











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
En los vertebrados, el ligando Sonic Hedgehog (Shh), expresado desde las primeras etapas de la embrio-génesis, desempeña un papel esencial en la regulación espacial y temporal del desarrollo del sistema nervioso central 1. Durante la organogénesis, Shh actúa como morfógeno, mitógeno y guía molecular, siendo fundamental para la maduración de diversas regiones del cerebro 1-2. Por ejemplo, la señalización mediada por Shh promueve la migración de las neuronas granulares hacia las láminas de Purkinje a través de la glía de Bergmann, un proceso clave para la correcta organización del cerebelo 3-4. Por su parte, la embriogénesis y la carcinogénesis comparten mecanismos celulares básicos, como la proliferación, migración y diferenciación, que requieren de una regulación precisa y coordinada.
En este contexto, las alteraciones en las vías canónicas y no canónicas de Shh, así como en sus funciones fisiológicas, están implicadas en el desarrollo de tumores cerebelosos específicos, incluido el meduloblastoma 5. El meduloblastoma, clasificado como un tumor embrionario neuroepitelial maligno de grado IV 6, es la neoplasia intracraneal más frecuente en pediatría, representando aproximadamente el 15 % de los casos en esta población 7. El estudio de los mecanismos moleculares que regulan el desarrollo normal y su disfunción en patologías como el meduloblastoma podría proporcionar información valiosa para el diseño de nuevas estrategias terapéuticas.
El objetivo de esta revisión es describir el papel fundamental de la vía Sonic Hedgehog, tanto en el desarrollo embrionario como en la tumorogénesis del sistema nervioso, con un enfoque en el meduloblas-toma. Se proporciona una base comparativa que facilita la comprensión de estos procesos y su potencial utilidad clínica y terapéutica. La revisión narrativa se llevó a cabo mediante una búsqueda en bases de datos como Medline y SciELO, utilizando palabras clave como: "Sonic Hedgehog", "nervous system", "embryogenesis", "neurodevelopment", "tumorige-nesis", "medulloblastoma" y "molecular mechanisms". Se seleccionaron artículos en inglés y español publicados en los últimos 20 años, eligiendo aquellos que responden al objetivo planteado por los autores.
Shh en el desarrollo y la diferenciación del tubo neural en vertebrados
La diferenciación del tubo neural sigue un eje ventral-dorsal, controlado por gradientes de concentración molecular que regulan este proceso 1,8-9. En la región ventral, la proteína Shh desempeña un papel fundamental, mientras que, en la región dorsal, la señalización mediada por la proteína morfogénica ósea (BMP, según sus siglas en inglés), modulada por la proteína Noggin, es crítica para los procesos de plegamiento, dorsalización, alargamiento, cierre y formación del tubo neural 10. Estructuralmente, el tubo neural primitivo se identifica como un epitelio cilíndrico pseudoestratificado, simétrico y bilateral, compuesto por células neuroepiteliales elongadas 11. Estas células progenitoras exhiben una orientación apical hacia el lumen del canal central y sus superficies basales forman los bordes laterales del tubo 12.
Durante la proliferación celular, los núcleos de las células progenitoras experimentan un movimiento estereotípico intercinético que regula la exposición de estos a las señales proliferativas 13. Por su parte, la mitosis tiene lugar en la superficie apical del epitelio, mientras que la síntesis del ADN ocurre en las células ubicadas en la superficie basal 14. Este proceso está controlado por un gradiente de señalización Notch que se extiende de manera apical a basal, con mayor actividad en los núcleos del compartimento apical y una regulación descendente hacia la superficie basal 5. Conforme los progenitores neurales se diferencian en neuronas posmitóticas, pierden su anclaje a la superficie apical del epitelio y migran lateralmente para posicionarse basalmente respecto a los cuerpos celulares de los progenitores 2,15. Este proceso implica la delaminación de la membrana apical mediante una constricción dependiente de actina y miosina, así como el desmantelamiento del cilio primario, una estructura crítica para la transducción de señales de la vía Shh 10,16.
Las células ubicadas en la línea media ventral del tubo neural se diferencian para formar la placa del piso, una estructura epitelial esencial para el desarrollo neural 17. La interacción de la placa del piso con Sonic Hedgehog (Shh) y la notocorda resulta crucial para el posicionamiento y la especificación de precursores neuroectodérmicos en la médula espinal y el romboencéfalo. Estas señales inducen la diferenciación de progenitores neurales en interneuronas ventrales y motoneuronas 18. En contraste, las células precursoras localizadas lejos de la línea media ventral generan neuronas de relevo sensorial e interneuronas asociadas a regiones dorsales de la médula espinal y el encéfalo posterior, procesos regulados por señales de proteínas Wnt expresadas en la placa del techo, una banda estrecha de células neuroepiteliales 19. La especificación neuronal depende críticamente de la concentración de Shh, ya que la diferenciación de cada tipo neuronal in vitro está correlacionada con la cantidad mínima de Shh requerida, reflejando la distancia desde su fuente de síntesis in vivo20. Neuronas generadas en regiones ventrales del tubo neural necesitan concentraciones más altas de Shh para su inducción 21. La remoción temporal de ligandos Shh provenientes de la notocorda y la placa del piso ha demostrado su importancia en la formación de la lámina perinoto-cordal, la somatogénesis, y la diferenciación de los precursores del esclerotomo, dermatoma dorsolateral y miotoma, además de regular la expresión del factor de crecimiento endotelial vascular ( VEGF, según sus siglas en inglés) 22-23. Alteraciones en esta vía, como la eliminación de la notocorda, la inhibición de Shh o la pérdida de sus receptores, provocan la pérdida del eje dorsal-ventral del tubo neural y la ausencia de motoneuronas e interneuronas ventrales 24-26.
Las rutas moleculares de Shh: vías canónica y no canónica
El mecanismo molecular de Shh se organiza en rutas canónicas y no canónicas 27. Ambas dependen críticamente del cilio primario, que también actúa como un centro de integración para otras moléculas señalizadoras, incluidas Wnt, el factor de crecimiento derivado de plaquetas (PDGF, según sus siglas en inglés) y notch28. La vía canónica de Shh está compuesta por cuatro elementos clave que interactúan para regular la transcripción de genes objetivo: el receptor patched-1 (PTCH1), con 12 dominios transmembrana; Smoothened (SMO), un receptor acoplado a proteína G; el regulador negativo supresor de homólogo fusionado (SUFU, según sus siglas en inglés); y los factores de transcripción GLI (GLI2, GLI2 y GLI3)29. GLI2 actúa exclusivamente como activador transcripcional, mientras que GLI2 y GL/3 desempeñan roles duales, siendo reguladores positivos o negativos dependiendo de modificaciones postranscripcionales y postraduccionales. GLI2, comúnmente considerado un activador, se acumula en el cilio primario y regula la transcripción en respuesta a Shh de manera independiente de los niveles de GLI3R, que es predominantemente un represor 30.
La unión de Shh al cilio primario inactiva PTCH2 , liberando la inhibición sobre SMO y permitiendo su translocación a la membrana ciliar, un paso esencial para la activación de los factores GLI29-30. Una vez activados, los factores GLI se trasladan al núcleo, donde promueven la transcripción de genes blanco que amplifican la señalización de Shh. Estos genes incluyen GLI2 y PTCH2, que generan retroalimentación positiva; ciclina D2 y MYC, que estimulan la proliferación celular; BCL-2, que inhibe la apoptosis; ANG2/2, que favorece la angiogénesis; Snail, que facilita la transición epiteliomesenquimal; y NANOGy SOX2, que promueven la autorrenovación de células progenitoras 31-32 (figura 1).
Fuente: elaboración propia.
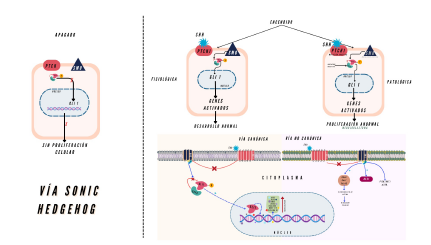
En ausencia del ligando Shh, SUFU regula negativamente la vía de señalización al unirse directamente a los factores de transcripción GLI, reteniéndolos en el citoplasma, facilitando su degradación y evitando la activación de genes blanco 33. SUFU también forma un complejo supresor que impide la interacción de GLl2 con el ADN, lo que inhibe la expresión génica mediada por GLl2 y convierte esta proteína en su forma reprimida (GLI-R). GLI-R, al translocarse al núcleo, bloquea la transcripción de genes clave como GLI2 y PTCH2 . La superación de esta inhibición requiere de la fosforilación de GLI mediada por diversas cinasas, incluidas PKA, PKC, GSK3ß y PIK334. Además, la activación de la histona deacetilasa 1 (HDAC2 ) desempeña un papel crítico en la modulación epigenética de la vía, potenciando la activación de GLI2 y GLI2A32. En presencia de niveles elevados de Shh en el cilio primario, la desacetilación de GLI2 y GLI2 promovida por HDAC2 incrementa significativamente la activación transcripcional de los genes regulados por esta vía 27,35.
En las vías no canónicas, la unión de Shh al receptor PTCH2 libera a SMO de su inhibición, lo que induce un cambio conformacional que activa diversas rutas no transcripcionales, sin involucrar directamente la transcripción mediada por GLI27,36. En este proceso, SMO interactúa con proteínas del citoesqueleto y otras moléculas señalizadoras, como proteínas G heterotriméricas y efectores aguas abajo ( RhoA, Rac2 y Cdc42), que regulan la dinámica del citoesqueleto de actina, crucial para la migración celular 37-39. Así mismo, se ha descrito que la señalización no canónica de Shh puede regular HES2 , un efector de la vía Notch y también puede interactuar con cascadas como PI3K/AKT, mTOR o MAPK, modulando procesos como la supervivencia, proliferación o migración, dependiendo del contexto celular 40-43.
La proteína interactuante con Hedgehog 1, HH/P2, una proteína secretada, actúa como antagonista de la señalización de Shh en colaboración con PTCH2, restringiendo su propagación a lo largo del tejido neural y manteniendo concentraciones más bajas en las regiones dorsales y distales del tubo neural 44 . En contraste, los correceptores transmembrana Cdon, Boc y Gasl facilitan la transducción de Shh en la región posterior del tubo neural 44. Durante etapas tempranas del neurodesarrollo, estos correceptores refuerzan la señalización de Shh en condiciones de baja concentración de esta proteína, sin embargo, con el aumento de los niveles de Shh, la activación de la vía conduce a una regulación negativa de los correceptores, modulando la intensidad de la señalización y previniendo una activación excesiva. Este mecanismo permite controlar las fluctuaciones de Shh y asegura una distribución continua que garantiza la ventralización adecuada del tubo neural 45 . Además de su papel en el tubo neural, estos correceptores están implicados en procesos críticos del desarrollo, como la morfogénesis cerebelosa, craneofacial y la especificación digital 1,8.
Shh y la evidencia vinculante a tumores
La importancia de las alteraciones en la señalización de Shh en el cáncer se evidenció inicialmente en estudios del síndrome de Gorlin, un trastorno autosómico dominante que se caracteriza por anormalidades craneofaciales y esqueléticas, además de un riesgo aumentado de carcinoma basocelular avanzado y meduloblastoma 46. En pacientes con este síndrome y carcinomas basocelulares y meduloblastomas concomitantes, las mutaciones en PTCH2 se identificaron como el principal factor genético asociado, con una frecuencia del 10 %, seguidas de mutaciones en SUFU (5-10 %) y SMO (5 %) 47.
En varios tipos de cáncer, las activaciones canónicas y no canónicas de la señalización Shh coexisten y, en algunos subtipos, los mecanismos no canónicos pueden predominar 48. Aunque la vía Shh es esencial durante el desarrollo embrionario, la homeostasis tisular se "apaga" después del nacimiento y solo permanece activa en ciertos subtipos de células troncales involucradas en procesos de reparación y regeneración tisular 49. Su reactivación en la infancia o la adultez puede dar lugar a procesos neoplásicos, y se ha observado una activación persistente de la vía Shh en varios tumores sólidos y hematológicos, asociándose con el desarrollo, progresión y recurrencia posquimioterapia 49-50. Estas vías pueden participar en la oncogénesis a través de tres mecanismos:
Activación independiente del ligando debido a mutaciones inactivas de PTCH2 o SUFU, mutaciones activas de SMO o expresión de genes amplificadores de GLI, como en el CBC y Shh-Mb.
Activación dependiente del ligando por vía autocrina o yuxtacrina, donde las células tumorales aumentan la expresión del ligando.
Activación dependiente del ligando por mecanismos paracrinos, en los cuales los ligandos son secretados por las células tumorales al estroma circundante, estimulando el crecimiento, la angiogénesis, la proliferación y la supervivencia tumoral.
En el año 2021, la Organización Mundial de la Salud publicó la clasificación de tumores del sistema nervioso central, subdividiendo el meduloblastoma en cuatro grupos según la vía de señalización implicada: activado por Wnt, activado por Shh, de grupo 3 y de grupo 4 6,51-52. El subtipo activado por Shh se origina en los núcleos cocleares y en células precursoras de neuronas granulares ATOH2 positivas, que proliferan en respuesta al ligando secretado por las células de Purkinje 3-4. Este subtipo, que representa entre el 25 % y el 30 % de los casos, se clasifica adicionalmente en TP53 mutante o silvestre 47,52. Aunque la mayoría de los meduloblastomas son esporádicos, aproximadamente el 5 % de los pacientes presentan síndromes hereditarios raros, como el síndrome de Gorlin y el síndrome de Li-Fraumeni. En este último, la mutación en TP53 destaca la importancia de su identificación, ya que la radioterapia, aunque esencial para el tratamiento, podría agravar el crecimiento de la lesión y aumentar el riesgo de neoplasias secundarias 6,53.
La activación de la vía Shh en este subtipo suele ser consecuencia de mutaciones en componentes clave como PTCH2, SMO y SUFU, lo que lleva a una proliferación celular descontrolada y al crecimiento tumoral 54-56 (figura 1). Adicionalmente, las forminas, proteínas que regulan la dinámica del citoes-queleto de actina, induciendo su polimerización, se han identificado como reguladoras de la señalización Shh, influyendo en la actividad transcripcional de GLl2 y afectando la invasividad de las células tumorales 57. Así mismo, el coactivador oncogénico YAP1 (proteína asociada a Yes-1), regulado por la vía Hippo, que controla el crecimiento celular, también se ha documentado sobreexpresado en los meduloblastomas con señalización aberrante de Shh. Por su parte, YAP1 promueve la proliferación de los precursores de las neuronas granulares cerebelosas, consideradas células madre de estos tumores, y está asociado con la recurrencia tumoral 58.
Identificar los distintos niveles de variación en la señalización de Shh permitirá desarrollar enfoques terapéuticos más específicos y efectivos para tumores como el meduloblastoma. Por ejemplo, se ha demostrado que el metabolismo alterado del colesterol influye directamente en la señalización de Shh a través de la modulación de la interacción con el receptor PTCH259, donde Shh es secretado como una proteína madura tras su modificación postra-duccional con lípidos como colesterol y palmitato, lo que facilita su incorporación en complejos lipoproteicos que permiten su transporte y gradiente en el tejido durante el neurodesarrollo. n este contexto, la vía de biosíntesis del colesterol podría ser entonces un blanco terapéutico interesante y, de hecho, las estatinas, al inhibir la producción de colesterol, han mostrado efectos inhibitorios sobre la proliferación de células neoplásicas en el meduloblastoma dependiente de Shh 60-61. Así, nuevas estrategias terapéuticas podrían combinar inhibidores de la biosíntesis del colesterol con tratamientos dirigidos específicamente a la vía Shh.
Dentro de dicha vía, los receptores SMO y los factores de transcripción GLI son blancos clave para los tratamientos actuales. Inhibidores como sonidegib y vismodegib, aprobados por la Administración de Alimentos y Medicamentos ( FDA, según sus siglas en inglés) para el carcinoma basocelular, actúan sobre SMO y demuestran la viabilidad clínica de estas estrategias en otros tumores dependientes de Shh 62-63; sin embargo, la resistencia a estos inhibidores resalta la necesidad de explorar alternativas como los inhibidores de GLI. Entre otros objetivos moleculares, se encuentran los reguladores epigenéticos DNMT2 (DNA metiltransferasa-1) y SMARCA5 (miembro de la subfamilia 5 del complejo relacionado con SWl/SNF), cuya inhibición ha demostrado eficacia en líneas celulares sensibles y resistentes a inhibidores de SMO, sugiriendo la posibilidad de un blanco terapéutico más allá de la vía Shh 64-65.
Además, los meduloblastomas Shh impulsados por el oncogén N-Myc dependen de mecanismos específicos de plegamiento de proteínas, con funciones críticas de la chaperona molecular Hsp70, lo que abre nuevas posibilidades terapéuticas 66. En los meduloblastomas Shh con mutaciones en TP53, la hiperactivación de la vía mTORC2 , relacionada con el crecimiento celular, representa una vulnerabilidad, y su inhibición reduce la carga tumoral y agresividad, constituyendo un blanco prometedor 67. Por último, la fosfatasa EYA2 (Eyes Absent-1), altamente expresada en este subtipo y esencial para la señalización de Shh, representa otro objetivo relevante, ya que su inhibición interfiere con la vía Shh y limita el crecimiento tumoral 68.
Conclusiones
La vía Sonic Hedgehog (Shh) participa en dos procesos biológicamente opuestos, pero profundamente entrelazados: el desarrollo embrionario y la tumo-rogénesis. En condiciones fisiológicas, esta vía es esencial para la especificación neuronal, la migración celular y la proliferación de células progenitoras en etapas tempranas del neurodesarrollo; no obstante, su reactivación o desregulación en etapas posteriores de la vida se asocia con la iniciación y progresión de tumores como el meduloblastoma, especialmente en contextos pediátricos. Se estima que aproximadamente el 30 % de los casos de meduloblastoma presentan alteraciones genéticas en componentes de la vía Shh, subrayando su relevancia como eje etio-patogénico.
Los datos aquí revisados permiten concluir que, si bien existen diferencias regulatorias entre los contextos fisiológico y patológico, los paralelismos moleculares que comparten refuerzan la necesidad de profundizar en el estudio de esta vía como puente explicativo entre embriogénesis y oncogénesis. En este sentido, identificar los distintos niveles de variación en la señalización de Shh, incluidas sus modulaciones por el metabolismo lipídico, representa un aporte clave al diseño de terapias más precisas y eficaces. Por ejemplo, la influencia del colesterol en la interacción Shh-PTCH2 abre nuevas posibilidades para interferir farmacológicamente en la señalización aberrante, como lo demuestran los efectos sinérgicos de estatinas con inhibidores específicos de la vía.
A pesar de los avances en el desarrollo de fármacos dirigidos contra componentes como SMO o GLI, la aparición de resistencias terapéuticas exige explorar nuevas dianas moleculares. Entre estas se destacan los reguladores epigenéticos, las proteínas chaperonas implicadas en el plegamiento tumoral dependiente de N-Myc, las alteraciones en mTORC2 asociadas a mutaciones en TP53 y la fosfatasa EYA2. Todos ellos representan vulnerabilidades terapéuticas relevantes en el subtipo Shh del meduloblastoma.
Desde la perspectiva de los autores, esta revisión contribuye a cerrar una brecha conceptual existente entre el conocimiento básico sobre la vía Shh y su aplicación traslacional en oncología pediátrica. En particular, se propone una integración de mecanismos canónicos y no canónicos como base para comprender fenómenos de progresión tumoral y recurrencia posquimioterapia. Este enfoque permite no solo sistematizar los conocimientos actuales sobre la vía, sino también plantear nuevas preguntas de investigación orientadas a la caracterización de bio-marcadores, la estratificación de pacientes y el desarrollo de terapias combinadas más efectivas.

