










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.16 no.3 Bogotá Sept./Dec. 2011
GENÉTICA DEL COMPORTAMIENTO: ABEJAS COMO MODELO
Behavior Genetics: Bees as Model
GUIOMAR NATES-PARRA1, M.Sc. 1 Profesora Titular, Laboratorio de Investigaciones en Abejas, Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia. Sede Bogotá. mgnatesp@unal.edu.co
Presentado 8 de marzo de 2011, aceptado 9 de junio de 2011, correcciones 1 de julio de 2011.
RESUMEN
La abeja de miel (género Apis, Familia Apidae) es uno de los organismos utilizados en estudios de comportamiento, debido a su forma de vida social, la cual requiere de coordinación entre todos los individuos de la comunidad. La división de trabajo dentro de una colonia de abejas es consecuencia de cambios fisiológicos relacionados con la edad de las obreras y con la variación genética entre ellas que hace que realicen diferentes tareas. Con los progresos en biología molecular, genómica y secuenciación del genoma de Apis mellifera, han surgido nuevas herramientas que permiten desentrañar las bases moleculares del comportamiento, en particular el comportamiento social. Numerosos estudios han mostrado que muchas de las conductas realizadas por las obreras están determinadas genéticamente (comportamiento defensivo, comportamiento higiénico) y además que hay variación genética entre poblaciones en el desempeño de tareas como recolección de agua, néctar y polen. Igualmente algunos aspectos del comportamiento social, como el control de la reproducción en las castas estériles, también están bajo influjo genético. En este trabajo se hace una revisión de las metodologías utilizadas para estudiar la genética del comportamiento, así como la base genética de algunas de las conductas más sobresalientes de abejas.
Palabras clave: Apis mellifera, anarquía, comportamiento higiénico, comportamiento agresivo, división de trabajo.
ABSTRACT
The honeybee Apis mellifera (Apidae) is a model widely used in behavior because of its elaborate social life requiring coordinate actions among the members of the society. Within a colony, division of labor, the performance of tasks by different individuals, follows genetically determined physiological changes that go along with aging. Modern advances in tools of molecular biology and genomics, as well as the sequentiation of A. mellifera genome, have enabled a better understanding of honeybee behaviour, in particular social behaviour. Numerous studies show that aspects of worker behaviour are genetically determined, including defensive, hygienic, reproductive and foraging behaviour. For example, genetic diversity is associated with specialization to collect water, nectar and pollen. Also, control of worker reproduction is associated with genetic differences. In this paper, I review the methods and the main results from the study of the genetic and genomic basis of some behaviours in bees.
Key words: Apis mellifera, anarchy, hygienic behavior, aggressive behaviour, division of labor.
INTRODUCCIÓN
Que el comportamiento de los animales está determinado genéticamente, no es nuevo si se considera que, Darwin en su famosa obra -El origen del hombre-, en 1871 mencionaba que -los rasgos del temperamento de los animales son heredados-. Pero solo hasta hace casi 50 años fue que la genética del comportamiento surgió como una especialidad importante dentro de la genética; desde esa época muchos son los avances que refuerzan la idea generalmente aceptada de que todos los patrones comportamentales están determinados por componentes tanto ambientales como genotípicos.
El análisis del control genético de un determinado comportamiento es complicado por el hecho de que las acciones primarias de un gene pueden afectar: 1. Órganos sensoriales, cambiando la información recibida. 2. Sistemas intermedios (nervioso, endocrino), alterando capacidades de coordinación y percepción. 3. Órganos efectores, alterando la respuesta. Las mutaciones inducidas, que bloquean o alteran los patrones normales de comportamiento, proporcionan una herramienta muy útil para entender como los genes influencian la conducta. Al respecto hay muchos ejemplos, cuyo conocimiento ha servido para controlar o seleccionar caracteres indeseables o deseables (respectivamente), importantes para el mejoramiento en algunas especies animales. Por ejemplo, con la llegada de la abeja africanizada a América del Sur en 1958, a Brasil (Kerr, 1967) llegaron también varios inconvenientes generados por el fuerte comportamiento defensivo de la nueva subespecie introducida (Apis mellifera scutellata), lo cual generó la iniciación de programas de investigación tendientes a conocer la biología y comportamiento de la nueva especie introducida, de manera que se pudieran establecer cepas de abejas menos defensivas, conjuntamente con otras características como productividad o comportamiento higiénico.
El establecimiento de las bases moleculares del funcionamiento celular y del desarrollo ha producido importantes avances en las últimas décadas. Sin embargo, desde hace pocos años se despertó el interés por descubrir las bases moleculares del comportamiento, y especialmente, del comportamiento social. Actualmente mediante técnicas basadas en transcriptómica es posible medir cambios en la expresión génica en el cere-bro, relacionados con cambios en el comportamiento.
¿QUÉ ES LA GENÉTICA DEL COMPORTAMIENTO?
La complejidad de las características comportamentales hizo que durante mucho tiempo esas características fueran estudiadas e investigadas menos ampliamente y con menor éxito que aquellas características estructurales y numéricamente clasificables. Los caracteres del comportamiento están regidos tanto por condiciones ambientales como por el genotipo de un organismo determinado. El ADN del genoma determina los potenciales fisiológicos, estructurales y comportamentales del individuo, pero no todos estos potenciales se expresan de la misma forma en el individuo en desarrollo. Así la genética del comportamiento trata de los efectos del genotipo en el comportamiento y del papel que desempeñan las diferencias genéticas en la determinación de comportamientos distintos en una población.
Ante la pregunta ¿la herencia está afectando el comportamiento directamente, o sola-mente define la etapa en que algunos patrones del comportamiento pueden ser moldeados por factores ambientales?; sicólogos y genetistas han manifestado opiniones opuestas. Sin embargo, las nuevas tendencias involucran también a ecólogos, porque las adaptaciones son sencillamente el producto de la acción de la selección natural sobre los genes.
Cuando Charles Darwin postuló su teoría de la selección natural, no tenía ningún conocimiento sobre los mecanismos de la herencia. La teoría de Darwin se puede reformular en términos genéticos de la siguiente manera (Krebs y Davies, 1996):
- Todos los organismos poseen genes que codifican para la síntesis de proteínas; tales proteínas regulan el desarrollo de sistema nervioso, músculo y de la estructura del individuo, por tanto determinan su comportamiento.
- En una población muchos genes están presentes en dos o más formas alternativas, o alelos, que codifican para formas diferentes de la misma proteína. Esto podría provo-car diferencias en el desarrollo y por tanto habría variación dentro de la población.
- Habrá competencia entre los alelos de un gene por un sitio (locus) particular en los cromosomas, puesto que solo uno de los posibles alelos puede ocupar un locus.
- Cualquier alelo que pueda hacer un número de copias sobrevivientes de sí mismo, podrá sustituir la forma alternativa en la población. La selección natural es la sobrevivencia diferencial de los alelos alternativos.
El individuo puede ser considerado como un vehículo temporal o una máquina de sobrevivencia, a través de la cual los genes sobreviven y se replican (Dawkins, 1976). Puesto que la selección de genes está mediada por el fenotipo, los genes más exitosos serán aquellos que promuevan de manera más efectiva la sobrevivencia y el éxito reproductivo de un individuo. Como resultado se debería esperar que los individuos se comportaran de forma tal, que promovieran su sobrevivencia genética (Krebs y Davies, 1996).
Para conocer más profundamente la biología de los organismos, especialmente de aquellos que de una manera u otra se relacionan con el hombre, hoy en día se requiere un trabajo integral y uno de los aspectos que bien vale la pena estudiar es su comportamiento como una expresión de sus genes. En el caso de los insectos es evidente que la herencia juega un papel muy importante, puesto que la mayoría de ellos tienen un modo de vida solitario, en el cual no tienen oportunidad de aprender de otros adultos cómo deben comportarse. El comportamiento de insectos es un campo de estudio atractivo para los biólogos, pero las bases del comportamiento innato todavía siguen en fase exploratoria por la dificultad de identificar genes responsables de comportamientos particulares (Yamamoto, et al., 1997). Sin embargo, con los progresos en biología molecular, genómica y secuenciación del genoma de A. mellifera (Robinson et al., 2006; Robinson y Weaver, 2006), se cuenta con más herramientas para desentrañar las bases del comportamiento. Esto es particularmente cierto cuando se revisan los avances en el entendimiento de las bases moleculares del comportamiento social.
II. MÉTODOS PARA HACER EL ANÁLISIS GENÉTICO DEL COMPORTAMIENTO
La genética del comportamiento como campo de investigación utiliza diversas metodologías que intentan evidenciar aquellos comportamientos que se creen influidos genéticamente. Tradicionalmente se han usado tres métodos principales para estudiar la forma en que los genes influyen sobre el comportamiento: uso de mutantes génicos, experimentos de selección artificial y, estudio de poblaciones con diferencias genéticas (Benzer, 1973; Krebs y Davies, 1996). Sin embargo recientemente, mediante la transcriptómica se han podido medir cambios de expresión de genes y su relación con cambios comportamentales. Uso de mutantes génicos. En Apis mellifera se han registrado muchos mutantes, pero la mayoría de ellos son de tipo morfológico: color y morfología de ojos (Rothenbuhler et al., 1952), tamaño de las alas (Hachinohe y Onischi, 1953) y color del abdomen (Mackensen, 1951). Soares, 1975 aplicando radiación gamma obtuvo una serie de mutantes, no sola-mente que afectaban color de ojos en abejas africanizadas, sino también que afectaban el aguijón en obreras. La mutación denominada -aguijón bífido-, hace que las piezas que lo componen queden separadas, impidiendo así que las obreras piquen e inyecten su veneno, minimizando así los efectos del comportamiento defensivo de las abejas africanizadas. Al cabo de 11 generaciones de selección se obtuvo una cepa en donde 62% de las obreras eran poseedoras de la mutación.
Experimentos de selección artificial. Se producen líneas seleccionadas a partir de parentales que tienen valores extremos para alguna característica comportamental particular, y que se transmite de generación a generación. Como ejemplo tenemos los trabajos de Stort, 1972, quien consiguió establecer líneas de A. mellifera de diferentes intensidades en su comportamiento defensivo, y el de Rothenbuhler, 1958, quien estableció cepas de abejas higiénicas (resistentes a enfermedades de las crías) y no higiénicas (susceptibles a enfermedades de las crías).
Poblaciones con diferencias genéticas. Poblaciones geográficamente diferentes (de una especie determinada), pueden presentar diferencias morfológicas y comportamentales que reflejan adaptaciones a condiciones ambientales distintas y que pueden estar correlacionadas con diferencias genéticas. Por ejemplo, Rothenbhuler, 1964, analizando líneas endocruzadas de A. mellifera mellifera (abeja alemana) y A. mellifera ligustica (abeja italiana) pudo establecer dos linajes perfectamente diferenciados de abejas mansas (van Scoy) y abejas defensivas (Brown). Con datos de retrocruzamientos, este autor demostró que el comportamiento aguijonear estaba controlado por más de dos loci.
Técnicas moleculares. El desarrollo de técnicas como RAPDS y PCR, permitió inicialmente hacer un mapa genético del genoma de A. mellifera y mapear genes que influencian caracteres como el comportamiento defensivo (Hunt y Page, 1995). Posterior-mente con el secuenciamiento del genoma de A. mellifera (HGSC, 2006) se lograron avances importantes en genética de comportamiento, especialmente en división de labores (Smith et al., 2008): la comunicación química es un aspecto fundamental para la división de labores; tanto la reina como las obreras utilizan gran cantidad de feromonas para coordinar todas sus actividades y su efecto comienza a ser conocido en términos moleculares; cambios en la expresión de genes en el cerebro son inducidos por feromonas que causan cambios rápidos y temporales en el comportamiento (Alaux y Robinson, 2007). La secuenciación del genoma de A. mellifera llevó también al descubrimiento de un sistema de metilación completamente funcional (HGSC, 2006), el primero encontrado en insectos, relacionado con la determinación de castas (Kucharski et al., 2008). Análisis de loci asociados con rasgos cuantitativos (QTL, quantitative trait loci), identificando genes candidatos (genes probables para determinar una característica específica, seleccionados dentro de un grupo de genes con funciones conocidas en otras especies), trasncriptómica (medición de cambio de expresión de genes relacionados con cambios en el fenotipo, mediante micro-arreglos han permitido conocer más acerca de los genes que están involucrados en la variabilidad de los distintos comportamientos en abejas (Hunt et al., 1998; Robinson et al., 2005).
Sin embargo es necesario resaltar algunos puntos:
- a.
- No necesariamente un gene es el responsable de la modificación en una estructura o comportamiento particular: puede haber varios pares de genes influenciando tal comportamiento, especialmente cuando se trata de caracteres cuantitativos. En este caso estaremos hablando de QTL que son loci que controlan caracteres cuantitativos y que muestran una asociación estadística entre marcadores genéticos y fenotipos medibles.
- b.
- Frecuentemente los genes afectan el comportamiento de forma simple (genes que codifican para enzimas que influencian el desarrollo de sistemas sensoriales, nerviosos o motores de un animal, y que a su vez afectan su comportamiento).
- c.
- La forma por la cual un comportamiento se desarrolla, resulta de una interacción compleja entre ambiente y genes, de manera que los genes por si solos no producen el comportamiento.
III. GENÉTICA DEL COMPORTAMIENTO EN ABEJAS
En el mundo existen aproximadamente 20.000 especies de abejas, dentro de las cuales solamente entre 5 y 10% son sociales; el hecho que dentro de este grupo haya tal variabilidad en la organización de sus colonias (desde solitarias, pasando por varios niveles de organización, hasta llegar al más alto que es la eusocialidad), permite hacer estudios comparativos y además aproximarse a los orígenes del comportamiento social. La abeja de miel (género Apis, Familia Apidae) es uno de los organismos utilizados en estudios de comportamiento, debido a su forma de vida social, la cual requiere de coordinación entre todos los individuos de la comunidad; esto a su vez implica comunicación, establecimiento de jerarquías y división de trabajo. La división de trabajo dentro de una colonia de abejas es consecuencia de cambios fisiológicos relacionados con edad de las obreras y con la variación genética entre ellas que hace que realicen diferentes tareas (Page y Robinson, 1991). Dentro de las características de los insectos sociales, se pueden mencionar la existencia de varias generaciones simultáneamente, cuidado cooperativo de los inmaduros, existencia de castas estériles (individuos que no se reproducen dentro de las colmenas) y comportamiento altruista.
Numerosos estudios han mostrado que varios de los comportamientos presentados por las obreras están determinados genéticamente (comportamiento defensivo, comportamiento higiénico) y además que hay variación genética entre poblaciones en el desempeño de tareas como recolección de agua, néctar y polen (Page et al., 2000). Igualmente algunos aspectos del comportamiento social, como control de la reproducción en castas estériles, también están bajo influjo genético (Oldroyd y Osborne, 1999). Gracias a la secuenciación del genoma de A. mellifera se ha podido avanzar, no solo en su estructura, sino también en el conocimiento de genes involucrados en diferentes aspectos del comportamiento de la especie. Se sabe que el genoma de A. mellifera está constituido por 10.000 genes (menos que Drosophila y la mitad de los que tienen los humanos); abejas y humanos comparten 45,7% de sus genes. En comparación con otros insectos, el genoma de la abeja productora de miel se desarrolló muy lentamente y contiene muchos genes para el sentido del olfato. La abeja dispone en su genoma de casi tres veces más receptores olfativos que la mosca de la fruta y el doble que el mosquito transmisor de malaria, pero una séptima parte de los receptores gustativos, es decir, su gusto es mucho peor. Los investigadores creen que se debe a que necesitan un buen sistema olfativo para detectar señales químicas que controlan su comportamiento dentro y fuera de las colmenas y que son la base de su éxito. Además, utilizan su excelente sentido del olfato para comunicación, orientación y búsqueda de alimento. La habilidad que tienen las abejas para diferenciar los sutiles perfumes de las flores tiene su explicación en los 157 receptores olfativos presentes en su genoma (HGSC, 2006).
Comportamiento defensivo. Una de las labores que desempeñan las obreras de A. mellifera es la defensa de la colonia. El primer contacto que mucha gente tiene con la abeja de miel, es precisamente con su comportamiento defensivo, y más aún después de la introducción de la abeja africana (A. mellifera scutellata) en América del Sur y su posterior dispersión a través de la región tropical y subtropical del Nuevo mundo (Kerr, 1967; Spivak, 1991; Guzmán y Page, 1994). Estudios recientes mencionan que el comportamiento defensivo involucra al menos dos tareas diferentes desempeñadas por obreras especializadas: 1. Comportamiento de guardia a la entrada del nido: obreras que vuelan y participan en ataques masivos siguiendo una secuencia básica. 2. Guardianas especializadas: obreras que patrullan la entrada y examinan las abejas entrando al nido y que tienen capacidad de reconocer a sus compañeras por el olor de su cutícula: abejas que no pertenecen al nido son rechazadas y agredidas (Breed et al., 1990; Hunt et al., 2007). Varios son los factores que desencadenan la respuesta de aguijonear: movimientos fuertes, vibraciones del sustrato, feromonas de alarma u otros olores. A partir de la metodología propuesta por Stort, 1970, se observaron líneas de abejas más defensivas (agresivas) y otras menos defensivas (mansas), lo cual hizo evidente la influencia genética sobre el comportamiento defensivo (Ruttner, 1988). Se determinó que ese comportamiento podía subdividirse en varias fases, tomadas desde el momento que la abeja inicia su defensa, el número de aguijones que pueden dejar en un -intruso- (una lámina de cuero o una bola de gamuza), la distancia de persecución, o el número de abejas que salen de la colonia en respuesta a una feromona de alarma; además se observaron fenotipos defensivos diferentes atribuibles a factores genéticos (Kerr et al., 1974; Stort y Goncalvez, 1991). Las líneas más defensivas responden más rápido a cualquiera de los estímulos desencadenantes del comportamiento de defensa. Por medio de cruzamientos entre cepas defensivas y mansas para obtener híbridos (F1) y en algunos casos retrocruzamientos, se determinó la dominancia genética para el carácter -aguijonear- lo cual puede ser ventajoso para abejas en áreas donde hay alto grado de depredación, pero desventajosa para apicultores en las labores de cría y manejo. Diferentes trabajos involucrando cruzamientos entre cepas de abejas europeas, cepas de abejas africanas e híbridos produjeron resultados variables, algunas veces mostrando dominancia y otras veces aditividad (Collins y Rinderer, 1991). Otros trabajos han mostrado correlaciones fenotípicas entre caracteres morfológicos, como tamaño del cuerpo o tamaño de las alas, y comportamiento defensivo; sin embargo, no hay evidencias de correlaciones genéticas (Collins et al., 1994; Quezada-Euan y Paxton, 1999). Hunt et al., 1998, lograron mapear 5 -sting- QTLs basados en la capacidad de las obreras de aguijonear a la entrada del nido. Estudios posteriores mostraron que solamente tres de esos QTLs (sting 1, sting 2 y sting3) afectaban el comportamiento individual de las guardianas (Arechavaleta-Velasco et al., 2003). Sting 1: responsable por el comportamiento de aguijonear y el único asociado con iniciación del ataque a nivel individual; presenta el efecto más fuerte en la variación fenotípica. Sting 2 y Sting 3 son responsables no solo de aguijonear sino de coordinar la respuesta a nivel colonial, frente a estímulos visuales y feromona de alarma (Hunt et al., 2007).
Comportamiento higiénico. Este comportamiento en la abeja A. mellifera se puede definir como la habilidad que tienen las obreras para detectar, desopercular y remover crías enfermas desde la cámara de cría hacia el exterior de la colonia. Muchos autores han demostrado que este comportamiento es un mecanismo natural de resistencia a la loque americana (enfermedad bacteriana que afecta larvas de abejas; Rothenbuhler, 1964; Spivak y Reuter, 1998) y a las enfermedades micóticas como ascosferosis (Milne, 1983; Gilliam et al., 1989; Spivak y Gilliam, 1993) o de origen parasitario como es el caso de Varroa destructor (Rath y Drescher, 1990; Guerra et al., 2000). Algunos programas de crianza han seleccionado este rasgo aumentando la frecuencia del comportamiento higiénico en poblaciones de abejas (Rothenbuhler, 1964; Spivak y Gilliam, 1993; Palacio et al., 2000). Rothenbuhler, 1964, demostró que el comportamiento higiénico de las abejas es controlado por dos genes recesivos independientes: uno responsable de desopercular la cría enferma (gene u) y el otro responsable de remover la cría enferma fuera del nido de cría (gene r). Otros estudios postularon que la determinación genética del comportamiento higiénico podría estar controlada por tres loci (Moritz, 1988). Gramacho, 1999, presentó una nueva hipótesis por la cual el control de este comportamiento se podría explicar por tres genes recesivos (d1/d1, d2/d2 = destapando y r/r = retiro). Sin embargo, no se ha probado ninguna de las dos hipótesis. Recientemente, Lapidge et al., 2002, con técnicas moleculares, han sugerido que son siete genes los que están implicados en comportamiento higiénico. Por otra parte, se ha determinado que las abejas higiénicas poseen mayor sensibilidad olfatoria y son capaces de discriminar entre crías normales y anormales a una baja intensidad de estímulo, detectando y removiendo crías enfermas, muertas o parasitadas de una manera rápida y eficiente (Gramacho y Spivak, 2003).
Comportamiento de forrajeo. La división de labores en abejas es uno de los aspectos importantes y característicos de su vida social. Hay dos actividades generales dentro de una colonia de A. mellifera: obreras que se dedican a labores al interior del nido y aquellas que salen a recolectar recursos alimenticios (néctar y polen); estas dos actividades están muy relacionadas porque por un lado siguen una secuencia etaria (las abejas mas jóvenes están dentro del nido y las más viejas salen a forrajear) y por otro, en líneas altamente seleccionadas para almacenamiento de polen, es más probable que las obreras salgan a forrajear precozmente y que posteriormente se dediquen a recolección de polen, que en líneas seleccionadas para poca recolección de polen (Page et al., 1998). Con base en análisis del comportamiento de forrajeo tanto a nivel de colonia total, como a nivel individual, se detectaron tres QTL para forrajeo de polen: Pln1, Pln2 y Pln3; Pln1 está asociado con el tamaño de las cargas corbiculares colectadas por las obreras; Pln2, también asociado con el tamaño de cargas corbiculares, pero, junto con Pln 3, influye en la discriminación de la concentración de azúcares del néctar de las flores (Hunt et al., 1995; Page et al., 2000). Un cuarto QTL (Pln4) se encontró como responsable por la respuesta a la sucrosa, pero también sobre la edad de inicio del forrajeo (Rueppell et al., 2004; Rueppell et al., 2006).
Comportamiento social. Quizás el área más interesante del estudio del comportamiento en abejas sea acerca de su vida en sociedad. La regulación del comportamiento social ha comenzado a conocerse, y como resultado ha nacido una nueva disciplina la sociogeonómica, que se enfoca en la determinación de los genes que regulan comportamiento social, su función particular, sus relaciones con el ambiente que los rodea y de esta manera estudiar la evolución de la diversidad del comportamiento (Robinson et al., 2008). Las abejas sociales, como A. mellifera son excelentes modelos para estudios sociogenómicos porque presentan distintas formas de sociabilidad, sistema haplodiploide de determinación de sexo, genoma secuenciado y facilidad de manipulación de colonias (Smith et al., 2008). En una colonia de A. mellifera la determinación de castas es un factor decisivo en la regulación del comportamiento social. La diferenciación en reinas y obreras se lleva a cabo por nutrición diferencial. Todas las larvas diploides se desarrollan como hembras y, pueden llegar a ser reinas u obreras. Las larvas menores de dos días son alimentadas con jalea real (sustancia proteínica producida en las glándulas hipofaringeanas de las obreras); a partir del tercer día de vida las larvas modifican su dieta hacia una mezcla de jalea real, miel y polen, con lo cual su desarrollo será hacia obrera; pero las larvas destinadas a ser reinas son alimentadas solamente con jalea real. Se han encontrado diferencias importantes en la expresión de muchos genes en esos dos tipos de larvas (Jordá y Peinado, 2009), lo cual se va a reflejar en el fenotipo y comportamiento de las hembras resultantes. Pero la expresión génica no depende solamente de factores de transcripción, sino que también están involucrados factores epigenéticos (factores no genéticos que intervienen en la regulación de la expresión génica sin cambio en la secuencia de nucleótidos); es así como la determinación de castas (Kucharski et al., 2008) y división de trabajo dependen de programas de expresión génicas que cambian a lo largo de su desarrollo; además, los genes se expresan o no, según se den ciertas condiciones bioquímicas como metilación de ADN, una de las marcas epigenéticas mejor caracterizadas ( Jordá y Peinado, 2009).
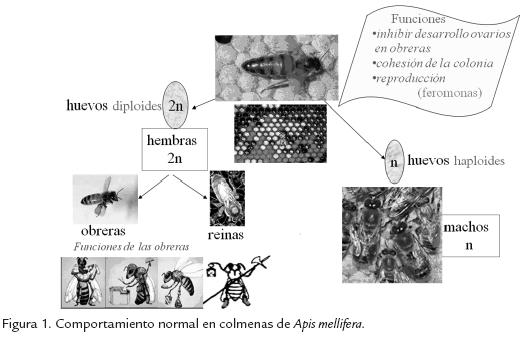
La reina, por medio de su feromona mandibular, desempeña un papel crucial en la regulación social pues inhibe el desarrollo de los ovarios de las obreras, influye en la edad a la cual las obreras cambian su función de tareas dentro de la colmena a tareas fuera de la colmena, evita la construcción de celdas reales (producción de más reinas) y muchos otros aspectos necesarios para el buen funcionamiento de la sociedad. Nelson et al., 2007, demostraron que genes que afectan procesos fisiológicos, como el gene vitellogenina, tienen efectos pleiotrópicos sobre caracteres de la vida social como inicio de forrajeo y longevidad de las obreras. En la Tabla. 1 se resumen algunos hallazgos obtenidos a través de estudios moleculares y que involucran distintos aspectos del comportamiento social en abejas.
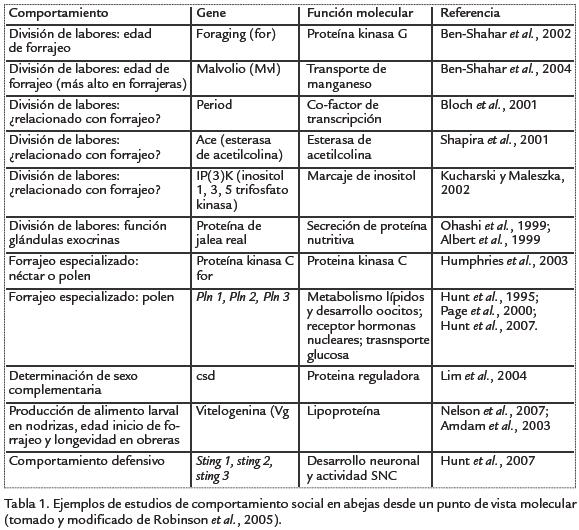
Altruismo y anarquía en abejas
Altruismo. A pesar de que las obreras de las abejas sociales tienen ovarios funcionales, no ponen huevos. Esto se ha interpretado como una forma de altruismo, puesto quees una acción que reduce la probabilidad de reproducción de un individuo particular en beneficio de otro u otros; en el caso particular de las abejas sociales se manifiesta cuando las obreras colaboran en el cuidado de las crías de sus hermanas y sacrifican la vida en defensa de la colonia, en cambio de tener sus propios hijos.
¿Por qué la selección natural, que generalmente favorece el éxito reproductivo de los individuos, hace que ellos ayuden a otros a costa de su propia reproducción? Desde que Hamilton, 1964, propusiera su teoría de -inclusive fitness- en la cual la selección natural favorece a individuos que presentan comportamiento altruista hacia otros genéticamente relacionados, esta ha sido la respuesta más aceptada; se supone que si en una sociedad de insectos, un individuo ayuda a otro relacionado genéticamente, a criar sus hijos, esto es un acto voluntario que beneficiaría al individuo altruista. En colmenas de A. mellifera (cuya reina ha sido fecundada por un solo macho) las obreras hijas comparten 50% de sus genes con la madre, pero entre hermanas esta relación se incrementa (75%), esto quiere decir que les resulta genéticamente más ventajoso ayudar a criar los hijos de sus hermanas que tener sus propios hijos (Hamilton, 1964). Sin embargo lo normal es que las reinas de A. mellifera se apareen con más de 10 machos, con lo cualel parentesco entre obreras se reduce drásticamente (varias subfamilias, cada una constituida por la reina, cada uno de los machos que la fecundaron y sus hijas), pero aún en esas condiciones, donde la relación genética entre las obreras es muy baja, el altruismo prevalece; se podría suponer que con un porcentaje de parentesco bajo entre obreras, el sistema altruista se debilite y surjan obreras que quieran poner su propios huevos Pero en la realidad, tan solo una entre 1.000 obreras intenta poner sus propios huevos (Ratnieks, 1993). Esto sucede porque existe presión social o coersión por parte de obreras vigilantes, que va desde la eliminación de los huevos puestos por obreras hasta ataques a las mismas obreras ponedoras; publicaciones recientes muestran que entonces el altruismo no es voluntario, sino obligado; las obreras renuncian a su propia reproducción debido a que las obreras vigilantes las castigan si no lo hacen (Ratnieks y Wenselers, 2007). Así, la coerción social ayuda a explicar el altruismo de las obreras y la cooperación en las sociedades de insectos sociales.
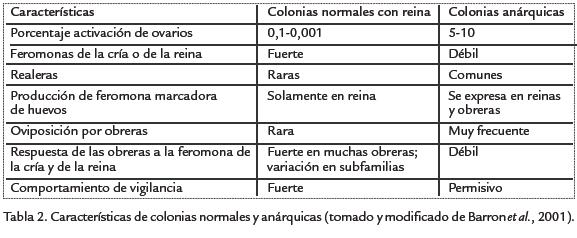
Comportamiento anárquico. En condiciones normales una colonia de A. mellifera está constituida por una reina (sistema monogínico), cuyas funciones principales son pos-tura de huevos, mantenimiento de la cohesión de la colonia, inhibición del desarrollo de ovarios en obreras y producción de realeras (nacimiento de más reinas). Por su parte, las obreras realizan las demás actividades tendientes al buen funcionamiento de la colonia: cuidado de crías (nodrizas), limpieza de la colonia, construcción, consecución de alimento (forrajeras) y vigilancia y protección de la colonia (abejas guardianas). Todas estas actividades están mediadas por feromonas producidas por la reina, los inmaduros y las mismas obreras. Es importante recordar, que además del sistema de determinación de castas (obrera y reina), una de las características fundamentales de la sociedad de abejas y de otros Hymenoptera, es la determinación de sexo. La reina de las abejas sociales y las hembras de abejas solitarias tienen la posibilidad de poner dos tipos de huevos: haploides (n), sin fecundar de los cuales nacen machos, por supuesto, haploides y huevos diploides (2n) fecundados, que van a dar origen a hembras diploides (Fig. 1).
En colonias con reina, las obreras no pueden poner huevos no solo por el efecto inhibidor de la feromona real, sino también por la actividad de las obreras vigilantes; en abejas europeas (A. mellifera) tan solo 1% de obreras tienen ovarios activos y pueden poner huevos (Vischer, 1996). En colonias sin reina (huérfanas), la reproducción de las obreras es muy común; algunas de ellas inician posturas y se tornan en ponedoras con la consiguiente producción de machos hijos de obreras; pero esas colonias huérfanas están destinadas a desaparecer puesto que poco a poco las obreras van muriendo y solo quedan machos; además se vuelven vulnerables a parasitismo por ausencia de obreras vigilantes (Nanork et al., 2005). Sin embargo, ocasionalmente en colmenas con reina, surgen obreras que escapando del influjo de la reina ponen sus propios huevos, haploides, con la consiguiente producción de machos hijos de obreras. Se han encontrado algunas colonias naturales de A. mellifera en las cuales el éxito reproductivo de las obreras es muy alto (evidenciado por la gran producción de descendencia de machos hijos de obreras). Las así llamadas colmenas anárquicas naturales, son muy raras; las primeras fueron encontradas en apia-rios de Nueva Zelanda en 1987; posteriormente fueron detectadas más colmenas en otros países como Estados Unidos (California) y Gran Bretaña (Barron et al., 2001). Algunas de estas colonias se han estudiando usando marcadores genéticos (Oldroyd et al., 1994; Montague y Oldroy, 1998). Se descubrió que la actividad reproductiva de obreras está confinada a unas pocas subfamilias dentro de una colmena, lo cual es indicativo de que este comportamiento está influenciado genéticamente, además de estar relacionado con factores ambientales (Hoover et al., 2006).
Generalmente, el sistema de feromonas y la acción de obreras vigilantes mantienen la división de labores reproductiva en colonias con reina. Sin embargo, todos los sistemas cooperativos tienen una mezcla de genotipos cooperativos (altruistas) y genotipos corruptos (anárquicos). En las sociedades humanas la anarquía se refiere al quiebr de un orden social establecido y ausencia de un control policial. De esta misma forma se puede aplicar en sociedades de insectos, particularmente abejas sociales en donde la presencia de muchas obreras con ovarios activos indica ruptura, pérdida o disminución de las señales que mantienen la esterilidad en obreras, y ausencia de vigilancia (Beekman y Oldroyd, 2008), así la reproducción de las obreras se torna mucho más común. Este comportamiento es contrastante con la situación normal en colmenas con reina y puede ser una estrategia utilizada por las obreras para intentar alcanzar su éxito reproductivo individual. Cuando las obreras vigilantes detectan obreras anárquicas utilizan dos mecanismos para reprimirlas: oofagia de huevos puestos por ellas y agresión directa. Las vigilantes detectan los huevos de obreras porque las reinas producen una feromona con la que marcan sus huevos (Ratnieks, 1995) y los hace diferentes a los de las obreras ponedoras. El síndrome de colonias anárquicas esta fuertemente influenciado por factores genéticos y que se genera en una simple subfamilia dentro de una colonia (Oldroyd et al., 1994; Montague y Oldroyd, 1998). En esas colonias, muchas obreras activan sus ovarios y producen hijos; en líneas seleccionadas para el anarquismo no hay inhibición de ovarios de obreras, las obreras ponen huevos en presencia de la reina, las obreras anárquicas evaden a las -obreras vigilantes- de manera que sus huevos son removidos lentamente o no son removidos (quizás esto sea debido a que las obreras anárquicas pueden -falsificar- la feromona marcadora de huevos de la reina; Ratnieks, 1992) y hay disminución de la vigilancia. En la Tabla. 2 se muestran características comparativas con colmenas normales.
Actualmente es posible producir colmenas anárquicas seleccionadas mediante inseminación de reinas normales con semen de machos normales hijos de reina y machos hijos de obreras anárquicas (Oldroyd y Osborne, 1999). A partir de cruzamientos como esos se evidenció que si bien en la primera generación no surgieron machos hijos de obreras anárquicas, en la segunda generación se obtuvieron cuatro clases de fenotipos (Oldroyd y Osborne, 1999):
1. Sin activación de ovarios en las obreras.
2. Activación de ovarios en las hijas de machos anárquicos, pero no se detectaron posturas de huevos ni producción de inmaduros.
3. Activación de ovarios en obreras hijas de machos anárquicos y de machos normales, pero sin producción de hijos.
4. Activación de ovarios y producción de hijos por obreras descendientes de machos anárquicos.
Estos resultados permitieron concluir que hay por lo menos dos pares de genes involucrados en el carácter anárquico, que segregan independientemente: activación de ovarios y habilidad para poner huevos que no son vigilados (Oldroyd y Osborne, 1999). El comportamiento anárquico solamente aparece cuando las obreras tienen las dos características: ovarios activos en colonias con reinas y postura de huevos que no son vigilados y pueden desarrollarse en machos (clase 4). Un tercer par de genes sería uno que induciría a alterar el comportamiento de las obreras vigilantes, las cuales serian más permisivas. A pesar que el anarquismo puede ser favorable a nivel individual, no lo es a nivel colonial. Las obreras anárquicas probablemente reducen la productividad de la colonia debido a varios factores: producción excesiva de machos, que no forrajean y descuido en el desarrollo de otras tareas importantes para la colonia como forrajeo, construcción de celdas, almacenamiento de alimento, atención a crías, disminución de cohesión social; sin embargo, este es un aspecto que todavía no esta lo suficientemente claro.
CONCLUSIONES
Gracias a las recientes herramientas proporcionadas por las técnicas moleculares modernas se ha podido determinar que el comportamiento de las abejas es producto de sus genes, ambientes ecológicos y fisiológicos, condiciones sociales de la colonia e interacción entre estos factores. La sociogenómica, es una nueva disciplina que se enfoca en la determinación de los genes que regulan el comportamiento social, su función particular, sus relaciones con el ambiente que los rodea y de esta manera estudiar la evolución de la diversidad del comportamiento. Con la secuenciación del genoma de A. mellifera se ha podido avanzar, no solo en su estructura, sino también en el conocimiento de genes involucrados en diferentes aspectos del comportamiento de la especies. Se sabe que el genoma de A. mellifera está constituido por 10.000 genes (menos que Drosophila y la mitad de lo que tienen los humanos); abejas y humanos comparten 45,7% de sus genes. En comparación con otros insectos, el genoma de la abeja productora de miel se desarrolló muy lentamente y contiene muchos genes para el sentido del olfato, los cuales son muy importante para la organización de la vida diaria de las abejas (HGSH, 2006). Los análisis genéticos del comportamiento de las abejas se han visto reforzados por los análisis genómicos (¿cuáles son las vías moleculares que están involucradas en la variación genética del comportamiento?), lo que ha permitido entender, por ejemplo, el funcionamiento molecular del comportamiento defensivo, comportamiento higiénico, la compleja y diversa división de labores entre obreras, la determinación de casta y sexo y comportamientos tan controvertidos como los generados por la diversidad genética dentro de las colonias, que a la par que ocasiona conflictos entre sus miembros, sin embargo incrementa su funcionamiento. (Smith et al., 2008). Otro aspecto importante en el estudio de la regulación genética del comportamiento en abejas, y que según Jordá y Peinado, 2009, es el paso a seguir después de la secuenciación del genoma, es el conocimiento de los mecanismos epigenéticos, los cuales son un puente entre ambiente y genética.
AGRADECIMIENTOS
A la Universidad Nacional de Colombia por proporcionar el espacio y el tiempo profundizar en el interesante mundo de las abejas. A los revisores anónimos y a los editores de este suplemento por su paciente labor. A Joanna Jaramillo, por su colaboración en organización de bibliografía
BIBLIOGRAFÍA
ALAUX C, ROBINSON GE. Releaser Pheromone Provokes Immediate-early Gene and Slow Behavioral Response. J Chem Ecol. 2007;33:1346-1350.
[ Links ]ALBERT S, BHATTACHARYA D, KLAUDINY J, SCHMITZOVA J, SIMUTH J. The Family of Major Royal Jelly Proteins and its Evolution. J Mol Evol. 1999;49:290-297.
[ Links ]AMDAM GV, NORBERG K, HAGEN A, OMHOLT SW. Social Exploitation of Vitellogenin. Proc Natl Acad Sci U S A. 2003;100:1799-1802.
[ Links ]ARECHAVALETA-VELASCO ME, HUNT GJ, EMORE C. Quantitative Trait Loci that Influence the Expression of Guarding and Stinging Behaviours of Individual Honey Bees. Behav Genet. 2003;33:357-364.
[ Links ]BARRON AB, OLDROYD BP, RATNIEKS FLW. Worker Reproduction in Honey-bees (Apis) and the Anarchic Syndrome: A Review. Behav Ecol Sociobiol. 2001;50:199-208.
[ Links ]BEEKMAN M, OLDROYD B. When Workers Disunite: Intraespecific Parasitism by Eusocial Bees. Annu Rev Entomol. 2008;53:19-37.
[ Links ]BEN-SHAHAR Y, ROBICHON A, SOKOLOWSKI MB, ROBINSON GE. Influence of Gene Action Across Different Time Scales on Behaviour. Science. 2002;296:741-744.
[ Links ]BEN-SHAHAR Y, DUDET NI, ROBINSON GE. Phenotypic Deconstruction Reveals Involvement of Manganese Transporter Malvolio in Honey Bee Division of Labor. J Exp Biol. 2004;207:3281-3288.
[ Links ]BENZER S. Genetic Dissection of Behaviour. Sci Am. 1973;229:24-37.
[ Links ]BLOCH G, TOMA DP, ROBINSON GE. Behavioral Rhythmicity, Age, Division of Labor and Period Expression in the Honeybee Brain. J Biol Rhythms. 2001;16:444-456.
[ Links ]BREED MD, ROBINSON GE, PAGE RE. Division of Labor During Honey Bee Colony Defense. Behav Ecol Sociobiol. 1990;27:395-401.
[ Links ]COLLINS A, DALY H, RINDERER T, HARBO J, HOELMER K. Correlations between Morphology and Colony Defence in Apis mellifera. J Apicult res. 1994;33:3-10.
[ Links ]COLLINS AM, RINDERER TE. Genetics of Defensive Behaviour I. En: Spivak M, Fletcher JDC, Breed MD, editors. The -African- Honey Bee. Westview. Boulder, CO; 1991; p.309-328.
[ Links ]DAWKINS R. The Selfish Gene. Oxford: Oxford University Press; 1976.
[ Links ]GILLIAM M, TABER S, LORENZ BGJ, PREST DB. Hygienic Honey Bees and Antagonistic Normal Microflora for Control of Chalkbrood Disease. XXXII International Apiculture Congress of Apimondia. Rio de Janeiro, Brazil. 1989;22-28.
[ Links ]GRAMACHO KP. Fatores que interferem no comportamento higiênico das abelhas Apis mellifera. [tesis de doctorado]. São Paulo: Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, USP; 1999.
[ Links ]GRAMACHO KP, SPIVAK M. Differences in Olfatory Sensitivity and Behavioral Responses Among Money Bees Bred for Hygienic Behavior. Behav Ecol Sociobiol. 2003;54:472-479.
[ Links ]GUERRA JR JCV, GONÇALVES LS, DE JONG D. Africanized Honey Bees (Apis mellifera L.) are more Efficient at Removing Worker Brood Artificially Infested with the Parasitic Mite Varroa jacobsoni Oudemans than are Italian Bees or Italian/Africanized hybrids. Genet Mol Biol. 2000;23:89-92.
[ Links ]GUZMÁN-NOVOA E, PAGE JR RE. The Impact of Africanized Bees on Mexican Beekeeping. Am Bee J. 1994;134:101-106.
[ Links ]HACHINOHE Y, ONISCHI N. Sobre a nova mutação -asa rudimentar- na abelha. Bull do Inst Cien Agr. 1953;7:139-145.
[ Links ]HAMILTON WD. The Genetical Evolution of Social Behaviour. I & II. J Theor Biol. 1964;7:1-52
[ Links ]HGSH (THE HONEYBEE GENOME SEQUENCING CONSORTIUM). Insights into Social Insects from the Genome of the Honeybee Apis mellifera. Nature. 2006;443(7114):931-949 NIH Public Access
[ Links ]HOOVER SE, HIGO HA, WINSTON ML. Worker Honey Bee Ovary Development: Seasonal Variation and the Influence of Larval and Adult Nutrition. J Comp Physiol B. 2006;176(1):55-63.
[ Links ]HUMPHRIES MA, MULLER U, FONDRK MK, PAGE JR RE. PKA and PKC Content in the Honeybee Central Brain Differs in Genotypic Strains with Distinct Foraging Behavior. J Comp Physiol A. 2003;189:555-562.
[ Links ]HUNT GJ, AMDAM GV, SCHLIPALIUS D, EMORE C, SARDESAI N, WILLIAMS CE, et al. Behavioral Genomics of Honeybee Foraging nd Nest Defense. Naturwissenschaften. 2007;94:247-267.
[ Links ]HUNT GJ, GUZMÁN-NOVOA E, FONDRK MK, PAGE JR RE. Quantitative Trait Loci for Honey Bee Stinging Behavior and Body Size. Genetics. 1998;148:1203-1213.
[ Links ]HUNT GJ, PAGE JR RE. Linkage Map of the Honey Bee, Apis mellifera, Based on RAPD Markers. Genetics. 1995;139:1371-1382.
[ Links ]HUNT GJ, PAGE JR RE, FONDRK MK, DULLUM CJ. Major Quantitative Trait Loci Affecting Honey Bee Foraging Behavior. Genetics. 1995;141:1537-1545.
[ Links ]JORDÁ M, PEINADO M. La regulación génica del comportamiento social de las abejas. Investigación y Ciencia. 2009;40-43.
[ Links ]KERR WE, BLUM MS, PISANI JF, STORT AC. Correlation between Amounts of 2- Heptanome and Iso-amyl-acetate in the Honeybees and their Agressive Behavior. J Apic Res. 1974;13:173-176.
[ Links ]KERR WE. The History of the Introduction of African Bees to Brazil. South Afr Bee J. 1967;39:3-5.
[ Links ]KREBS JR, DAVIES NB. Introdução à Ecologia comportamental. Sao Paulo: Ed. Atheneu; 1996.
[ Links ]KUCHARSKI R, MALESZKA R. Molecular Profiling of Behavioural Development: Differential Expression of mRNAs for Inositol 1,4,5-trisphosphate 3-kinase isoforms in Naïve and Experienced Honeybees (Apis mellifera). Mol Brain Res. 2002;99:92-101.
[ Links ]KUCHARSKI R, MALESZKA J, FORET S, MALESZKA R. Nutritional Control of Reproductive Status in Honeybees Via DNA Methylation. Science 2008;319:1827-1830.
[ Links ]LAPIDGE KL, OLDROYD BP, SPIVAK M. Seven Suggestive Quantitative Loci Influence Hygienic Behavior of Honey Bees. Naturwissenschaften. 2002;89:565-568.
[ Links ]LIM MM, WANG Z, OLAZÁBAL DE, REN X, TERWILLIGER EF, YOUNG LJ. Enhanced Partner Preference in a Promiscuous Species by Manipulating the Expression of a Single Gene. Nature. 2004;429:754-757.
[ Links ]MACKENSEN O. Viability and Sex Determination in the Honey Bee (Apis mellifera L.). Genetics. 1951;36:500-509.
[ Links ]MILNE CP. Honey Bee (Hymenoptera: Apidae) Hygienic Behavior and Resistance to Chalk Brood. Annals of Entomological Society of America. 1983;76:384-387.
[ Links ]MORITZ RFA. A Re-evaluation of the Two Locus Model for Hygienic Behavior in Honeybees (Apis mellifera L.). J Hered. 1988;79:257-262.
[ Links ]MONTAGUE CE, OLDROYD BP. The Evolution of Worker Sterility in Honey Bees: An Investigation Into a Behavioural Mutant Causing Failure of Worker Policing. Evolution. 1998;52:1408-1415.
[ Links ]NANORK P, PAAR J, CHAPMAN NC, WONGSIRI S, OLDROYD BP. Asian Honey Bees Parasitize the Future Dead. Nature. 2005;437:829.
[ Links ]NELSON M, IHLE KE, FONDRK MK, PAGE R, AMDAM G. The gene Vitellogenin has Multiple Coordinating Effects on social Organization. PLoS Biol. 2007;5:673-677.
[ Links ]OHASHI K, NATORI S, KUBO T. Expression of Amylase and Glucose Oxidase in the Hypopharyngeal Gland with an age-dependent Role Change of the Worker Honeybee (Apis mellifera L). Eur J Biochem. 1999;265:127-133.
[ Links ]OLDROYD BP, OSBORNE KE. The Evolution of Worker Sterility in Honeybees: The Genetic Basis of Failure of Worker Policing. Proc R Soc London B. 1999; 266:1335-1339.
[ Links ]OLDROYD BP, SMOLENSKI AJ, CORNUET JM, CROZLER RH. Anarchy in the Beehive. Nature. 1994;371:749.
[ Links ]PAGE JR RE, ERBER J, FONDRK MK. The Effect of Genotype on Response Thresholds to Sucrose and Foraging Behavior of Honeybees (Apis mellifera L.). J Comp Physiol A. 1998;182:489-500.
[ Links ]PAGE JR RE, FONDRK MK, HUNT GJ, GUZMÁN-NOVOA E, HUMPHRIES MA, NGUYEN K, et al., Genetic Dissection of Honeybee (Apis mellifera L.) Foraging Behavior. J Hered. 2000;91:474-479.
[ Links ]PAGE JR RE, ROBINSON GE. The Gnetics of Division of Labor in Honey Bee Colonies. Adv insect physiol. 1991;117-169.
[ Links ]PALACIO MA, FIGINI E, RODRIGUEZ E, RUFINENGO S, BEDASCARRASBURE E, DEL HOYO M. Changes in a Population of Apis mellifera Selected for its Hygienic Behaviour. Apidologie. 2000;31:471-478.
[ Links ]QUEZADA-EUAN, JJG, PAXTON RJ. Rapid Intergenerational Changes in Morphology and Behaviour in Colonies of Africanized and European Honey Bees (Apis mellifera) from Tropical Yucatan, Mexico. J Apicult Res. 1999;38:93-104.
[ Links ]RATH W, DRESCHER W. Response of Apis cerana Fab. Towards Brood Infested with Varroa jacobsoni Oud. and Infestation Rate of Colonies in Thailand. Apidologie. 1990;21:311-321.
[ Links ]ROBINSON GE, EVAN JD, MALESZKA R, ROBERTSON HM, WEAVER DB, WORLEY K, et al., Sweetness and Light: Illuminating the Honey Bee Genome. Insect Mol Biol. 2006;15:535-539.
[ Links ]ROBINSON GE, GRÖZINGER CM, WHITFIELD CW. Sociogenomics: Social Life in Molecular Terms. Nat Rev Genet. 2005;6:257-270.
[ Links ]ROBINSON GE, FERNALD R, CLAYTON D. Genes and Social Behavior. Science. 2008;322:896-899.
[ Links ]ROBINSON G, WEAVER D. The Honey Bee Genome Project: A Model of Cooperation between Academia, Government, and Industry. Am Bee J. 2006;870.
[ Links ]RATNIEKS FLW. Evidence for an Egg Marking Pheromone in the Honey Bee. Am Bee J. 1992;132:813.
[ Links ]RATNIEKS FLW. Egg-laying, Egg-removal, and Ovary Development by Workers in Queenright Honey-bee Colonies. Behav Ecol Sociobiol. 1993;32:191-198.
[ Links ]RATNIEKS FLW. Evidence for a Dueen-produced Egg-marking Pheromone and its Use in Worker Policing in the Honey Bee. J Apicult Res. 1995;34:31-37.
[ Links ]RATNIEKS FLW, WENSELEERS T. Altruism in Insect Societies and Beyond: Voluntary or Enforced?. Trends Ecol Evol. 2007;23:45-52.
[ Links ]ROTHENBUHLER WC. Genetics of a Behaviour Difference in Honey Bees. Proc 10th Inter Cong of Genetics. 1958;11:242.
[ Links ]ROTHENBUHLER WC, GOWEN JW, PARK OW. Five Mutant Genes in Honey Bees (Apis mellifera). Genetics. 1952;37:620-621.
[ Links ]ROTHENBUHLER WC. Behavior Genetics of Nest Gleaning in Honey Bees. IV. Responses of F1 and Backcross Generations to Disease-killed Brood. Am Zoologist. 1964;4:111-123.
[ Links ]RUEPPELL O, PANKIW T, PAGE RE. Pleiotropy, Epistasis and New QTL: The Genetic Architecture of Honeybee Foraging Behavior. J Hered. 2004;95:481-491.
[ Links ]RUEPPELL O, CHANDRA SBC, PANKIW T, FONDRK MK, BEYE M, HUNT G, et al., The Genetic Architecture of Sucrose Responsiveness in the Honeybee (Apis mellifera L.). Genetics. 2006;172:243-251
[ Links ]RUTTNER F. Biogeography and Taxonomy of Honey Bees. Berlin: Springer-Verlag; 1988.
[ Links ]SHAPIRA M, THOMPSON CK, SOREQ H, ROBINSON GE. Changes in Neuronal Acetylcholinesterase Gene Expression and Division of Labor in Honeybee Colonies. J Mol Neurosci. 2001;17:1-12.
[ Links ]SMITH CR, TOTH AL, SUAREZ AV, ROBINSON GE. Genetic and Genomic Analyses of the Division of Labour in Insect Societies. Nat Rev Genet. 2008;9:738-748.
[ Links ]SOARES AEE. Efeitos da radiação gama de Cobalto-60 em Apis mellifera-(Dissertação de Mestrado). Facultad de Medicina de Ribeirão Preto, Universidade de São Paulo, Brasil; 1975.
[ Links ]SPIVAK M, REUTER GS. Performance of Hygienic Honey Bee Colonies in Commercial apiary. Apidologie. 1998;29:291-302.
[ Links ]SPIVAK M. The Africanization Process in Costa Rica. En Spivak M, Fletcher JDC, Breed MD, editors. The African Honey bee Westviwe; Boulder CO; 1991. p. 137-155.
[ Links ]SPIVAK M, GILLIAM M. Facultative Expression of Hygienic Behaviour of Honey Bees in Relation to Disease Resistance. J Api Res. 1993;32:147-157.
[ Links ]STORT AC. Metodologia para o estudo da agressividade de Apis mellifera. . I Congresso Brasileiro de .Apicultura. 1970;36-51
[ Links ]STORT AC. Relações entre caracteres do comportamento agressivo e caracteres morfológicos de abelhas do genero Apis. Hom. a Warwick Esteban Kerr. 1972;275-283.
[ Links ]STORT AC, GONÇALVES LS. Genetics of Defensive Behavior II. En: Spivak M, Fletcher JDC, Breed MD, editors. The -African- Honey Bee. Westview; Boulder CO; 1991; p. 329-356.
[ Links ]VISCHER PK. Reproductive Conflict in Honey Bee: A Stalemate of Worker Egg-laying and Policing. Behav Ecol Sociobiol. 1996;39:237-244.
[ Links ]YAMAMOTO D, JALLON J, KOMATSU A. Genetic Dissection of Sexual Behavior in Drosophila melanogaster. Annu Rev Entomol. 1997;42:551-585.
[ Links ]
