











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCTION
Changes in land use, associated with urbanization, and expansion of agricultural frontiers may promote a water quality reduction due to the increase in the nutrient load, especially of phosphorous and nitrogen (Serna et al., 2019). The Brazilian semiarid region has a great susceptibility to desertification process, due to historical climate conditions (Vieira et al., 2013), intensive land-use and human growing leading to fragilization of the terrestrial ecosystems and soil loss (e.g. Silva et al., 2018; Azare et al., 2020).
Eutrophication is one of the well-described impacts of the present time, almost always associated with the increase in cyanobacteria biomass and a drastic reduction of submerged macrophytes (Dondajewska et al., 2019), besides the simplification of the trophic structure, loss of biodiversity and species extinction (Dubey and Dutta, 2020). Therefore, an increase of nutrients is directly associated with the primary production increase and, consequently excessive growth of algae (Ballah et al., 2019). Some authors indicate that reservoirs in the Brazilian semiarid region are more prone to eutrophication (Barros et al., 2019). Currently, prolonged, and extreme drought has been accelerating this process (Andrade and Nunes, 2017, Costa et al., 2015).
In well-preserved watershed, with functional riparian forests, changes in factors such as the watercolour and concentrations of suspended solids and phosphorus, occur in a less magnitude (Li et al., 2019). The preservation degree of the riparian zone indicates the relevance of vegetation cover as a mechanical impediment for surface runoff, reducing soil loss and sediment transport to the reservoirs (Flores et al., 2019).
Seasonal changes in the water quality may be monitored through abiotic and biological variables accumulated in the surface sediments (Wang et al., 2019). The sediment layers are the result of the deposition of biological, physical, and chemical materials from the surrounding area of the reservoir and can be attributed to both natural and anthropic factors (Silva et al., 2019). Diatoms, present in the surface sediments of aquatic ecosystems, are originated in different habitats such as plankton, benthos, and periphyton (Tudesque et al., 2019; Riato and Leira, 2020). Due to characteristics, such as populations with large numbers of individuals, sensitivity or tolerance to short and long-term changes, easier sampling, and high rates of migration and colonization of new habitats (Marquez and Espino, 2019), diatoms have been used as bioindicators (Szulc and Szulc, 2013). In addition, the high abundance in several types of aquatic ecosystems (lotic environments, estuarine, marine, and hypersaline waters), high preservation of frustules in sediments (Goghe and Rynearson, 2017; Rebolledo et al., 2019) and are associated with variable pH conditions, from acidic to alkaline (Sienkiewick and Gasiorowski, 2017).
The objective of this study was to understand the characteristics of the diatomological community and the use and occupation of the soil around the three reservoirs (Arara, Rio do Canto and Araçagi) and in the natural lagoon (Lagoa do Paó) (Northeast Brazil) according to some limnological variables and types of land use and land cover.
MATERIALS AND METHODS
Study Area
The man-made waterbodies are in the Mamanguape River (Northeast region), the physiographically known as an open ombrophilous forest (altitude swamps), deciduous and semi-deciduous seasonal forests (AESA, 2006) (Fig. 1). Santos et al. (2015) identified three regions with different patterns of accumulated rainfall, western region (between 400 to 800 mm), central region (800 to 1400 mm), and eastern region (1000 to 1600 mm).
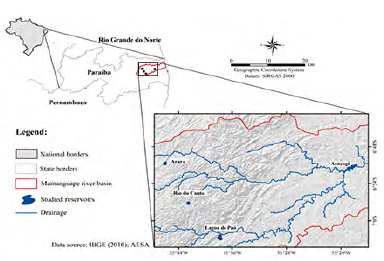
In this study, we selected four water bodies according to the trophic state and rainfall gradient: Arara (6° 50'41 85"S 35° 46'02 85" W), Rio do Canto (6° 57'07 56 "S 35° 42'35 65" W), Lagoa do Paó (7° 02'31 8 "S 35° 37'50 8" W) and Araçagi (6° 49 635'S 35° 21 413'W).
Land use and land cover mapping
We used spatial images with high-resolution (2012, 2014, 2015) captured from Google Earth and Bing Maps. The visual interpretation technique was estimated according to Novo (2008).
The influence area (buffer) was established according to the size of the water bodies. Therefore, after vectorization, all the polygons corresponding to the same class were merged to calculate the respective influence area.
Sampling and limnological analyses
Water and surface sediment samples from the limnetic zone were collected between September and November 2012. Temperature (oC) and pH were estimated "in situ" with a multiparameter probe. Transparency was estimated "in situ" with a Secchi disk (Cole, 1994). Euphotic zone (Zeu) was calculated as 2.7 times the Secchi depth (Cole, 1994) and the coefficient of light attenuation (k) was calculated according to Poole and Atkins (1929). Water total phosphorus ((µg.L-1) was measured (APHA, 1998). The trophic state index (TSI) was estimated through the Carlson (1977) modified for tropical environments by Toledo et al. (1983).
Sediment sampling and determination of total phosphorus and organic matter
The total phosphorus of the sediments was determined according to Andersen (1976). The organic matter content (g.kg-1) was analysed through the Walkley and Black (1934).
Diatom analysis
The diatoms composition and abundance were determined using the method described by Battarbee (1986) and Battarbee et al. (2001), by allowing approximately 3 g of wet mass dried. The oxidation was based on in an oven at 50 °C for 24 hours. The diatoms were oxidized with hydrochloric acids and, permanent slides were prepared with Naphrax® as an inclusion medium (IR = 1.73). For the identification of species, the specific literature was used (Patrick and Reimer, 1975; Round et al., 1990).
Abundant species were determined obtained through the 400-valve counting (Bicudo, 1990). The species abundance was calculated (Battarbee, 1986) and frequency of occurrence was obtained according to Matteucci and Colma (1982), considering the number of samples in which a given taxon occurred in relation to the total number of samples analysed.
Statistical analysis
Principal Component Analysis (PCA) was used to assess the patterns of distribution of diatoms in the reservoirs according to the limnological variables and, the land use in the watershed. For PCA analysis of all data series, the matrix with data transformed by log (x + 1) was used. Statistical analyses were performed with the R statistical package (R Core Team, 2017).
RESULTS
Land Use and Land Cover
Four types of land use were identified: urban or constructed area (URB), exposed soil (EXPS), arbustive/ arboreal vegetation (ARB) and anthropic landscape (ANTR) composed by agriculture and livestock. This land use was dominant in the watershed (> 50 %) (Fig. 2 and Table 1).
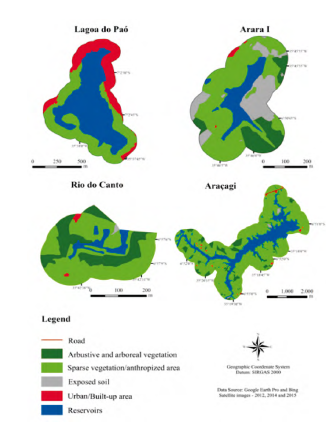
Despite the high proportion of the anthropic landscape surroundings of Arara (59.61 %) and Araçagi (57.20 %), these reservoirs had clear waters (Zeu ≥ 2.7 m), alkaline pH (≥ 8.5), low total phosphorous concentrations (≥ 21.3 (µg.L-1), and high levels of total phosphorus in the surface sediments (≥ 107.5 Mg.g-1) (Table 1). In the Rio do Canto, we also observed the dominance of the anthropogenic landscape (67.49 %), with perennial trees and extensive livestock. Besides it, approximately 30.28 % of the area was occupied by arbustive/arboreal vegetation, reforested areas, and remnants of riparian forest (Table 1). A high concentration of total phosphorus (53.3 Mg.L-1) and low transparency (≥ 0.8 m) was observed in the waters.
Table 1 Limnological variables and types of land use and occupation of the studied reservoirs.
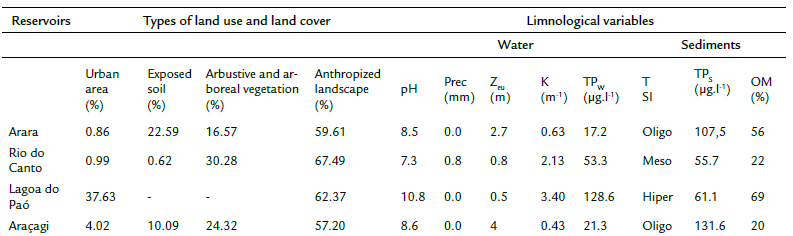
Abbreviations: pH (Hydrogen potential); Pre (Precipitation); ZEU (euphotic zone); K (coefficient of light attenuation); TPw (total phosphorus in water); TPs (total phosphorus in the sediments); TSI (trophic status index); Oligo (oligotrophic); Meso (mesotrophic); Hyper (hypereutrophic).
In Lagoa do Paó, we observed a high percentage of the urbanized area (37.63 %), with a total absence of vegetation cover. Consequently, Zeu was lower than other reservoirs (Zeu = 0.5), with the high phosphorus concentrations in the water (128.6 µg.L-1).
The PCA results explained 80.79 % the total variance of data in the first two axes (Axis 1: 43.33 % and Axis 2: 37.46 %) (Fig. 3). Axis 1 showed the sampling units associated with the urbanized area (r = 0.42), high phosphorus concentration in water (r = 0.41), and a high light attenuation coefficient (r = 0.29) and hypereutrophic conditions. In contrast, Axis 2 represented oligotrophic and mesotrophic reservoirs, associated with the anthropogenic landscape (r = - 0.50) and high phosphorus concentrations in the sediments (r = - 0.38).
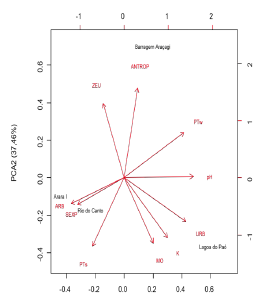
Figure 3 Principal Component Analysis (PCA) of the limnological variables, land use and land cover in the studied reservoirs. References: URB-Urban area/Built-up area, ARB- Arbustive and arboreous vegetation, EXPS - Exposed soil, K- coefficient of light attenuation, MO- organic matter, pH- Hydrogen potential, ZEU- Euphotic zone, ANTR- anthropized vegetation, PTw- Total phosphorous in water, PTs- Total phosphorous in the sediments.
Diatom assemblages
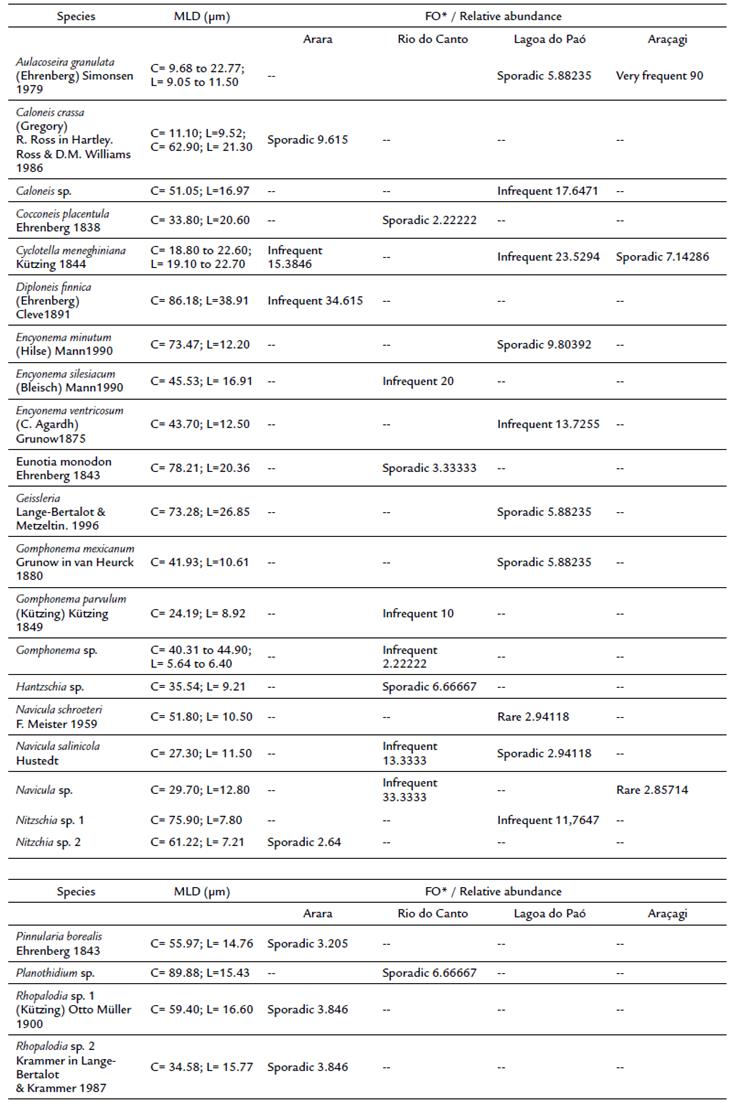
A total of 24 taxa were identified, distributed in the 12 genera and 10 families. Regarding the genera level, Aulacoseira, Caloneis, Cocconeis, Cyclotella, Diploneis, Encyonema, Eunotia, Geissleria, Gomphonema, Hantzchia, Navicula, Nitzchia, Pinnularia, Planothidium and Rhopalodia were the most representative and the frequency of occurrence ranged from sporadic to very frequent (Table 2). The part of the taxa corresponded to Pennales (19 %). Between the waterbodies, Rio do Canto presented the highest richness (11 taxa), a low abundance (627 valves/g), and four exclusive genera (Hantzschia, Cocconeis, Eunotia, Planothidium). In contrast, the Araçagi reservoir had the lowest richness (3 taxa) and the highest abundance (4182 46 valves/g).
The PCA summarized 70.4 % of the total variability of data on the first two axes (Fig. 4). The positive side of Axis 1, Rio do Canto sampling units was associated with Navicula salinicola Hustedt (0.25), Eunotia sp. (0.25), Gomphonema parvulum (Kützing) Kützing (0.27) and Cocconeis placentula Ehrenberg (0.27). The sampling units of the Arara, Lagoa do Paó and Araçagi were ordered on the negative side of the axis 2 represented to the Nitzchia sp. (- 0.10), Diploneis finnica (Ehrenberg) Cleve (-0.12), Geissleria sp. (- 0.10), Cyclotella meneghiniana Kützing (- 0.23) and Aulacoseira granulata (Ehrenberg) Simonsen (- 0.03).
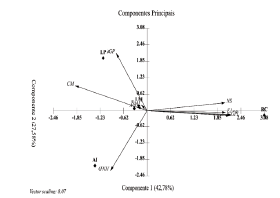
Figure 4 Principal Component Analysis (PCA) of the diatoms from the surface sediments of the reservoirs during the study period.
DISCUSSION
Land Use
In the arid and semi-arid regions worldwide, vulnerability related to the uneven distribution of rainfall and the high evapotranspiration rates are the main barriers for regional development (Andrade and Nunes, 2017) and maintenance of the water bodies water level. The expansion of economic activities may impact the landscape management and land use, which potential reduction of the resilience of the aquatic ecosystem. Such vulnerability favors an increase in primary production and, consequently, the eutrophication, almost always accompanied by changes in the composition and abundance of some groups of primary producers, such as cyanobacteria (Azare et al., 2020).
The PCA based on the limnological variables and land-use, showed two groups associated with the urbanization degree and its influence on the trophic state. Lagoa do Paó, classified as hypereutrophic, had the highest urbanized area, and the total absence of arbustive / arboreal cover. The lack of natural vegetation, the drainage of the untreated domestic sewage associated with the absence of aquatic macrophytes, might have caused an increase in the water phosphorus concentrations and, consequently, permanent blooms of toxin-producing cyanobacteria (Felix et al., 2015). Regarding the Rio do Canto, the predominance of reforested areas and remaining of riparian forests might have favoured the nutrients inputs in this reservoir, influencing the high phosphorus concentrations in water and low transparency, to a lesser extent than in Lagoa do Paó.
In this regard, the maintenance of mesotrophic state might be associated with the high density of emergent aquatic macrophytes (Eichornia crassipes Mart. (Solms) and Pistia stratioites L.). Macrophytes may act as a filter for nutrients and suspended organic matter and act as a nutrient sink after the vegetation senescence (Tan et al., 2019).
Diatom assemblages
Teixeira et al. (2018) and Hansika and Yatigammana (2019) observed that the Aulacoseira, Encyonema, Geissleria, Gomphonema and Nitzchia genera are considered frequent and distributed in meso-eutrophic environments. Bartozek (2018) considered the genera Cyclotella, Navicula and Diploneis (Faustino et al., 2016) as oligotrophic. This observation corroborates the dominance found in the present study.
Alexandre et al. (2017) reported that anthropization and the sewage discharge increase the vulnerability of water bodies to enrichment with nutrients, indicating the eutrophication process. Data such as these corroborate those reported in this study.
The greatest richness of diatoms was found in the sediments of Lagoa do Paó and Rio do Canto. This may be related to the high speed and intensity with which allochthonous and autochthonous materials accumulate in the sediments of shallow ecosystems with an advanced trophic state. Eutrophic environments are more prone to sedimentation due to improper land use (Bartozek et al., 2019), bringing together planktonic and peripheral diatoms from recent years in the upper centimeters of sediments.
According to Costa (2017) and Silva-Lehmkuhl (2019), genera such as Eunotia and Planothidium, as well as Hantzschia and Cocconeis (Faustino et al., 2016) tend to develop in the pleasant mesotrophic trophic. Corroborating the results seen in the mesotrophic Rio do Canto reservoir.
Navicula was one of the most abundant genus in Araçagi, Rio do Canto and Lagoa do Paó. Species of this genus have a great tolerance to pollution (Medeiros et al., 2017) and their occurrence is determined by conditions of low salinity and slightly acidic to alkaline pH (Zalat et al., 2019). According to Round et al. (1990), Navicula is widespread, occurring in oligotrophic to mesotrophic conditions, which can be related to its high adaptive plasticity.
Cyclotella was also registered in three of the four aquatic ecosystems (Arara, Lagoa do Paó and Araçagi), although it was more abundant in Lagoa do Paó, probably due to the shallow, turbid (Hansika and Yatigammana, 2019) and nutrient-enriched waters (Medeiros et al., 2017; Cutrim et al., 2019). In addition, it iscommonly associated with changes in the water volume due to the water balance of the reservoir and/or climate regional condition (e.g. Costa et al., 2015).
In the Araçagi and Lagoa do Paó reservoirs, Aulacoseira showed the highest abundance, commonly found in nutrient-enriched and turbid habitats (Medeiros et al., 2017; Sharaf et al., 2019) as a Lagoa do Paó. In contrast, the species of this genus were already reported in oligotrophic waters (Batista and Fonseca, 2018; Bartozek et al., 2019), such as Araçagi reservoir.
CONCLUSIONS
In summary, our results showed low species richness and high similarity between water bodies with different trophic gradients. Occurrence of epiphytic diatoms, benthic and planktonic, with distribution and frequency adapted to tolerance to organic pollution. Additional studies are recommended to monitor the water quality of semi-arid aquatic ecosystems, to assess the impact caused by the release of domestic sewage and the increase in human pressures and climate warming.

