










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.27 no.2 Bogotá May/Aug. 2009
PROTECCIÓN DE CULTIVOS
Rodrigo Gil1, 5, Alexander Smith2, Bernardo Chaves3, Kris Wyckhuys1, Clemencia Forero4 and Jaime Jiménez1
1 Centro de Investigaciones y Asesorías Agroindustriales, Facultad de Ciencias Naturales, Universidad de Bogotá Jorge Tadeo Lozano, Chia (Colombia).
2 Laboratorio de Control Biológico, Centro de Biotecnología y Bioindustria (CBB), Centro de Investigación Tibaitatá, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Mosquera (Colombia).
3 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
4 Departamento de Microbiología, Pontificia Universidad Javeriana, Bogota (Colombia).
5 Autor de correspondencia. rodrigo.gil@utadeo.edu.co
Received for publicaton: 13 February, 2009. Accepted for publication: 2 July, 2009
ABSTRACT
White mould: Sclerotinia spp., is the lettuce main disease in the Colombian high Andes. It causes yield losses of up to 70% and is primarily managed with chemical fungicides, while alternative control strategies are poorly used. Soil solarization and a set of chemical and bio-fungicides for Sclerotinia management efficacy in lettuce, were determined. Biofungicides included different doses of Allium sativum and Matricaria recutita extracts and Trichoderma koningiopsis suspensions. The fungicide procymidone was also evaluated. Field trials included single or combined treatments application. Non-linear modelling and AUDPC were employed to determine the efficacy of management strategies. Both S. sclerotiorum and S. minor caused white mould, being S. minor the dominant species. Solarization significantly lowered Sclerotia population in the upper soil layers and consequently lowered disease incidence. Highest levels of Sclerotinia control were achieved with solarization, particularly in combination with procymidone or T. koningiopsis (79.08% and 41.50% disease reduction, respectively). Non-linear modeling of disease epidemic progress curve proved a valuable alternative to AUDPC, which allowed multiple disease development parameters estimation and indirectly the efficacy of management strategies. This work should help the advance of environmentally-sound management of Sclerotinia spp. in lettuce, under the highly specific growing conditions of the high Andes.
Key words: high Andean tropics, plant extracts, procymidone, soilborne pathogens, Trichoderma koningiopsis.
RESUMEN
El moho blanco causado por Sclerotinia spp. es la principal enfermedad de la lechuga en la zona altoandina colombiana, causando hasta 70% de pérdidas. El principal control de esta enfermedad ha sido con fungicidas químicos, y poco se han usado otros métodos de control alternativos. En este estudio se determinó el efecto de la solarización y su combinación con biofungicidas y un fungicida químico para el control del hongo Sclerotinia spp. en lechuga. Para los biofungicidas se incluyeron diferentes dosis de los extractos de Allium sativum, Matricaria recutita y una suspensión de Trichoderma koningiopsis. El fungicida químico fue procymidona. Se empleó modelación no lineal y ABCPE para evaluar la eficacia de las estrategias de manejo. Se encontró que tanto S. sclerotiorum como S. minor causan moho blanco, siendo S. minor la especie dominante. La población de esclerocios en capas superficiales del suelo disminuyó significantemente con solarización, bajando así incidencia de la enfermedad y demostrando el mejor control, especialmente en combinación con procymidona o T. koningiopsis (79,08 y 41,50% de reducción, respectivamente). La modelación no lineal de la curva de progreso epidémico de la enfermedad, es una valiosa alternativa al ABCPE, que permitió estimar múltiples parámetros de desarrollo de la enfermedad e indirectamente la eficacia de estrategias de manejo. El presente trabajo contribuye en la elaboración de estrategias ambientalmente seguras para el control de Sclerotinia spp. en lechuga, en las condiciones específicas del trópico alto Andino.
Palabras clave: Trópico alto andino, extractos de plantas, promicidona, patógenos del suelo, Trichoderma koningiopsis.
Introduction
Throughout the world, one of the most important diseases of lettuce is white mould, caused by Sclerotinia minor Jagger or S. sclerotiorum (Lib.) de Bary (Abawi and Grogan, 1979; Subbarao, 1998). For example, in California (USA), yield losses ascribed to Sclerotinia spp. occasionally amount to 60% (Hao and Subbarao, 2005), while in Colombia, yield losses fluctuate between 20 and 70%. Colombian lettuce production is concentrated in the high Andean plateau, a region with highly-specific climatic conditions (Pérez, 2003). In recent years, Sclerotinia infestations have caused a sharp reduction of the lettuce acreage in various key production zones for this crop.
The symptoms in both species are: watery soft rot with development of white cottony mycelia on different parts of lettuce plants (Abawi and Grogan, 1979; Bolton et al., 2006). At given times during the disease cycle, survival structures such as sclerotia or apothecia are produced (Abawi and Grogan, 1979; Clarkson et al., 2004). Sclerotia can remain viable in the soil for up to 10 years (Adams and Ayers, 1979), which complicates management of Sclerotinia spp. A key prerequisite for successful Sclerotinia sp. management is the sound appreciation of disease development under specific (e.g., climatic, crop management) conditions.
Currently, Sclerotinia is most commonly managed using the fungicide procymidone (Patrício et al., 2006; Wilson et al., 2005). A sole reliance on chemical fungicides can lead to Sclerotinia resistance development, while also impacting biodiversity and interfering with key ecosystem services (Sorensen and Stewart, 2000). Additionally, Sclerotinia management is not based on a sound knowledge of causal agent or appreciation of extent of infestation. For the particular case of Sclerotinia, a single pest management strategy has not yielded satisfactory control and integrated tactics (combining physical, cultural, chemical or biological control) are urgently needed (Subbarao, 1998).
Recent research has spurred the development of alternatives for chemical fungicides. Soil disinfection through solarization has proven an effective strategy for control of pathogens, such as Sclerotinia (Phillips, 1990; Katan, 2000; Ferraz et al., 2003). Although solarization combined with applications of procymidone yields substantial reductions of S. minor incidence (Patrício et al., 2006), efficacy of such practice depends on local climatic conditions. Biological control of Sclerotinia has also been investigated, with various antagonistic fungi affecting white mould sclerotia both under in vitro as in field conditions (Jones and Stewart, 2000; Cheng et al., 2003; Rabeendran et al., 2006). Although successful biological control of Sclerotinia is mainly restricted to greenhouse production systems, Ávila and Gutiérrez (1991) report effective control of S. sclerotiorum with Trichoderma harzianum under field conditions in Colombia. Also, Trichoderma koningiopsis (Th003) Oudemans, has shown potential for Sclerotinia control (Cotes et al., 2007). Lastly, extracts from Allium sativum and Matricaria recutita have shown promise for management of fungi like Botrytis and Colletotrichum under in vitro conditions (Bianchi et al., 1997). However, the efficacy of several of these management alternatives remains to be investigated under the particular growing conditions of the Colombian high Andes.
Non-linear models are commonly used for the evaluation of disease development and quantification of host pathogen and environmental effects on fungal epizootics (Bowers and Kinkel, 1997). Such models allow the analysis of disease development and can help assess the efficacy of disease management strategies (Campbell and Madden, 1990). Non-linear models could therefore serve to determine the efficacy of management alternatives for Sclerotinia sp. in lettuce under specific environmental (i.e., climatic) conditions.
At present, Colombian lettuce growers mainly rely on chemical fungicides and largely lack workable alternatives for Sclerotinia disease management. The aim of this article was assessment of the effects of soil solarization and application of chemical or bio-fungicides on Sclerotinia spp. in lettuce in the Colombian high Andes. The specific objectives of this work are to evaluate the effect of two plant extracts and T. koningiopsis suspensions in a lettuce crop under open-field conditions; and to contrast the use of non-linear modelling with that of area under disease progress curve (AUDPC).
Materials and methods
Study area: Research was carried out in the municipality of Cota (4o49'05'' N and 74o07'20'' W) (Cundinamarca, Colombia). Specific climatic conditions of this locality are: annual mean temperature: 13.7°C, annual rainfall: 700 mm, and altitude: 2,547 m. Experiments were conducted in fields planted (700 m2) with lettuce (Lactuca sativa var. Coolguard®), 11 plants/m2, between January and May 2006. Lettuce fields with high Sclerotinia spp. incidence were previously selected in November 2005.
Sample collection and disease identification. A total of 20 diseased plants were randomly collected in a selected field for experimentation and taken to the Phytopathology laboratory at the "Centro de Investigaciones y Asesorías Agroindustriales" (CIAA) (Chia, Cundinamarca) to confirm identity of pathogens. Fungal samples were isolated in potato dextrose agar (PDA) and pathogens were identified based on macro and microscopic characteristics. For Sclerotinia identification, we used identification keys developed by Kohn (1979). In the laboratory (22.5±0.5°C, 92±5% RH), we described morphological and developmental characteristics of the collected fungal species.
In addition, soil samples of selected field were taken and Sclerotia found were characterized morphologically (e.g. number of sclerotia produced) and their infectivity was quantified through Koch's postulates trials (Agrios, 2005). In these trials, we used viable sclerotia obtained from soil samples and reproduced on PDA in the laboratory. Next, lettuce plants were inoculated with agar discs containing isolates of the collected fungi. A total of 20 replicates were conducted per fungal species.
Field experimental design: In January 2006, a former lettuce field (700 m2) was superficially tilled, and 42.0 x 1.5 m beds were established. The field experiment was set up as a randomized split-plot design with five replications. The main treatment was solarization, while subplots consisted of seven different treatments: plant extracts of garlic (Allium sativum L.) and chamomile (Matricaria recutita L.) at two concentrations (1 and 25 g L-1), T. koningiopsis WG (1 g L-1), procymidone (1 g L-1) and an absolute control (without applications). For the application of treatments was used a water volume of 300 L. Applications in each treatment were conducted on the basis of each plant. Each bed was divided into two equal 21 m2 plots, excluding the area of the edges, which were assigned to the main treatment (i.e., with or without solarization); each subplot had an area of 3 m2.
For the solarization treatment, a 152 μm thick transparent plastic cover was placed during four weeks on each main plot, while no-solarization beds were left bare. Throughout the experiment, we recorded climatic data with a climate station (Micrometros®) and monitored soil temperature profiles up to 5 cm deep with T-type (coppec-constantan) thermocouples coupled to a remote datalogger (EJ-2E, Escort DLS, Auckland, New Zealand) probes at 10 min intervals. Daily average soil temperature was calculated for subset data at major radiation hours (10 a.m. and 3 p.m.) and used to determine the solarization effect on soil temperature. After four weeks, plastic cover was removed from the solarization plots. Furthermore, we took seven soil samples up to 25 cm of depth (100 g) per main plot and recorded the number of sclerotia per soil sample through sieving (2 mm, 1 mm and 250 μm) and visual counts. Previous verifications of necessary assumptions over data, a Student's t-test were used to determine statistical differences in temperature and number of viable sclerotia between treatments: solarization and non-solarization.
Biofungicides: Garlic and chamomile extracts have shown to inhibit growth of S. clerotiurum mycelia under in vitro conditions (Espinosa and Niño, 2008). Both plant extracts were elaborated using a "Purines" method. "Purines" are obtained through anaerobic fermentation of selected plant tissues. For "garlic-purin" were used cloves of garlic, while for "chamomile-purin" were used flowers and leaves (Jiménez et al., 2007). T. koningiopsis (Th003) is a biopesticide prototype developed by the biological control lab of the Bioindustry and Biotechnology Centre of the Colombian Corporation for Agricultural Research-Corpoica, the dose used was 1x106 spores/mL. Prior to experimental use, we determined the colony forming units (cfu) per cc of the fungal suspension.
Chemical product: The chemical fungicide treatment was procymidone application, a product that is commonly used by local farmers (Jiménez et al., 2007). Within each of the experimental plots, lettuce seedlings were planted with four open leaves at 0.3 x 0.3 m distance. Plant extracts and fungal suspensions were applied on a weekly basis following planting, while procymidone was applied every two weeks until 30 days prior to harvest. Prior to planting, the fungal suspension was also applied at a weekly basis on lettuce seedlings and upon planting. At planting, we incorporated 1 g of T. koningiopsis WG.
Data collection and analysis: In each treatment, all plants were visually checked and were recorded the number of diseased plants at a weekly basis. Temporal patterns of Sclerotinia incidence were adjusted to different mathematical models (Campbell and Madden, 1990; Xu, 2006), obtaining the best fit with the logistic model presented in equation (1).
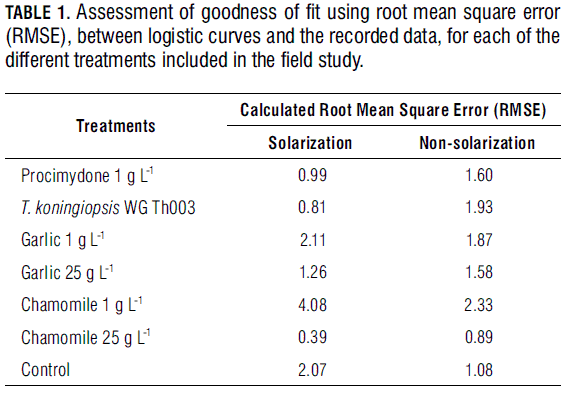
Where Yi is the number of diseased plants during the ith observation, α the maximum disease incidence (horizontal asymptote), e the natural log basis, K the slope in the inflection point, dat the number of days after planting and γ the time (in dat) at which the curve inflection point occurs. For each treatment, α, K and γ were calculated. To determine the logistic curves goodness of fit to the recorded data, the root mean square error was calculated (RMSE). For Sclerotinia disease development within each treatment, the area under the disease progress curve (AUDPC) was determined using the trapezoidal integration method (Campbell and Madden, 1990; Xu, 2006) as indicated in equation (2).

Where, Yi is the number of diseased plants at the ith observation, ti is days after planting, and n the total number of observations.
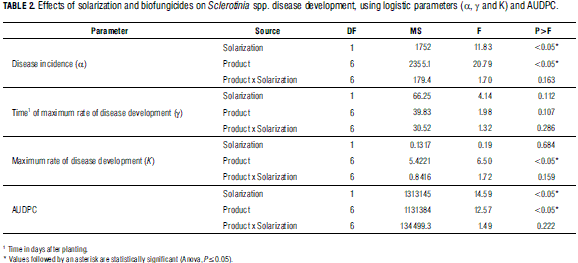
Statistical differences for α, K, γ and the AUDPC between the various treatments were determined, using a factorial Anova, followed by a Tukey HSD post-hoc analysis. Statistics were carried out using statistical software package R (R Development Core Team, 2006).
Results
The following results with respect to white mould are reported at first under the high Andes conditions. Laboratory observations indicate that two different species of Sclerotinia were present: S. minor and S. sclerotiorum. Sclerotinia forming patterns differed between both species: either in concentric circles (S. minor) or random (S. sclerotiorum). In laboratory, cultures of both species, the presence of Fusarium spp. colonies that limited or inhibited Sclerotinia spp. development was observed. In Koch's postulate trials, 85% and 75% of plants inoculated with S. minor and S. sclerotiorum sclerotia respectively, showed typical disease symptoms (Subbarao, 1998; Ekins et al., 2005), identical to the ones observed in the field.
Effect on soil temperature: Temperatures in solarization plots (45.3±10.8°C) were significantly higher than in plots without solarization (29.3±3.6°C) (Student's t-test, t =79.16, df= 4734, P<0.0001) (Fig. 1). Upon establishment of the lettuce crop, the number of sclerotia in solarization plots was significantly lower than in non-solarization plots (Student's t-test, t =3.90, df=23, P=0.0018). In plots without solarization, we recorded 3.91±0.57 viable sclerotia/100 g of soil, while solarization plots had 0.80±0.31 viable sclerotia/100 g of soil.
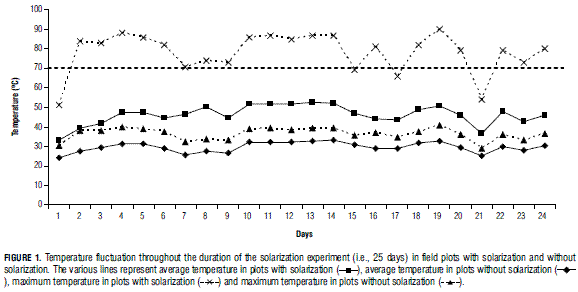
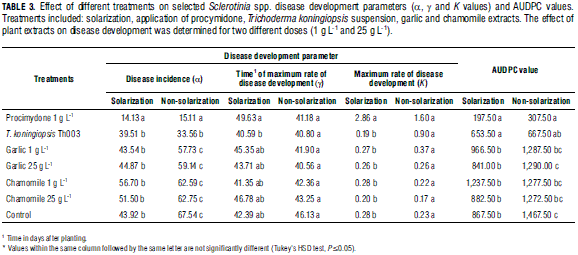
Effect on disease development: Use of functions fitted to disease curve progress for assessment management practices is one of key the foci of the study. In general, logistic curves fit well to the recorded data (Tab. 1). Solarization treatments had a significant effect on maximum Sclerotinia incidence (α), while biofungicides significantly affected both α and K (Tab. 2). The absence of significance for the interaction between solarization and product (Tab. 2) indicates that the effects are independent; furthermore, Fig. 2 suggest that the combination of solarization and product is different to the completely untreated (nonsolarization) control. Also, time until maximum disease rate (γ) did not differ between treatments. Although disease symptoms generally appeared after approx. 20 d, K was reached 40-50 d after planting. Maximum disease rate (between 40 and 50 d) was reached independent of treatment (Fig. 2). The solarization treatment significantly affected disease incidence (α), compared to non-solarization plots (Tab. 3). Maximum disease incidence (α) was significantly lower in the procymidone treatment than in plots where T. koningiopsis or plant extracts were applied. Also, significant differences were found for α between "T. koningiopsis" and "plant extracts" treatments. The procymidone treatment was also typified by the highest rate of Sclerotinia development (K). Identical results were obtained using an interpretation of the AUDPC (Tab. 2).
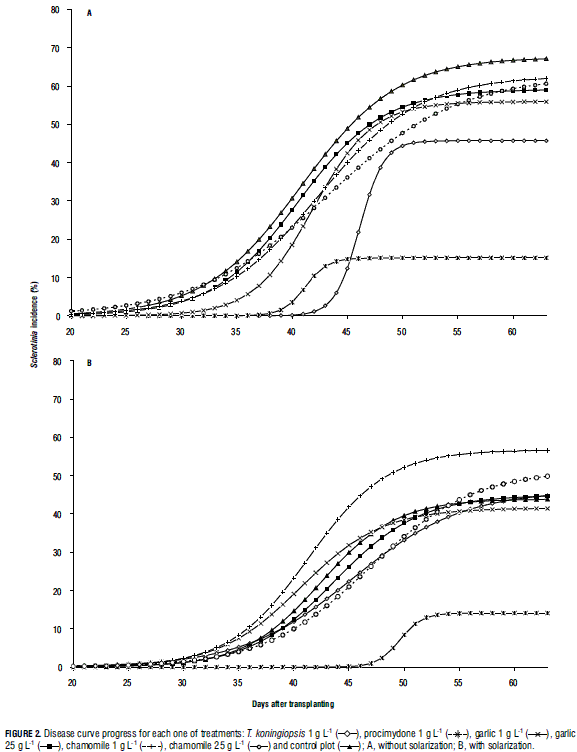
Discussion
Experimental evidence showed that white mould in lettuce in the Colombian high Andes is caused by both S. minor and S. sclerotiorum. Although solarization reduced the number of viable sclerotinia, procymidone and T. koningiopsis were most effective in lowering white mould disease incidence. Except for use of T. koningiopsis, the combined application of solarization and biofungicides yielded a greater level of Sclerotinia spp. control. However, Hoyos-Carvajal et al. (2008) suggest that mycoparasitic capacity of Trichoderma species varies according to pathogen. Therefore, the results of this study would be restricted to the dominant species, S. minor and vary in areas where the dominant species is S. sclerotiorum.
Results reported in this study only emanate from experiments conducted during one cropping cycle, but with all treatments repeated five times. Given the difficulty in locating farmer-owned fields with high Sclerotinia incidence and setting up trials with farmers' consent, we were unable to repeat the experiment for another cycle. Nevertheless, the results were deeming indicative of the different disease management strategies effect, under the specific environmental conditions of the study region.
Although S. sclerotiorum has been reported from the Colombian high Andes (Avila and Gutierrez, 1991; Arias et al., 2007), this is the first record of S. minor in this region. The work showed that white mould in lettuce is caused by both species, but that S. minor was the dominant species in most lettuce fields. Upon presence of both Sclerotinia spp. in a given site, one species commonly causes the majority of disease symptoms (Subbarao, 1998). Such results could indicate that in the study region, S. minor likely is the main cause of white mould. Also, as S. minor produces more sclerotia than S. sclerotiorum, the former species may more likely cause fungal epidemics. Characterization of the Sclerotinia species complex associated with white mould in lettuce has broader implications for management of this disease.
Our work also hinted the occurrence of (natural) biological control of Sclerotinia in lettuce fields, with a potential role of naturally-occurring Fusarium sp. colonies. In the laboratory, these colonies inhibited Sclerotinia mycelia growth, indicating that Sclerotinia populations could be naturally regulated by other fungi (Adams and Ayers, 1979; Rodriguez et al., 2006). Such interactions may eventually be manipulated to improve Sclerotinia biological control in field conditions. However, additional research is needed to determine the specific interactions between Fusarium spp. and both Sclerotinia species. Additionally, it is necessary identify the species of Fusarium and to quantify their potential as biological control agents.
The effectiveness of solarization greatly depends on overall weather conditions, and more specifically on ambient temperature or solar radiation. In general, solarization treatments are effective when temperatures surpass 70°C for >30 min (Stapleton et al., 2000; Flint and Gouveia, 2001). During the field trials conducted in the present study, temperatures in solarization plots exceeded 70°C for at least 4 h daily throughout the experiment, and this practice therefore successfully disinfected soil. The observed reduction of white mould incidence following solarization, is in agreement with reports of its effect on pestiferous fungi in general (Stapleton et al. 2000; Ferraz et al. 2003) and specifically on lettuce plants infected by S. minor (Patricio et al., 2006). Also, solarization may easily be adopted in the region, as plastics are abundantly available as waste materials from the local floriculture sector. As experiments were conducted during January and May the efficacy of solarization as a disease management practice will need to be validated during the remainder of the year.
Although solarization has considerable potential for soil disinfection and Sclerotinia control, its effect on beneficial soil-inhabiting organisms remains controversial. Increased periods of solarization generally allow toxic substances to accumulate and may have sub-lethal effects on beneficial organisms (Flint and Gouveia, 2001). On the other hand, certain antagonistic organisms are heat-resistant and may even be activated by solarization practices (Ferraz et al., 2003; Stevens et al., 2003; Porras et al., 2007). Considering the potential importance of Sclerotinia biological control (i.e., through action of Fusarium spp.), solarization treatments may require continuous monitoring and finetuning. More specifically, long-term effects of elevated temperatures on beneficial fungi need to be investigated and their impact on Fusarium inoculum density and viability waits to be determined.
The high efficacy of procymidone treatment reported in this study corresponds with previous research (Wilson et al., 2005; Patricio et al., 2006; Arias et al., 2007; Cotes et al., 2007). However, the independent effect when using procymidone in solarized plots is somewhat surprising. For example, Patricio et al. (2006) reported increased control of S. minor when combining solarization with procymidone treatments. Contrarily, Arias et al. (2007) reported vastly lower levels of S. sclerotiorum control using the same experimental set-up in the high Andes region. This discrepancy between both studies could be attributed to the fact that Arias et al. (2007) carried out their research during a different time of the year (with dissimilar climatic conditions) and in plots with low Sclerotinia infestation. Results in this study are similar to those of Arias et al. (2007), with respect to low efficacy of combined use of solarization and procymidone.
The level of Sclerotinia spp. control by T. koningiopsis Th003 was lower than that described by Escande et al. (2002) for T. koningiopsis strains TK1 and TK2 and for T. harzianum (Avila and Gutierrez, 1991; Jones and Stewart, 2000). Such differences could be attributed to high Sclerotinia spp. infestation levels in the experimental plots or the particular formulation of the fungal suspension, considering it was a prototype. Nevertheless, this work showed significant bio-control action of T. koningiopsis Th003 against Sclerotinia spp., with this bio-pesticide eventually constituting a viable alternative to procymidone. The description of Sclerotinia spp. control with T. koningiopsis is one of few reports of Sclerotinia biological control under field conditions (Alabouvette et al., 2006). Along this line, although little is known about Trichoderma spp. behaviour in the field, results hint at a reduced activity of T. koningiopsis in solarization plots. At the end of the experiment, T. koningiopsis cfus were lower (6.91 cfu/mL) in solarization plots than in non-solarization plots (8.57 cfu/mL), which may indicate that solarization eventually compromises the efficacy or viability of T. koningiopsis. Secondary succession of microorganisms, after solarization, may affect the establishment of T. koningiopsis.
Although garlic and chamomile extracts effectively inhibited Sclerotinia growth in the laboratory, no control was reported under field conditions. This lack of Sclerotinia control can be ascribed to two factors: firstly, certain environmental factors could cause a quick degradation of the active ingredients and impede the action of plant extracts under field conditions. Secondly, the artisan preparation of plant extracts may allow contamination with micro-organisms, which may cause degradation of active ingredients. Additional research is needed to determine the exact causal factors of this lack of control of plant extracts.
Modelling approaches are routinely used for evaluating disease development and determining the effect of various factors (e.g., disease management) on fungal epidemics. AUDPC values are valuable descriptors of disease development, and have been widely used in plant pathology (Campbell and Madden, 1990; Van Maanen and Xu, 2003; Xu, 2006). However, results indicated that a more useful alternative to AUDPC analyses is the adjustment of the epidemic progress curve to a theoretical model, which allowed estimation of multiple parameters of disease development over time. Computation of these parameters then provides the basis for informed pest management decision making, such as the targeting of fungicidal sprays. Field results indicated that, in order to be effective, control measures should be taken prior to γ, i.e. during the first 40 d after planting.
This research is of crucial importance for the formulation of disease management protocols against Sclerotinia in Colombia and eventually throughout the Andes. Solarization and application of biofungicides (i.e., T. koningiopsis) have proven to be viable alternatives to chemical control of white mould in lettuce, while being safe, low-cost and environmentally-sound technologies which can easily be adopted by local small-scale vegetable farmers.
Acknowledgements
We would like to thank R. García and his family for granting access to their farm. We are grateful to L. Espinosa for help with in vitro assays of plant extracts, the Biotechnology and Bioindustry Center of Corpoica, and O. Duarte for his invaluable help in revising this manuscript. This research was supported by a grant from the Universidad de Bogota Jorge Tadeo Lozano and Instituto Colombiano para el Avance de la Ciencia y la Tecnologia "Francisco Jose de Caldas"-Colciencias.
Cited referentes
Abawi, G.S. and R.G. Grogan. 1979. Epidemiology of diseases caused by Sclerotinia species. Phytopathol. 69, 899-904. [ Links ]
Adams, P.B. and W.A. Ayers. 1979. Ecology of Sclerotinia species. Phytopathol. 69, 896-898. [ Links ]
Agrios, G. 2005. Plant pathology. 5th ed. Elsevier Academic Press, New York, NY. [ Links ]
Alabouvette, C., C. Olivain, and C. Steinberg. 2006. Biological control of plant diseases: the European situation. Eur. J. Plant Pathol. 114, 329-341. [ Links ]
Arias, L., A. Táutiva, W. Piedrahíta, and B. Chaves. 2007. Evaluación de tres métodos de control del Moho blanco (Sclerotinia sclerotiorum (Lib.) de Bary) en lechuga (Lactuca sativa L.). Agron. Colomb. 25(1), 131-141. [ Links ]
Ávila, C. and A. Gutiérrez. 1991. Control biológico de Sclerotinia sclerotiorum (Lib.) de Bary en lechuga (Lactuca sativa L.). I. Selección de cepas antagónicas y medio de cultivo. Revista ICA 26, 35-42. [ Links ]
Bianchi, A., A. Zambonelli, A. Zechini, and F. Bellesia. 1997. Ultrastructural studies of the effects of Allium sativum on phytopathogenic fungi in vitro. Plant Dis. 81, 1241-1246. [ Links ]
Bolton, M.D., B.P. Thomma, and B.D. Nelson. 2006. Pathogen profile: Sclerotinia sclerotiorum (Lib.) de Bary: biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 7, 1-16. [ Links ]
Bowers, J.H. and L.L Kinkel. 1997. Interactive modeling of disease progress curves. pp. 20-23. In: Franci, L.J. and D.A. Neher (eds.). Exercises in plant disease epidemiology, APS Press, St. Paul, MN. [ Links ]
Campbell, C.L. and L.V. Madden. 1990. Introduction to plant disease epidemiology. John Wiley and Sons, New York, NY. [ Links ]
Cheng, J., D. Jiang, X. Yi, Y. Fu, G. Li, and J. Whipps. 2003. Production, survival and efficacy of Coniothyrium minitans conidia produced in shaken liquid culture. FEMS Microbiol. Lett. 227, 127-131. [ Links ]
Clarkson, J.P., K. Phelps, J.M. Whipps, C.S. Young, J.A. Smith, and M. Watling. 2004. Forecasting Sclerotinia disease on lettuce: toward developing a prediction model for carpogenic germination of sclerotia. Phytopathol. 94, 268-279. [ Links ]
Cotes, A.M., C.A. Moreno, L.F. Molano, L.F. Villamizar, and W. Piedrahíta. 2007. Prospects for integrated management of Sclerotinia sclerotiorum in lettuce. IOBC/wprs Bulletin 30 (6), 391-394. [ Links ]
Ekins, M.G., E.A.B. Aitken, and K.C. Goulter. 2005. Identification of Sclerotinia species. Australasian Plant Pathol. 34, 549-555. [ Links ]
Escande, A., F. Laich, and M. Pedraza. 2002. Field testing of honeybee- dispersed Trichoderma spp. to manage sunflower head rot (Sclerotinia sclerotiorum). Plant Pathol. 51, 346-351. [ Links ]
Espinosa, L. and N. Niño. 2008. Evaluación in vitro de extractos vegetales para el control de Sclerotinia sclerotiorum. (Lib. de Bary) y Botrytis cinerea (Pers.) en lechuga (Lactucca sativa). Editorial Universidad Jorge Tadeo Lozano, Bogotá. Resultados de Investigación 1(8), 1-24. [ Links ]
Ferraz, L.C., A. Bergamin, L. Amorim, and L.C. Nasser. 2003. Viabilidade de Sclerotinia sclerotiorum após a solarização do solo na presença de cobertura monta. Fitopatol. Bras. 28, 17-26. [ Links ]
Flint, L.M. and P. Gouveia. 2001. IPM in practice: principles and methods of integrated pest management. University of California, Oakland, CA. [ Links ]
Hoyos-Carvajal, L., P. Chaparro, M. Abramsky, I. Chet, and S. Orduz. 2008. Evaluación de aislamientos de Trichoderma spp. contra Rhizoctonia solani y Sclerotium rolfsii bajo condiciones in vitro y de invernadero. Agron. Colomb. 26(3), 451-458. [ Links ]
Hao, J.J. and K.V. Subbarao. 2005. Comparative analyses of lettuce drop epidemics caused by Sclerotinia minor and S. sclerotiorum. Plant Dis. 89, 717-725. [ Links ]
Jiménez, J., R. Gil, L.S. Fuentes, C. Barreto, L. Espinosa, M. Romero, D. Carrillo, M. García, and H. Ubaque. 2007. Aportes al manejo integrado de plagas en cultivos ecológicos de hortalizas con énfasis en cultivos de lechuga. Editorial Fundación Universidad de Bogotá Jorge Tadeo Lozano, Bogota. [ Links ]
Jones, E.E. and A. Stewart. 2000. Selection of mycoparasites of sclerotia of Sclerotinia sclerotiorum isolated from New Zealand soils. New Zeal. J. Crop Hort. 28, 105-114. [ Links ]
Katan, J. 2000. Physical and cultural methods for the management of soil-borne pathogens. Crop Prot. 19, 725-731. [ Links ]
Kohn, L. 1979. Delimitation of the economically important plant pathogenic Sclerotinia species. Phytopathol. 69, 881-886. [ Links ]
Patrício, F.R., C. Sinigagliaa, B.C. Barrosa, S.S. Freitas, J. Tessarioli, H. Cantarellab, and R. Ghini. 2006. Solarization and fungicides for the control of drop, bottom rot and weeds in lettuce. Crop Prot. 25, 31-38. [ Links ]
Pérez, S. 2003. La pudrición blanda de la lechuga causada por el hongo Sclerotinia sclerotiorum (Lib.) de Bary o S. minor Jagger. Ph.D. thesis. Agronomy Faculty, National University of Colombia, Bogota. [ Links ]
Phillips, A.J. 1990. The effects of soil solarization on sclerotial populations of Sclerotinia sclerotiorum. Plant Pathol. 39, 38-43. [ Links ]
Porras, M., C. Barrau, F.T. Arroyo, B. Santos, C. Blanco, and F. Romero. 2007. Effects of soil solarization and Trichoderma on strawberry production. Plant Dis. 91, 142-146. [ Links ]
Rabeendran, N., E.E. Jones, D.J. Moot, and A. Stewart. 2006. Biocontrol of Sclerotinia lettuce drop by Coniothyrium minitans and Trichoderma hamatum. Biol. Control 39, 352-362. [ Links ]
R Development Core Team. 2006. R: a language and environment for statistical computing. In: R Foundation for Statistical Computing, URL http://www.R-project.org/; consulted: june 2009. [ Links ]
Rodríguez, M.A., G. Cabrera, and A. Godeas. 2006. Cyclosporine a from a nonpathogenic Fusarium oxysporum suppressing Sclerotinia sclerotiorum. J. Appl. Microbiol. 100, 575-586. [ Links ]
Sorensen, A. and P.A. Stewart. 2000. Factors affecting the adoption of new technologies. In: Kennedy, G.G. and B.T. Sutton (eds.). Emerging technologies for integrated pest management: concepts, research, and implementation. APS Press, St. Paul, MN. [ Links ]
Stapleton, J.J., C.L. Elmore, and J.E. DeVay. 2000. Solarization and biofumigation help disinfest soil. Calif. Agric. 46, 42-45. [ Links ]
Subbarao, K.V. 1998. Progress toward integrated management of lettuce drop. Plant Dis. 82, 1068-1078. [ Links ]
Van Maanen, A. and X.M. Xu. 2003. Modelling plant disease epidemics. Eur. J. Plant Pathol. 109, 669-682. [ Links ]
Wilson, C.R., J. de Little, J.A. Wong, P.J. Schupp, and L.J. Gibson. 2005. Adjustment of soil-surface pH and comparison with conventional fungicide treatments for control of lettuce drop (Sclerotinia minor). Plant Pathol. 54, 393-400. [ Links ]
Xu, X. 2006. Modelling and interpreting disease progress in time. pp. 215-238. In: Cooke, B.M., D.G. Jones, and B. Kaye (eds.). The epidemiology of plant diseases. Springer, Dordrecht, The Netherlands. [ Links ]

