











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
El declive global de la biodiversidad incrementa la necesidad de entender cada vez mejor la relación que existe entre diversidad y provisión de los servicios ecosistémicos, ya que estos son esenciales para la supervivencia humana (Aizen et al., 2009). Los ecosistemas naturales contribuyen al bienestar humano a través de una variedad de funciones ecológicas, que son definidas como la capacidad de proveer servicios que mantienen un ecosistema y benefician a la sociedad (De Groot et al., 2002). De esta manera, la polinización es una función ecológica clave para la producción de biomasa, la reproducción de las plantas y la producción de alimento. La polinización implica la transferencia de polen co-específico entre los órganos reproductores de las flores, lo cual posibilita la fecundación de la planta, y la posterior producción de frutos y semillas. Los insectos himenópteros, específicamente las abejas, son considerados los principales polinizadores de plantas silvestres y de cultivo (Kevan et al., 1990; Kluser et al., 2007). Se estima que el 35 % de las plantas consumidas por el hombre dependen especialmente de himenópteros para su polinización (Klein et al., 2007; Potts et al., 2010). Así, la polinización como función ecológica tiene incidencia directa en el bienestar de la humanidad. No obstante, desde el 2004 se ha documentado el declive progresivo de poblaciones y especies de himenópteros en los sistemas de cultivo y áreas naturales a nivel global (Gallai et al., 2009; Nicholls y Altieri, 2013). Esta disminución en la abundancia y riqueza de polinizadores advierte sobre una potencial crisis en la polinización (Klein et al., 2007), que se traduciría también en una crisis alimentaria. Sin embargo, a pesar de la relevancia de la polinización, existe poco conocimiento sobre las redes de polinizadores y sus dinámicas en la polinización de comunidades de plantas de cultivo en regiones tropicales (Garibaldi et al., 2013).
Las redes de interacción ecológica describen la estructura de comunidades biológicas a partir de índices que permiten medir el papel ecológico de cada una de las especies interactuantes (Bascompte y Jordano, 2006). La representación gráfica de la red incluye nodos, que son las especies de los dos grupos tróficos interactuantes, los cuales están interconectados por medio de enlaces. Según el tipo de interacción, pueden existir redes antagónicas que describen relaciones de depredador-presa (e.g. herviboría, parasitismo) y redes mutualistas, que incluyen interacciones benéficas para ambas partes (e.g. polinización y dispersión de semillas) (Ings et al., 2009). Se ha encontrado que las redes de polinización son altamente estructuradas y que la fuerza de las interacciones contribuye a entender el nivel de interdependencia de las especies que interactúan (Bascompte y Jordano, 2007). Igualmente, se ha establecido que las redes de polinización exhiben altos niveles de especialización y complementariedad funcional (Blüthgen y Klein, 2011). A pesar de la relevancia del estudio de redes y su potencial aplicación a la productividad agrícola, pocas veces se ha cuantificado la red de polinización en agroecosistemas. El enfoque de redes de interacción ofrece la oportunidad de cuantificar el nivel de interdependencia de las especies que hacen posible la productividad y sostenibilidad de los agroecosistemas.
Los agroecosistemas son concebidos como ecosistemas de producción y no sólo como parcelas productivas (i.e. visión reduccionista de la agricultura industrial), ya que este tipo de manejo promueve la diversificación biológica de los cultivos, elimina el uso de agrotóxicos y fomenta la permanencia de funciones ecológicas como la polinización y la dispersión de semillas (Altieri, 2002; Francis et al., 2003). De esta forma, los agroecosistemas tienen un manejo diferente a la agricultura industrial, el cual va dirigido a promover la sostenibilidad ecológica. Los agroecosistemas favorecen el mantenimiento de las relaciones entre especies de diversas taxa. Estas relaciones son de vital importancia dado que influencian la capacidad de coexistencia de las especies y dan estructura y estabilidad a las comunidades biológicas (Waser et al., 1996; Blüthgen y Klein, 2011).
Dentro de los patrones que surgen de las interacciones planta-polinizador se destacan la especialización ecológica, la complementariedad y la redundancia funcional. La especialización ecológica se define como el nicho ecológico restringido para cada especie en términos del uso de los recursos y/o hábitats disponibles; es decir, que un especialista utiliza un menor rango de los recursos, en comparación a un generalista (Devictor et al., 2010). La complementariedad funcional se refiere al efecto que las especies con diferentes nichos generan en una función ecológica particular. Así, la interacción conjunta de las especies tiene mayor efecto en una función ecológica que cuando las especies actúan de manera independiente (Blüthgen y Klein, 2011). En cambio, la redundancia funcional implica que las especies con un nicho similar contribuyen de manera equivalente a una función ecológica y pueden ser mutuamente sustituibles (Blüthgen y Klein, 2011). Sin embargo, estos tres conceptos se relacionan entre sí, pues la complementariedad funcional requiere de cierto nivel de especialización y la redundancia se asocia con un alto nivel de generalización de las especies de una comunidad (Blüthgen y Klein, 2011). Por lo anterior, la cuantificación de estos patrones ecológicos en los sistemas de polinización de agroecosistemas podría ayudar a comprender de una manera más precisa las relaciones de interdependencia existentes en sistemas productivos (Bohan et al., 2013).
Los niveles de especialización y complementariedad funcional pueden estar influenciados por el acople de los rasgos morfológicos de la red planta-polinizador (Maglianesi et al., 2014). El acople se refiere a la correspondencia de características fenotípicas de las especies de una comunidad (Blüthgen et al., 2008), el cual tiene un efecto ecológico en las funciones llevadas a cabo por las especies. Por ejemplo, los rasgos morfológicos tienen una incidencia directa en el uso y explotación de los recursos. En la red planta-polinizador, el acople de los rasgos morfológicos podría representar una alta calidad en el servicio de polinización de comunidades vegetales y una alta eficiencia en el uso de los recursos de los polinizadores (Dohzono et al., 2011).
Desde el punto de vista de las plantas, el tamaño de la flor afecta las relaciones con los polinizadores (Thompson, 2001) y condiciona el acople entre tamaño de la flor y tamaño del polinizador, lo cual podría tener un efecto positivo en la polinización. Igualmente, la abundancia floral es otro atributo ecológico importante que determina la cantidad de recursos disponibles para forrajeo de los polinizadores en una comunidad y, generalmente, hay una relación positiva entre abundancia de flores y de polinizadores (Potts et al., 2003). Desde la perspectiva de los polinizadores, los rasgos más importantes son la masa corporal y el largo de la lengua. La masa corporal de los polinizadores juega un papel clave en las redes de interacción ya que influye en la frecuencia, duración de las visitas y en la capacidad de contacto con los órganos reproductivos de las flores (Stang et al., 2009). Por otro lado, el largo de la lengua determina aspectos ecológicos importantes como la selección de las flores, la eficiencia en la adquisición de los recursos florales y la efectividad en la polinización (Cariveau et al., 2016). Sin embargo, hace falta evaluar el efecto de los rasgos morfológicos de plantas y polinizadores en sistemas de cultivo, ya que estos podrían influenciar los patrones de especialización y complementariedad que vendrían a determinar la eficiencia de la polinización.
En este estudio se cuantificó la red de polinizadores asociados a un agroecosistema y se evaluó el papel de la abundancia de las flores y de los principales rasgos morfológicos tanto de flores como de polinizadores sobre la duración de las visitas en un agroecosistema. El tiempo de contacto del polinizador en las flores tiene incidencia en la cantidad de polen que pueden recoger y transferir de una flor a otra (Wilson y Thompson, 1991). Por ejemplo, Ivey et al., (2003) encontraron una relación positiva entre la duración de la visita y la cantidad de polen transferido, así como con la eficiencia de la polinización. Por lo tanto, la duración de las visitas podría contribuir a entender la relación subyacente entre los polinizadores y las partes internas de la flor, en términos del potencial de los himenópteros como polinizadores. Así, los objetivos de esta investigación son 1) describir las interacciones entre las plantas de un agroecosistema y sus himenópteros polinizadores; 2) evaluar la importancia de los rasgos morfológicos en la polinización; y 3) determinar la relevancia de los himenópteros en la polinización de algunas plantas de consumo humano dentro de un agroecosistema en la región tropical.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se realizó en un agroecosistema en el corregimiento Los Andes del departamento del Valle del Cauca, Colombia (03°24' N y 76°33' W). El agroecosistema está ubicado en una finca de 6,3 ha donde se han implementado 0,75 ha de cultivo con esquemas agroecológicos. El corregimiento Los Andes se encuentra ubicado en la cordillera Occidental a 1031 m.s.n.m, al oriente del casco urbano de la ciudad de Cali y limita al occidente con el Parque Nacional Natural Farallones. El área de estudio corresponde a bosque seco tropical (Holdrige, 1976) y la temperatura media anual es de 23 °C.
Plantas
Se estableció una parcela de 10 m x 30 m donde se seleccionó un grupo de ocho especies de plantas del agroecosistema, todas dependientes de himenópteros para su polinización y en etapa de floración durante el tiempo de muestreo (Fig. 1). Las especies de plantas se dividieron en dos grupos: plantas de cultivo yplantas arvenses. Se seleccionaron entre dos y cuatro individuos para las especies de cultivo y entre cuatro y cinco individuos para las arvenses (para un total de 28 individuos). Las observaciones de los visitantes florales en dichos individuos se realizaron entre enero y febrero del 2017 durante 24 días, entre las 8:00 am y 12:00 m (para un total de 96 h de observación). Las observaciones se distribuyeron de la siguiente manera: dos horas de observación consecutivas para arvenses y dos horas para las plantas de cultivo, alternando la hora de inicio para cada grupo de plantas entre los días muestreados. Las observaciones se realizaron a 0,5 m de distancia de las plantas, siguiendo la actividad de los polinizadores y en días sin lluvia.

Figura 1 Especies de plantas involucradas en la red de polinizadores en el agroecosistema de Los Andes, Valle del Cauca, Colombia. De izquierda a derecha los nombres comunes y taxonómicos de las especies de planta, C: Correo (Cosmos caudatus); Aj: Ajenjo macho (Artemisia vulgaris); B: Botoncillo (Acmella mutisii); Am: Amaranto (Amaranthus spinosus); M: Mango (Mangifera indica); Bd: Badea (Passiflora quadrangularis); Al: Albahaca (Ocimum basilicum); Z: Zapallo (Cucurbita maxima).
Plantas y rasgos morfológicos.
Los rasgos morfológicos de las flores más relevantes para los insectos en la adquisición de los recursos nutricionales son el largo y ancho de corola (Hoehn et al., 2008), así como la abundancia floral (Potts et al., 2003). En campo se colectaron flores de otros individuos fuera de la parcela de observación, a las cuales se les midió el largo y ancho de la corola con un pie de rey. Se tomaron flores de tres individuos por especie de planta (tres flores por especie) y se obtuvo un promedio de cada rasgo por especie. Se contó el número de unidades florales de cada especie de planta antes de cada jornada de muestreo, para obtener un promedio de la abundancia floral disponible. La abundancia fue estimada como el número máximo de flores disponibles por especie de planta.
Red de polinizadores.
A partir de las frecuencias de visita registradas para cada especie de polinizador se elaboró una matriz de interacción entre todas las especies de plantas y polinizadores registradas en la parcela. Con esta matriz se construyó la red de interacción y se obtuvieron los índices cuantitativos de la estructura de la red. Tanto para el cálculo de las métricas de red, como para la visualización de la red de polinizadores, se usó el paquete informático 'Bipartite' del Software R (Dorman et al., 2008; R Core Team, 2016).
Polinizadores
Para cuantificarlas visitas de los polinizadores se realizaron muestreos diarios de cuatro horas de observación focal. Se asumió que los visitantes florales fueron polinizadores si estos tuvieron contacto con las partes reproductivas de la flor. Para cada polinizador se midió la duración de la visita en cada planta utilizando un cronómetro. Posteriormente, se obtuvo un promedio de duración de visita para cada especie de polinizador en cada especie de planta. La identificación en campo de las especies de polinizadores se realizó a nivel de morfoespecie con base en características morfológicas diferenciales y evidentes a simple vista como tamaño y coloración. Después de las observaciones focales se realizaron capturas de himenópteros en áreas aledañas a la parcela de observación, para la identificación en el laboratorio. La identificación se realizó usando claves taxonómicas (Smith-Pardo y Vélez-Ruiz, 2008) y ejemplares de referencia del Museo de Entomología de la Universidad del Valle (MUSENUV). El 59 % de los especímenes fueron identificados hasta género y el 35 % hasta especie. Sólo una morfoespecie fue identificada hasta familia (Megachilidae) y solamente una morfoespecie no se pudo colectar en campo y no se pudo identificar, por lo que fue excluida de los análisis estadísticos. Todas las observaciones se realizaron en días soleados y sin lluvia.
Polinizadores y rasgos morfológicos.
Para evaluar el efecto de los rasgos morfológicos en la duración de la visita floral se estimó el largo de la lengua y la masa corporal de las abejas, usando la distancia entre los dos puntos de inserción de las alas (distancia inter-tegular, DI). Se obtuvo un promedio por especie de la DI a partir de la medición de dos ejemplares por especie. Se usó la DI y la información de familia taxonómica para predecir el largo de la lengua y la masa corporal utilizando el paquete BeeIT de R, desarrollado por Cariveau et al., (2016). Este paquete está diseñado sólo para las ocho familias de abejas pertenecientes a la superfamilia Apoidea, por lo tanto, la longitud de lengua no fue estimada para las avispas. Sin embargo, se obtuvo un promedio de masa corporal de las especies de avispas pesando dos individuos por especie con una balanza analítica (0,1 mg ± 1).
Análisis de datos
Métricas de la red.
La estructura de la red se evaluó a partir de métricas cuantitativas. A nivel de red se obtuvo el grado de especialización complementaria mediante el índice H2', el cual mide la segregación de las especies, donde valores cercanos a uno sugieren una partición de nicho elevada y una alta especialización a nivel de comunidad; los valores cercanos a cero sugieren que el nivel de especialización es bajo (Blüthgen et al., 2006; Fründ et al., 2010). También se cuantificó el grado de anidamiento de la red, representado con el índice NODF. Este índice se interpreta como la asimetría en la especialización de la red, es decir, la relación de los especialistas (especies con pocos enlaces) con los generalistas (especies con muchos enlaces) en la red. Los valores de NODF oscilan entre cero y 100, valores altos se traducen en alto anidamiento y valores bajos en poco anidamiento (Almeida-Neto et al., 2008). Adicionalmente, se obtuvo el índice de conectancia (C) y el de robustez (R). La conectancia es la proporción realizada entre los enlaces de las interacciones entre plantas y polinizadores de la red. Los valores cercanos a uno se traducen en conectancia perfecta (Dunne et al., 2002). El grado de robustez de la red (R) se mide como una capacidad de resiliencia frente a eventos de extinción de especies (Bascompte y Jordano, 2007). Finalmente, se estimó la superposición de nicho (SN), métrica que corresponde a la similitud en los patrones de interacción (i.e. uso de un mismo recurso) entre especies de un mismo grupo. Para este índice, los valores cercanos a cero indican una partición de nicho elevada y, valores cercanos a uno, una superposición de nicho total (Blüthgen, 2010).
A nivel de especies se calcularon las métricas más relevantes para describir el rol ecológico de cada especie dentro la red. El índice d' mide el nivel de especialización y describe la fuerza del uso de recursos en comparación con la disponibilidad de este. Los valores de d' cercanos a cero corresponden a un comportamiento generalista, y los cercanos a uno, especialista (Blüthgen et al., 2006; Fründ et al., 2013). También se obtuvo el grado de distribución de las especies (i.e. degree) en la red, el cual corresponde al promedio del número de enlaces por especie (Bascompte y Jordano, 2007). Para los polinizadores se midió el índice de servicio de polinización (PSI), el cual mide la importancia de cada himenóptero como polinizadorpara la comunidad de plantas; los valores cercanos a cero indican irrelevancia de la especie como polinizador, y valores cercanos a uno corresponden a la importancia máxima como polinizador (Dormann, 2011). Todas las métricas fueron estimadas con el paquete 'Bipartite' del Software R (Dormann et al., 2008).
Análisis estadístico.
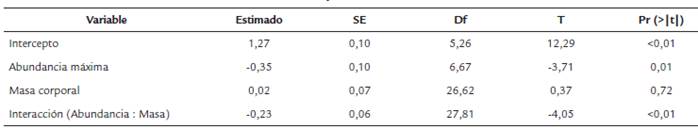
El efecto de los rasgos morfológicos de plantas y polinizadores sobre la duración de la visita se evaluó mediante un modelo lineal mixto (LMM). Previamente se evaluó la correlación entre las variables predictoras para excluir aquellas variables altamente correlacionadas (r > 0,7). El ancho de la corola fue excluido de los análisis debido a su alta correlación con el largo de la corola (r = 0,95). De igual forma, el largo de la lengua fue excluido por su alta correlación con la masa corporal de los himenópteros (r = 0,91). Así, las variables predictoras que finalmente se incluyeron en el modelo fueron la abundancia de las flores, el largo de la corola y la masa corporal de los polinizadores. Como variable de respuesta se usó la duración de las visitas (i.e. promedio de la duración de las visitas individuales por especie de polinizador en cada especie de planta). Todas las variables se transformaron logarítmicamente (base 10), se escalaron y se centraron para ser incluidas en el modelo. Para identificar el mejor modelo, se construyó un modelo global incluyendo todas las variables predictoras como factores fijos (i.e. abundancia floral, largo de la corola, masa corporal de los polinizadores y la interacción entre la abundancia y masa). La identidad de la especie de planta fue incluida como factor aleatorio en el modelo, lo cual permite tener en cuenta el efecto de las múltiples observaciones registradas para cada especie de planta. Se compararon todas las combinaciones de modelos posibles dentro del modelo global usando el criterio de información de Akaike (AICc), mediante el uso de la función de 'dredge' (paquete 'MuMIn' en R, Barton, 2016). Se consideraron sólo los modelos con un (AICc < 2 y que fueron significativos (Burnham y Anderson, 2002). Al final, solo se obtuvo un único modelo como el mejor modelo y no hubo necesidad de obtener promedios ponderados de los estimados de múltiples modelos. Todos los análisis se hicieron con el paquete 'MuMIn' en el Software R (Barton, 2016; R ver. 3.3.3, R Core Team 2016).
RESULTADOS
Las plantas de cultivo seleccionadas pertenecen a cinco familias taxonómicas, mientras que todas las plantas arvenses pertenecieron a la familia Asteraceae. En relación con los polinizadores, se identificaron en total 17 especies de cinco familias del orden Hymenoptera (Fig. 2), las cuales visitaron las plantas presentes en el agroecosistema. Todas las especies de polinizadores son nativas de Colombia, excepto la abeja doméstica Apis mellifera (Nates-Parra, 2005). La familia de himenópteros con mayor cantidad de especies registradas fue Apidae con 11 especies, de las cuales, ocho pertenecen a la tribu Meliponini. Esta tribu es una de las más representativas de la fauna himenóptera nativa de Colombia (Nates-Parra y Rosso-Londoño, 2013). Las avispas estuvieron representadas únicamente por dos especies, Pepsis sp. y Mischocyttarus sp.
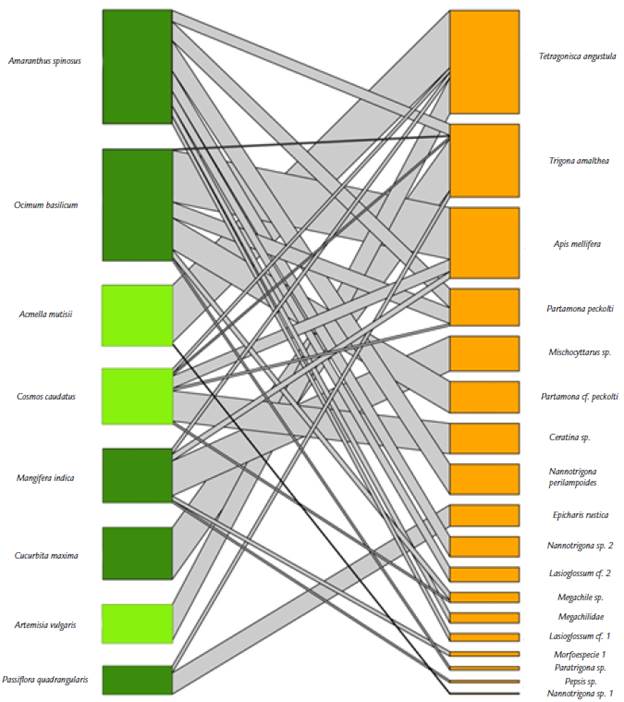
Figura 2 Red de polinizadores del agroecosistema de Los Andes, Valle del Cauca, Colombia. La red incluye ocho especies de plantas (cinco plantas de cultivo y tres especies arvenses) y 17 de polinizadores pertenecientes a cinco familias del orden Hymenoptera. El color verde claro representa las plantas arvenses, el verde oscuro las plantas de cultivo y el color naranja representa los polinizadores; las bandas grises son el número de enlaces entre plantas y animales. El grosor de la banda representa la fuerza o importancia de la interacción entre el par de especies.
Red de polinizadores.
La red de polinizadores estuvo conformada por 25 especies entre plantas e himenópteros (Fig. 2). Las plantas tuvieron un promedio de enlaces por especie de 4,6, mientras que los polinizadores de 2,6. Se encontró que la red es especializada, con un H2' = 0,8 y con un grado de anidamiento notablemente bajo (NODF= 10,4). Igualmente, la conectancia a nivel de red fue baja (C = 0,2).
Se encontró que los polinizadores serían más robustos (R= 0,7) a la pérdida de plantas, y que las plantas son levemente más sensibles a la pérdida de polinizadores (R= 0,6). La superposición de nicho para las plantas fue notablemente baja, es decir, que hay una alta partición de los nichos de las plantas (SN = 0,10). Igualmente, para los polinizadores la superposición de nicho fue baja (SN = 0,18), lo que muestra complementariedad funcional y poca redundancia en las interacciones.
A nivel de especies se encontró que las plantas fueron altamente especializadas y dependientes de unos pocos polinizadores (Tabla 1), siendo el zapallo (Cucurbita maxima) la especie con el índice más alto de especialización (d' = 0,85), seguida por la badea (Passiflora quadrangularis) (d' = 0,80).
Tabla 1 Métricas de la red de interacción a nivel de especie para las plantas y polinizadores en el agroecosistema de Los Andes, Valle del Cauca, Colombia. Las métricas representan el valor del número de interacciones por especie (degree), el índice de especialización (d') y la importancia en el servicio de polinización (PSI) para los himenópteros.
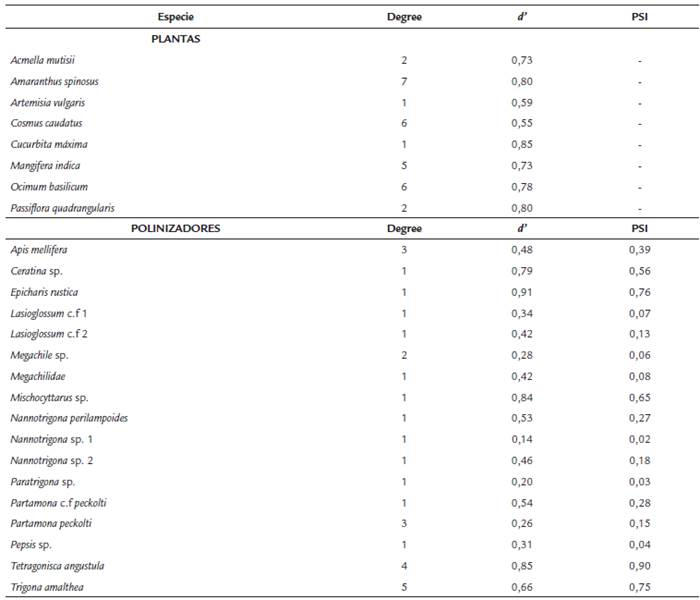
El zapallo sólo fue visitado por una especie de polinizador (Trigona amalthea) y la badea por dos especies (T. amalthea y Epicharis rustica). Se encontró que las plantas del agroecosistema son más especialistas que los polinizadores, ya que el 75 % de las especies tienen un d' > 0,7.
Con relación a los polinizadores, se encontró que cuatro especies de polinizadores son especialistas (d' > 0,7) y 13 son generalistas (Tabla 1). Los polinizadores más especializados de la red fueron E. rustica (d' = 0,91), Tetragonisca angustula (d' = 0,85), Mischocyttarus sp. (d' = 0,84) y Ceratina sp. (d'= 0,79). Por el contrario, A. mellifera tuvo un comportamiento generalista (d' = 0,48). Con respecto a los valores del índice de servicio de polinización (PSI), se encontró que T. amalthea, E. rustica y T. angustula son las especies de himenópteros más importantes en el agroecosistema debido a su servicio como polinizadores, ya que fueron las especies con los valores más altos de PSI (i.e. 0,75; 0,76; 0,90; respectivamente). Las 13 especies restantes no mostraron un servicio de polinización considerable en la comunidad de plantas agroecológicas (Tabla 1).
Rasgos morfológicos.
El análisis de los rasgos morfológicos reveló que la duración de las visitas de los polinizadores está influenciada por la masa de los polinizadores, la abundancia floral y la interacción entre masa y abundancia (Fig. 3). Se encontró que la abundancia de las flores tuvo un efecto negativo en la duración de la visita y la masa corporal de los polinizadores no tuvo efecto significativo como variable individual. Sin embargo, la combinación de abundancia y masa corporal tuvo un efecto negativo significativo en la duración de la visita (Tabla 2).
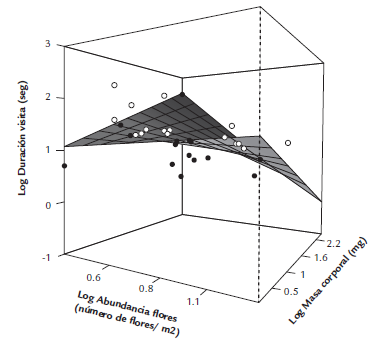
Figura 3 Relación entre la abundancia de las flores, la masa corporal de los polinizadores y la duración de la visita de los himenópteros de la red de polinizadores del agroecosistema de Los Andes, Valle del Cauca, Colombia. Los valores representados en los ejes corresponden al logaritmo base 10 de los valores reales. Los valores mayores a la predicción del modelo se muestran en color blanco y los valores menores en color negro.
DISCUSIÓN
Los resultados de nuestro estudio muestran que las interacciones entre plantas de uso agrícola y sus polinizadores son altamente especializadas, tanto a nivel de red como a nivel de especie. A nivel de red se encontró un grado de especialización particularmente alto (H2' = 0,8). Se encontró además un nivel de anidamiento bajo, y por ende, una baja redundancia en las interacciones. Esto indica que tanto plantas como polinizadores tienen una partición de nicho elevada, es decir que las especies se estructuran en grupos funcionales complementarios. Sin embargo, aunque se ha propuesto que la diversidad de especies y la riqueza de nichos funcionales reduce la competencia interespecífica (Blüthgen y Klein, 2011) y mejoran la provisión del servicio de polinización (Fründ et al., 2013), la alta especialización y complementariedad encontrada en este agroecosistema sugiere vulnerabilidad ante posibles perturbaciones como la desaparición parcial o total de alguna de las especies interactuantes en el agroecosistema (Bascompte y Jordano, 2007).
A nivel de especie nuestros resultados sugieren que las plantas son más especialistas y menos resilientes que los polinizadores, lo que indica que las plantas son más dependientes a polinizadores específicos, particularmente a especies nativas. Como resultado, habría un menor grado de resiliencia en caso de pérdida de alguna especie de polinizador en el agroecosistema. En contraste, los polinizadores no mostraron altos grados de especialización, lo que indica una mayor flexibilidad de los himenópteros a cambios en la oferta de flores gracias a su capacidad de vuelo que les permite encontrar recursos según su disponibilidad (Blüthgen y Klein, 2011). Nuestros resultados indican que los valores de PSI para tres especies de polinizadores fueron altos, lo cual sugiere que esas especies son altamente requeridas dentro de la comunidad agroecológica por su contribución a la polinización. Si bien A. mellifera es ampliamente utilizada en programas de polinización dirigida, se ha demostrado en varios estudios que la presencia de especies de himenópteros nativos es indispensable para asegurar la polinización de algunos cultivos neotropicales, por ejemplo de tomate, aguacate y cucurbitáceas (e.g. Quezada-Euán, 2009; Garibaldi et al., 2011; 2013). En nuestro sistema de estudio, A. mellifera no obtuvo un índice de servicio de polinización alto para ninguna especie de planta. En contraste, las especies nativas obtuvieron un índice de servicio de polinización alto, por ejemplo, T. amalthea (PSI = 0,75; d' = 0,66) con relación al zapallo, E. rustica (PSI = 0,76; d' = 0,91) en relación a la badea y la avispa Mischocyttarus sp. (PSI = 0,65; d' = 0,84) quien fue importante en la polinización del mango. En consecuencia, nuestro estudio resalta la importancia de las especies de abejas y avispas nativas en la polinización de alimentos comunes en la dieta de los colombianos.
Se ha sugerido que los mecanismos responsables de los grados de especialización y complementariedad funcional en las redes de polinización son los rasgos morfológicos y ecológicos de las especies interactuantes (Maglianesi et al., 2014). Nuestros resultados destacan la relación existente entre la masa corporal de los insectos y la abundancia de las flores, como dos factores que influencian la duración de las visitas en una comunidad de plantas agroecológicas. El comportamiento de forrajeo de los polinizadores está fuertemente vinculado a la disponibilidad de los recursos florales y el número de flores visitadas depende de las distancias entre flores y/o plantas (Ghazoul, 2005; Gutiérrez-Chacón et al., 2018). Nuestros resultados muestran que hay una relación negativa entre abundancia floral y duración de visita, lo que indica que la duración de las visitas de los himenópteros de menor tamaño aumenta cuando la abundancia de flores disminuye. Es decir, si la abundancia de flores en una comunidad es baja, los polinizadores se verían forzados a permanecer más tiempo en las flores para obtener la mayor cantidad de recursos nutricionales posibles, antes de movilizarse a otros recursos (Ortiz-Sánchez y Tinaut, 1994). Este comportamiento podría facilitar la polinización de especies particulares, debido a que las visitas largas podrían incrementar el contacto entre el polen recolectado y las partes internas de la flor, así, la deposición del polen podría ser más efectiva para la polinización. Por el contrario, si hay una mayor abundancia de flores, las visitas de los polinizadores de gran masa corporal tienden a ser cortas (Ortiz-Sánchez y Tinaut, 1994), lo que podría incidir en un mayor movimiento de polen entre las flores de una especie. Nuestros resultados ilustran que la masa corporal únicamente tiene un efecto ligado a la abundancia de las flores (i.e. la interacción), es decir que cuando los polinizadores son grandes y el recurso floral es abundante, las visitas tienden a ser cortas y viceversa. Esto sugiere que los himenópteros de gran masa corporal podrían ser más eficientes que los himenópteros pequeños en el forrajeo, lo cual podría deberse a que los himenópteros grandes requieren mayores cantidades de proteínas y azúcares, por lo que resulta más eficiente buscar dichos componentes cuando la abundancia floral es alta (Greenleaf et al., 2007; Blüthgen y Klein, 2011). En nuestro estudio, especies de gran tamaño como E. rustica y Pepsis sp. realizaron visitas prolongadas en plantas con poca abundancia como la badea y el mango, respectivamente, lo que sugiere que hay un mayor aprovechamiento de los recursos ofrecidos por estas especies, en vista de la poca abundancia de flores. Nuestros resultados se corresponden con estudios previos que examinan el comportamiento de los polinizadores dentro de las flores para determinar su efectividad en la polinización. Ivey et al., (2003) encontraron que, el taxón de insectos más efectivo en la transferencia de polen consume más tiempo forrajeando en cada flor. Esto sugiere que las plantas atractivas para polinizadores que invierten mucho tiempo en las flores suelen tener un éxito de polinización mayor. De esta forma, los requerimientos nutricionales de los polinizadores, el tamaño corporal, la abundancia de las flores y el comportamiento de forrajeo de los insectos podrían determinar la intensidad en la explotación de los recursos disponibles en una comunidad o agroecosistema.
La alta dependencia de las plantas agroecológicas a polinizadores específicos encontrada en este estudio sugiere una mayor vulnerabilidad de las plantas frente a eventos de pérdida de polinizadores. Esto se relaciona con la potencial crisis que enfrentan los sistemas agrícolas a nivel mundial debido a la tendencia actual de disminución de poblaciones de polinizadores naturales (Kremen et al., 2002; Martin, 2015). Una alternativa para atraer polinizadores es tener diferentes tipos de flores. Por ejemplo, en el caso de la agroecología, uno de sus principios es la promoción de plantas silvestres y/o arvenses dentro de los agroecosistemas, pues éstas contribuyen al mantenimiento y atracción de la diversidad de polinizadores naturales (Nicholls y Altieri, 2013). Nuestros resultados muestran que las plantas arvenses fueron atractivas para varios polinizadores de las plantas de cultivo. Por ejemplo, la especie arvense Cosmos caudatus fue visitada por seis especies de polinizadores, donde la mayoría de ellos visitaron todas las especies de cultivo del agroecosistema. Por lo tanto, nuestro estudio señala la importancia de implementar sistemas agrícolas heterogéneos y diversos (i.e. diversidad de especies cultivadas y plantas arvenses) para atraer y maximizar la diversidad de polinizadores que visitan los agroecosistemas, y así minimizar la vulnerabilidad en la polinización. De esta manera, se podrían incorporar prácticas para ayudar al mantenimiento de la diversidad de polinizadores en los sistemas de producción. Así, los agroecosistemas tienen la capacidad de potencializar, desde una perspectiva ecológica, la relación entre diversidad y función.
La cuantificación de la especialización y la estructura de las redes ecológicas podrían estar afectadas por sesgos del muestreo. Sin embargo, estudios previos como el realizado por Fründ et al., (2016) demuestran que el sesgo asociado al muestreo es menor en redes especializadas que en redes generalistas. En las redes especializadas, se ha identificado que las métricas con menos riesgo a sesgo son d', H2' y PSI, métricas que se usaron en este estudio. De igual forma, se ha establecido que una posible regla para determinar el número de observaciones requerido para obtener una red cuantitativa y con métricas confiables se logra a partir de un rango de 15-24 observaciones por especie de consumidor (Fründ et al., 2016). En nuestro estudio, el número de observaciones realizadas a 17 especies de polinizadores se ubica dentro de ese rango sugerido. Solo dos especies, Pepsis sp. y Nannotrigona sp. (con dos y una observación, respectivamente) quedaron por debajo del rango; éstas fueron especies raras dentro de la red. Así, aunque el muestreo fue corto, las métricas estimadas generan resultados sólidos. No obstante, cabe la posibilidad que un muestreo más intensivo y a largo plazo, que incluya tanto polinizadores diurnos como vespertinos, genere cambios en los patrones de especialización de la red de interacción, lo que podría originar mayores valores de anidamiento o modularidad (Fründ et al., 2016).
CONCLUSIONES
Nuestro estudio evidenció que la estructura de la red de himenópteros polinizadores revela patrones como complementariedad funcional y especialización en las interacciones entre plantas y polinizadores de un agroecosistema, y que estas interacciones son dependientes de los rasgos específicos de las especies interactuantes. Futuras investigaciones podrían ser encaminadas a abordar el espectro completo de las demás interacciones planta-animal que ocurren en los agroecosistemas (i.e. las relaciones de herbívora). Al conocer la red antagónica de las plantas se podrían ejercer mecanismos de control adecuados para maximizar el rendimiento de estos sistemas de cultivo y controlar las pérdidas por daño de herbivoría en los agroecosistemas. La inclusión y manejo de otras redes de interacción en los agroecosistemas fortalecería los fundamentos sobre los cuales se basa la agroecología, y le darían valor y sostenibilidad en el tiempo. La implementación del estudio de redes de interacción, particularmente las de polinización, en agroecosistemas podría ser una herramienta útil para monitorear y optimizar la producción de alimentos en los territorios rurales del país y América Latina.


