodal, ranging from 2200 to 2700 mm between April and December, with irregular patterns in frequency and intensity (Valencia and Ligarreto, 2010). The agroclimatic conditions for soybean grain and seed production in the Colombian Orinoquía region are diverse and contrasting. During the first semester, high rainfall is common, affecting soybean ripening and harvest by producing very humid grain, which in turn accelerates seed deterioration (Valencia- Ramírez and Tibocha Ardila, 2023). These variations in water availability affect the crop depending on the genotype and the time of stress occurrence.
Likewise, Shelar et al. (2008) states that excessive rainfall and high temperatures during maturity and harvest are key factors influencing the incidence of seed-borne fungi. The fungal genera reported to have higher incidence on soybean during the rainy season- and considered critical due to their impact on reducing seed quality -are Phomopsis sp., Fusarium sp., Cercospora sp., Colletotrichum sp., Macrophomina sp., Rhizoctonia sp., Curvularia sp., and Aspergillus sp (Cortina et al., 2017; Soesanto et al., 2020).
On the other hand, the association between traits of interest in plant breeding can be assessed through phenotypic correlations, which reflect genetic and environmental interactions. However, according to Rodrigues et al. (2010), simple correlation coefficients can be misleading due to the indirect effect of other traits. Therefore, Wright (1960) developed Path Analysis, a method that decomposes the correlations between variables into direct and indirect effects, thereby determining the contribution of independent to dependent variables (Pérez et al., 2013; Faria Bisinotto et al., 2017). This method, implemented in the SPS AMOS Guide 5.0 software (Arbuckle, 2003), is illustrated using arrow diagrams that indicate the direction and magnitude of the relationships (path coefficients), and assesses model fit using indices such as GFI, CFI, and RMSEA (Mondo et al., 2012). Knowing the correlation of a particular trait with others is essential for the indirect selection of desirable genotypes (Carpentieri-Pípolo et al., 2005). For example, Valencia-Ramírez and Ligarreto-Moreno (2012) identified the number of pods per plant and the number of pods with three grains as valuable traits for maximizing the yield of soybean varieties. On the other hand, Singh et al. (1978) reported a high negative association between germination and seed weight, with no significant association between protein and oil content and germination. Lastly, Xu et al. (2022) identified in a meta-analysis direct negative effects of precipitation (-0.29) on grain yield, which possibly was associated with low seed quality.
In consequence, seed producers of the Orinoquía region face significant challenges due to the soil and climate conditions, which affect seed quality. Therefore, the aim of this research is to identify the most suitable environments for seed production, proposing a strategy for selecting genotypes with the greatest potential for high seed quality. It is hypothesized that the interaction between the edaphoclimatic conditions of the production environment and the seed genotype significantly affects the germination and sanitary quality of soybean seeds.
Materials and methods
Study area
Six soybean varieties were evaluated in the Orinoquía region in five locations: two during the second semester of 2020 (B) (La Libertad (LIBB), and Taluma (TALB)), and three during the first semester of 2021(A) (La Libertad (LIBA), Taluma (TALA), and Leonas). The climatic conditions of the five locations are described in Table 1.
Table 1 Climatic conditions of the locations under study
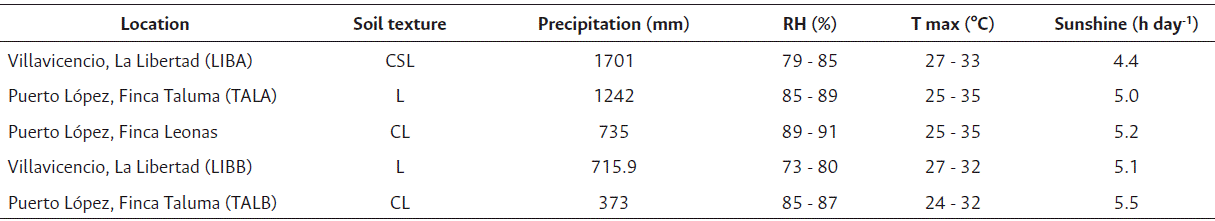
*RH: relative humidity; T max: maximum temperature; CSL: Clay Sandy Loam; L: Loam; CL: Clay Loam; LIBA: La Libertad, first semester 2021; LIBB: La Libertad, second semester 2020; TALA: Taluma, first semester 2021; TALB: Taluma, second semester 2020
To evaluate climatic variables, data were collected from the Davis Vantage Pro2 Plus meteorological stations located 3 km from the Taluma Experimental Farm and the La Libertad Research Center, ascribed to the Institute of Hydrology, Meteorology and Environmental Studies (IDEAM). Daily rainfall (mm), minimum and maximum temperatures (°C), relative humidity (%), and sunshine (h day-1) data from each location were processed for both semesters. Water excesses and deficits during the crop cycle were determined using the water balance model described by Valencia-Ramírez and Tibocha Ardila (2022).
Plant material
Six soybean varieties with different vegetative cycles and growth habits were evaluated in this study and are described in Table 2.

Soil characteristics
The study was carried out in soils of the flat highland and on high terraces of the Colombian Orinoquía region. The first area has class-IV Oxisols characterized by low organic matter (OM), high aluminum content, and low fertility. The second area has soils with medium to high aluminum saturation and medium to low exchangeable base content. Soil samples were collected at a depth of 0 - 20 cm to determine the physical and chemical characteristics at each location. The samples were subsequently analyzed in the Soil Chemistry laboratory of Tibaitatá Research Center (C.I. Tibaitatá) of AGROSAVIA. The main soil characteristics are described in Table 3.
Table 3 Physicochemical characteristics of soils at the study locations in the Colombian Orinoquía
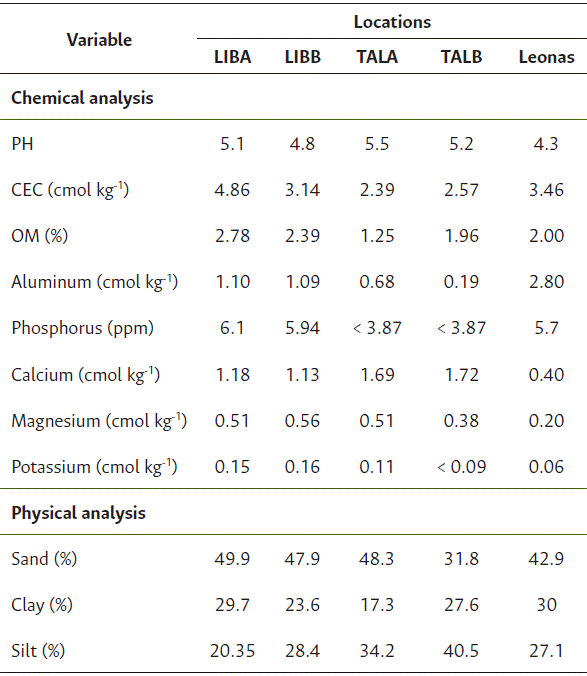
*CEC: Cation exchange capacity; OM: Organic matter; LIBA: La Libertad, first semester 2021; LIBB: La Libertad, second semester 2020; TALA: Taluma, first semester 2021; TALB: Taluma, second semester 2020; Leonas: first semester 2021.
Experimental conditions and evaluation variables
Planting was performed manually, and fertilization was carried out based on the results of the soil analysis and the nutritional requirements of soybeans (AGROSAVIA, 2006). The varieties were shown using a completely randomized block design, with four replicates per location. The experimental units consisted of six plots with 5-meter-long rows, spaced 50 cm apart. Vegetative and reproductive traits were evaluated using the two central rows as the useful plot area. Grain samples from each variety and location were collected for analysis of protein, oil, and amino acid content using an FT-NIR spectrometer (Bruker, USA).
Isolation of fungal pathogens
For the study of seed-borne fungi, Martín et al. (2022) reported that discrete probability distributions, such as Poisson regression and Negative Binomial distribution (NB), are useful for this type of study. However, Poisson regression may lead to overdispersion in count data analysis, which occurs when the observed variance exceeds the average. In such cases, the use of the NB regression model is appropriate, even for dichotomous variables with two possible outcomes, namely, the presence or absence of the fungus in the seed.
Samples from each variety were collected at harvest, according to their vegetative cycle, to conduct pathogen analyses in the Microbiology Laboratory of the La Libertad Research Center of AGROSAVIA. The most common fungi found in soybean seeds, including Phomopsis spp., Colletotrichum truncatum, Fusarium spp., and Cercospora kikuchii, were identified following the methodology described by ISTA (2024), with modifications.
Statistical analysis
The statistical analysis of the localities was performed using the General Linear Model (GLM) procedure in SAS Enterprise Guide 8.2. To assess the associations among germination-related variables, Pearson’s correlation coefficients were calculated, considering statistical significance at p < 0.05. Based on these results, the direct and indirect relationship between the variables was analyzed through a path analysis using the AMOS software (Pérez et al., 2013). The direct effects of one variable on another were estimated using standardized path coefficients, while the magnitude of indirect effects was assessed by multiplying the path coefficients along the pathways connecting the related variables (Pérez et al., 2013). Three predictive models were evaluated to identify the best fit: model 1 (semester A), model 2 (semester B), and model 3 (combined), using the goodness- of-fit statistics and reference criteria suggested by Pérez et al. (2013). The GENMOD procedure in the SAS software was used to evaluate fungal incidence in the seeds, assuming a negative binomial (NB) distribution.
Results and discussion
Influence of soil and climate conditions on germination
The soil and climate conditions across the evaluated environments with varieties were highly contrasting, significantly influencing seed physiological quality. Significant variations were observed in precipitation, water excess and deficit, relative humidity, temperature, and soil fertility (Tables 1, 2 and 3). During semester A, excess rainfall predominated (642 mm accumulated), whereas deficits were pronounced in semester B, as registered at the TALB location. At TALA, total rainfall reached 1242 mm, with relative humidity ranging from 85 % to 89 % and an average maximum temperature of 31 °C. This site exhibited the highest water excess between physiological maturity and harvest (305 mm), along with low fertility soils (OM: 1.25 %, CEC: 2.39 meq/100 g), which was reflected in a low average germination rate of only 35 % (Figure 1).
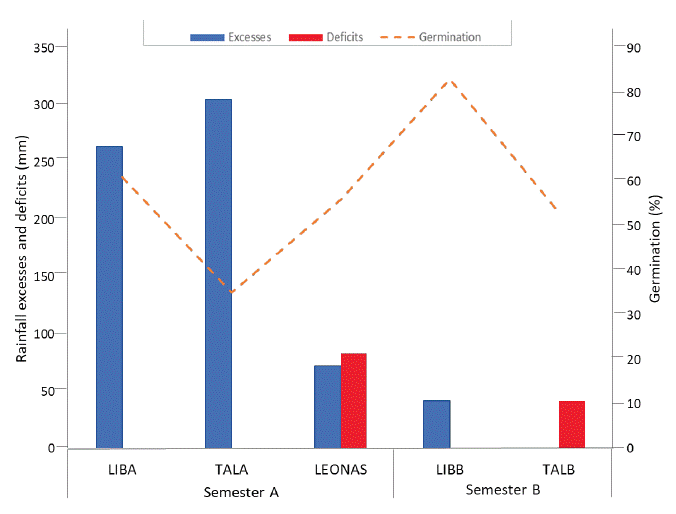
Figure 1 Rainfall excesses and deficits at maturity and harvest vs. germination stages during the first semester of 2021 (A) and second semester of 2020 (B) in the Orinoquía region, Colombia.
According to Delouche (1968), frequent rainfall lasting several days during the period of physiological seed maturity, combined with high temperatures, leads to a rapid decline in seed quality. According to Weerasekara et al. (2021), the occurrence of accumulated rainfall exceeding 100 mm during R6 - R8 is associated with a rapid decline in seed viability and vigor in the field, highlighting the vulnerability of this stage to excessive moisture.
LIBA location recorded high rainfall (1701 mm), relative humidity (RH) ranging from 79 % to 85 %, maximum temperatures of 29 °C, and moderate soil fertility (OM: 2.78 %, CEC: 4.86 meq/100 g). Despite water excesses exceeding 200 mm at the maturity stage, a germination rate of 60 % was observed, likely attributable to varietal traits such as earliness and efficient drying capacity. Leonas, with 735 mm of rainfall, high RH (89 % - 91 %), temperatures between 25 ºC and 35 °C, and low fertility (OM: 2.0 %, CEC: 3.46 meq/100 g), recorded a water deficit of 83 mm at the end of the cycle and an average germination rate of 56 % (Figure 1). Although no marked excesses were observed, the deficit combined with high atmospheric humidity could have limited physiological drying.
TALB presented the lowest rainfall during the study (373 mm), along with low soil fertility levels (OM: 1.96 %, CEC: 2.57 meq/100 g), and prolonged moisture deficits (> 4 days) during the period from maturity to harvest. These conditions led to reduced germination rates (53 %) (Figure 1). According to Valencia-Ramírez and Tibocha Ardila (2022), high temperatures combined with water deficit during grain filling may cause physiological changes such as stomatal closure, leading to premature leaf and flower drop, embryo abortions, pod loss, and reduced grain yield and quality.In contrast, the LIBB locality exhibited the most balanced conditions during semester B, with a total rainfall of 715 mm-which, according to Almanza (2006), is close to the crop’s water requirement-, moderate relative humidity, good soil fertility (OM: 2.38 %, CEC: 3.14 meq/100 g), and a low water excess at the maturity stage (41 mm). These favorable conditions contributed to germination rates exceeding 80 % (Figure 1).
This evidence confirms that the physiological quality of seeds depends largely on adequate water balance during critical developmental stages. Mondo et al. (2012) emphasize that well-nourished plants are better able to produce seeds of both higher quantity and quality, a trend that is particularly evident in environments with more fertile soils. This also reinforces the importance of ensuring optimal water and nutrient availability during the critical stages R3 - R6 to maximize seed quality (Tibocha Ardila, 2023).
Overall, the results indicate that both water stress due to excess (as recorded at TALA) and to deficit (as observed at TALB and Leonas) affect seed quality, with soil fertility playing a key modulatory role. Moreover, balanced environments such as LIBB should be prioritized for seed production, while more extreme environments should be monitored using more rigorous agronomic criteria.
Fungi in seeds
The seed health evaluation revealed that Phomopsis sp. and Fusarium sp. were the most frequently detected fungi infecting soybean seeds, with marked differences across locations. Foliar diseases like frogeye leaf spot (Cercospora sojina) and bacterial pustule (Xanthomonas axonopodis) were also identified in the field, although their presence was low in the study environments. Overall, the varieties exhibited low incidence levels of these foliar diseases, with Corpoica Superior 6 standing out as the least affected variety.
According to López-Cardona et al. (2022), among the pathogens with the greatest impact on seed quality are species from the Diaporthe/Phomopsis complex and the purple seed stain (or purple blotch) caused by Cercospora kikuchii (Formento, 2021). This is consistent with the higher incidence of Phomopsis sp. observed in this study.
Statistical analysis revealed significant differences in the incidence of Phomopsis sp. between locations and their interactions, as well as significant differences between locations, varieties, and interactions for Fusarium sp. Since the variance exceeded the mean, a negative binomial distribution was applied, which is appropriate for data with high dispersion. As shown in Figure 2, the highest probabilities of infection by Phomopsis sp. and Fusarium sp. were recorded during the first semester, particularly at the TALA location, which was characterized by excessive rainfall (642 mm) and temperatures over 30 °C. In contrast, during the second semester (at LIBB and TALB), the incidence was significantly lower due to the absence of excessive rainfall.
Gally et al. (2006) point out that when maturity and harvest coincide with periods of rainfall, grains can be colonized by Phomopsis sp., leading to reductions in seed size, weight, and germination capacity. Similarly, González-Vera et al. (2017) report that this fungus negatively affects seed health when high temperatures and rainfall occur at the maturity stage. Likewise, Henning (2005) associates its incidence with mechanical damage, excess moisture, and insect attacks. Regarding Fusarium sp., Cochran (2015) reports that it causes seed discoloration and loss of viability, particularly when harvest is delayed or seeds deteriorate under field conditions. Regarding varietal behavior, Corpoica Orinoquía 3, Corpoica Taluma 5, and BRS Barreiras exhibited a lower probability of infection by Phomopsis sp., while Corpoica Superior 6, Soyica P-34, and Agrosavia Primavera 11 were more susceptible. In contrast, for Fusarium sp., Corpoica Orinoquía 3 recorded the highest incidence, while Corpoica Superior 6 was the least affected (Figure 2).
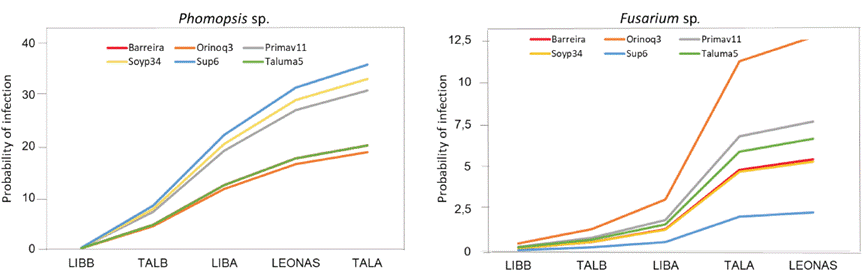
Figure 2 Probability of infection (incidence %) of Phomopsis sp. and Fusarium sp. in soybean seeds in five environments of the Orinoquía region, Colombia.
Concerning foliar diseases, the incidence of Cercospora sojina and Xanthomonas axonopodis was low across the study locations. Although all varieties exhibited low infection levels, Superior 6 was the least affected variety.
Path analysis for seed germination
The correlation coefficients from the combined analysis of germination and other variables of interest are presented in Table 4. The variables with a high contribution and positive and significant correlations were sunshine, oil, and lysine, while excess rain, Phomopsis sp., Fusarium sp., and days to harvest were negatively correlated with germination. These results are consistent with literature, as excessive rainfall reduces germination and creates ideal conditions for pathogen growth, which diminishes seed quality. However, it is essential to specify how the variables are interrelated and to distinguish their direct and indirect effects on germination.
Table 4 Phenotypic correlation coefficients of variables associated with seed germination under environmental conditions in the Colombian Orinoquía
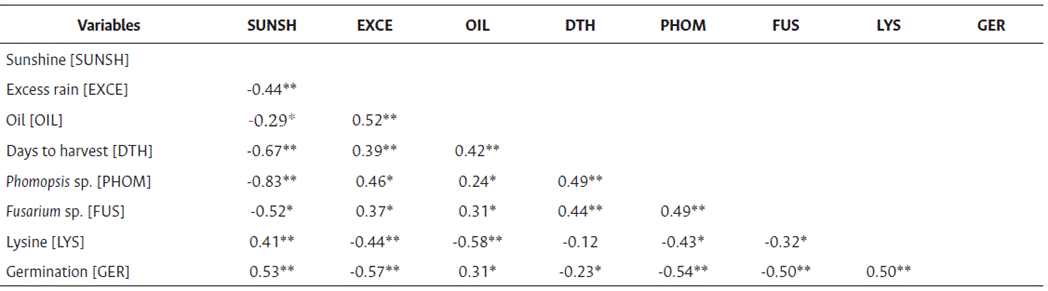
*Correlation is significant at p < 0.05 and **highly significant at p < 0.01.
In the predictive model analysis, model 3 showed the best fit with a goodness-of-fit index (GFI) of 0.95, a comparative fit index (CFI) of 0.97, a root means square error of approximation (RMSEA) of 0.09, and a lower Akaike information criterion (AIC) value (Table 5).
Table 5 Fit indices for models of variables associated with germination

*GFI: goodness-of-fit index; CFI: comparative fit index; RMSEA: root means square error of approximation; AIC: Akaike information criterion.
Path analysis explained 51 % of the observed variability in the endogenous variable (germination). Among the exogenous variables, sunshine had the most significant indirect effect on germination, mediated by its negative association with the incidence of Phomopsis sp. (path = -0.93) and Fusarium sp. (path = -0.45) (Figure 3). This suggests that greater solar radiation availability during crop development reduces the presence of these fungi in the seeds, improving their physiological quality.
This finding is supported by literature. Pathogens like Phomopsis sojae and Colletotrichum dematium, associated with seed infections, negatively affect germination and vigor, which results in increased electrical conductivity and decreased physiological potential (Wain-Tassi et al., 2012). In this sense, environmental factors, particularly light availability during the reproductive and maturation stages, play a determining role in pathogenic dynamics. Solar radiation can limit the sporulation and viability of phytopathogenic fungi, thereby reducing their capacity to colonize seed tissue. Consequently, variations in sunshine during seed filling and maturation stages represent a key modulating factor in the final quality of the lot by indirectly influencing inoculum pressure and the expression of seed- associated diseases.
Phomopsis sp. and Fusarium sp. exhibited direct negative effects on germination, with path coefficients of -0.17 and -0.24, respectively, confirming their detrimental impact on seed viability. These findings are consistent with those of TeKrony et al. (1984), who identified Phomopsis sp. as the main factor affecting seed quality, reporting a negative correlation (r = -0.88). Similarly, Gally et al. (2006) demonstrated that both Fusarium sp. and Phomopsis sp. Can simultaneously infect seeds, reduce the emergence of healthy seedlings and increase the incidence of abnormal or dead seeds.
Excessive rainfall had the strongest direct negative effect on seed germination, with a correlation coefficient of -0.40. However, it also exerted significant indirect effects through variables such as oil and lysine content. Specifically, excessive rainfall increased seed oil content (0.42), and this increase, along with the presence of Phomopsis sp., led to a significant reduction in lysine levels (-0.61 and -0.14, respectively) (Figure 3).
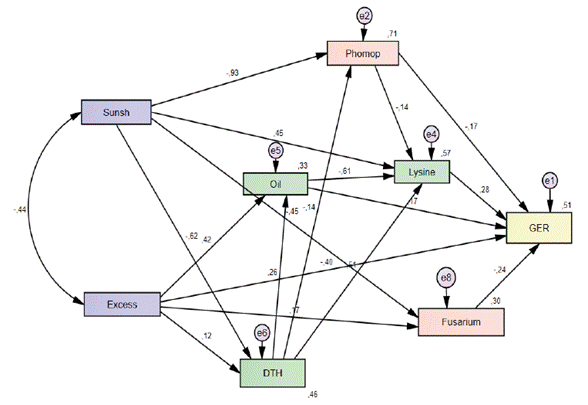
Lysine, as a key component of seed chemical composition, showed a direct positive relationship with germination (0.28), indicating that a higher lysine content is associated with improved germination capacity. This essential amino acid is limited in soybeans and is positively correlated with initial vigor, suggesting that its enhancement could improve both the nutritional value and the physiological performance of seeds (Li et al., 2024; Capelin et al., 2022). Likewise, recent advances such as genomic selection have enabled the identification of loci associated lysine content and other key amino acids involved in seed protein metabolism (Chen et al., 2025).
The variable days to harvest had an important indirect effect on germination by increasing the oil content (0.26). Previous studies have reported that the decline in seed physiological potential is associated with alterations in cell membrane composition, particularly in the fatty acids of the phospholipid bilayer, which can lead to ion leakage and loss of cellular structural integrity (Khare and Singh, 2023).
Bellaloui et al. (2017) reported a positive association between germination and palmitic acid levels, as well as a negative relationship between germination and linolenic and linoleic acids. Woyann et al. (2019) noted that soybean breeding has led to reduced levels of linoleic and linolenic acids, and increased abundance of oleic, stearic, and palmitic acids to improve the quality and stability of soybean oil used for biodiesel production.
Sunsh: Sunshine, DTH: Days to harvest, Phomop: Phomopsis sp., GER: germination.
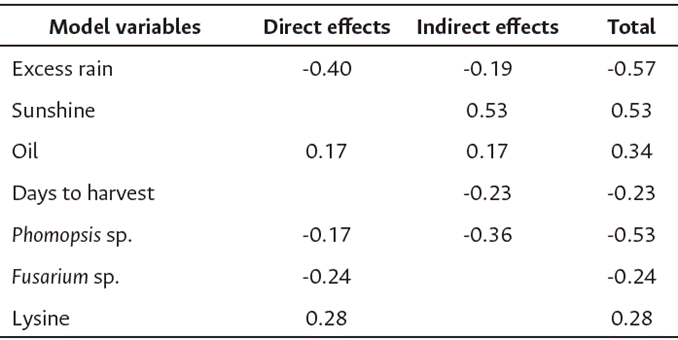
The path analysis coefficients of the direct and indirect effects of the climate and fungal variables on germination are described in Table 6. Days to harvest and lysine content are instrumental variables that can serve as selection criteria for optimizing soybean seed quality. Climate, pathogens, and oil content are valuable predictors of soybean germination under the environmental conditions of Colombian Orinoquía. These genetic traits are essential for closing the seed quality and quantity gap in the Orinoquía region, particularly in the highlands, where genetic improving remains the only viable strategy.
Conclusions
In the Colombian Orinoquía, higher seed quality was achieved during the second semester at the La Libertad Research Center, which is characterized by higher soil fertility, more favorable rainfall distribution, a dry period during physiological maturity, and a lower incidence of pathogens.
Phomopsis sp. and Fusarium sp. showed the highest probability of infection in Taluma during the first semester of 2021, a period marked by excessive rainfall (642 mm) and high temperatures (above 30 °C).
In soybean genetic improving programs focused on seed quality, days to harvest and lysine content are instrumental variables that should be considered as selection criteria. Additionally, excess water has a direct negative effect on germination, as it favors the proliferation of pathogens such as Phomopsis sp. and Fusarium sp. In contrast, sunshine exerts the most relevant indirect effect on germination by limiting the growth of these pathogens.

