










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.33 no.3 Bogotá Sep./Dec. 2015
https://doi.org/10.15446/agron.colomb.v33n3.50237
Doi: 10.15446/agron.colomb.v33n3.50237
1 Departamento de Ciencias Agronómicas, Facultad de Ciencias Agrarias, Universidad Nacional de Colombia. Medellin (Colombia). tmsaldanav@unal.edu.co
Received for publication: 23 June, 2015. Accepted for publication: 17 November, 2015.
ABSTRACT
It is necessary to study the photosynthate distribution pattern and allocation in Guaneña and Latina potato varieties, which come from the Colombia variety and are relatively new in agriculture, for potato crops in Colombia. The aim of this study was to determine the distribution behavior of dry matter among the organs that make up the potato plant in order to determine the key moments in the crop cycle, where the filling of the tubers takes place, and to establish the pattern of biomass distribution in these varieties. Three consecutive experiments were performed at the "EstaciónAgraria Paysandú" in Santa Elena (Medellin - Antioquia), at 2,538 m a.s.l. with average temperatures of 13.6, 14.6 and 13.81°C. The field trials were established under a random complete blocks design with a 3x2 factorial layout and five replications. The first factor corresponded to the potato varieties and the second factor corresponded to the two fertilization levels: 7 and 21 g of fertilizer per planting site, representing 259.25 and 777.77 kg of fertilizer per ha. The higher fertilizer dose is the one normally used by farmers. An NPK fertilizer compound (10 - 20 - 20 - 5(S)) was used. There were 30 experiment units with an area of 18.27 m2 (seven 2.7 m furrows separated by 0.90 m); the distance between the plants was 0.30 m, for a density of 37,037 planting sites per ha. Weekly samplings were taken, a whole plant per experiment unit. The direct measuring evaluated variables for the dry weights of each organ, which were used to obtain curves and the distribution pattern. The development time was evaluated in accumulated growing degree-days and threshold temperatures of 2 and 29°C were used. In all of the varieties and the two fertilization treatments, most of the assimilates in the early stages of the cycle were concentrated in the roots, leaves and stems; once the tuber formation started, the leaves and stems started to allocate dry matter towards the filling of the accumulation organs.
Key words: tubers, source sink relations, photosynthesis, nutrient transport, Guaneña variety, Latina variety.
RESUMEN
Se hace necesario estudiar el patrón de distribución y asignación de los fotoasimilados en variedades de papa criolla Guaneña y Latina que provienen de la variedad Criolla Colombia y son relativamente nuevas en la agricultura en torno al cultivo de la papa en Colombia. El objetivo de este trabajo fue conocer el comportamiento de la distribución de la materia seca entre los órganos que conforman la planta de papa, con el fin de determinar los momentos claves en el ciclo de cultivo donde se está llevando a cabo el inicio del llenado de tubérculos y establecer el patrón de distribución de biomasa en las variedades mencionadas. Se realizaron tres experimentos consecutivos en la Estación Agraria Paysandú, ubicada en el Corregimiento de Santa Elena (Medellín - Antioquia) a 2.538 msnm. Se utilizó un diseño experimental de bloques completos al azar con dos niveles de fertilización correspondientes a 7 y 21 g de fertilizante por sitio que representan 259,25 y 777,77 kg de fertilizante/ha, y cinco repeticiones. La unidad experimental fue un área de 18,27 m2 (7 surcos de 2,7 m de largo), la distancia entre surcos fue de 0,90 m, la distancia entre plantas fue de 0,30 m para una densidad de 37,037 sitios de siembra/ha. Se hicieron muestreos semanales correspondientes a una planta entera por unidad experimental. Las variables evaluadas de medición directa fueron los pesos secos de cada órgano y a partir de estos pesos se determinaron las curvas que determinan el patrón de distribución. El tiempo de desarrollo se evaluó en grados día acumulados y se utilizaron las temperaturas umbrales de 2 y 29°C. En todas las variedades en los dos tratamientos de fertilización la mayor parte de los asimilados en las fases iniciales del ciclo se concentran en las raíces, hojas y tallos y una vez se inicia la formación de tubérculos, las hojas y los tallos vierten la materia seca acumulada hacia el llenado de los órganos de acumulación.
Palabras clave: tubérculos, relación fuente sumidero, fotosíntesis, transporte de nutrientes, variedad Guaneña, variedad Latina.
Introduction
The plants, as a result of the interaction between the photosynthesis, respiration, assimilate transport and production, hydric relationships and mineral nutrition accumulated and distributed dry matter in their organs and increased their volume, length or area as a result of the cell differentiation, expansion and division, i.e, plants grow and develop to fully comply with the vegetative and reproductive phase (Larcher, 2003; Fourcaud et al., 2008; Taiz and Zeiger, 2010).
There are regulation centers involved in the partition control of photoassimilates through which enzyme activities and the f low of metabolic pathways can change (Warren, 1981). In the assimilate builder organs or assimilate sources, the fixed carbon is shared according to the physiological needs of the plant, such as the RuBisCO reconstitution involved in the Calvin cycle, in the photo-respiration processes, and in the phosphoglyceraldehyde production for starch emergence located inside the chloroplasts or inside the sucrose that is sent to the cytoplasm of cells (Taiz and Zeiger, 2010).
Plants produce carbohydrates (sugars) in their leaves by photosynthesis; and consumer organs and accumulators known as dumping organs that require products of this reaction to grow and develop (Foyer and Paul, 2001); in the case of potatoes, these organs are the roots, stolons, f lowers, fruits and mainly tubers. The filling and formation of tubers will depend primarily on the availability of previously produced and temporarily stored assimilates in the stems and leaves, their capacity to store these sugars in their tissues, genetic aspects of the plant and environmental conditions under which they are being developed (Mora et al., 2005). This partitioning of photosynthates to tubers will then present a quantifiable result of growth and development that is also influenced by cultural practices (Foyer and Paul, 2001).
In potato crops, a good translocation of assimilates from the source to the tubers is decisive for the final production, along with an understanding of the patterns of that biomass allocation between the organs of the cultivated varieties and the effect of environmental conditions that improve the selection of materials for a location or general purpose (Van Heemst, 1986; Tekalign and Hammes, 2005a).
Dry matter accumulation is used as an indicator for growth because of its considerable economic significance since, in the case of the potato, the tuber is the organ that has photoassimilate storage mainly as starch (Tekalign and Hammes, 2005b).
Among other studies on the distribution behavior of photoassimilates, there is one by Kooman and Rabbinge (1996) in which the influence of temperature, photoperiod and radiation on potato crop growth, Solanum tuberosum L., and cycle length was determined; the authors emphasized that, in the first stage of the crop, the biomass is distributed preferentially in leaves and stems; in the second phase, the mass f low toward new and existing leaves is reduced because the formation and filling of tubers starts and, in the third part of the cycle, the tubers are the organs with the highest filling capacity and the only organs that continue to grow until the harvest day, while the aerial part of the plant starts to decay.
Ñústez et al. (2009) assessed the accumulation and distribution of dry matter in the organs of the varieties Diacol Capiro, Pastusa Suprema, Betina and Esmeralda from the S. tuberosum species and found differences in the dry matter accumulation in the stems, leaves and tubers; these differences corresponded to the moment with greater accumulation and duration of biomass in a particular organ.
Santos (2010) conducted an evaluation of source-demand in terms of dry matter partition and allocation in the states of vegetative and reproductive development of the diploid potato varieties Colombia, Guaneña, Latina and Galeras. In this study, the varieties Guaneña and Galeras stood out because they allocated the largest proportion of assimilates to the tubers, resulting in a higher yield, between 25,6 and 49,6 t ha-1.
In Mexico, Aguilar et al. (2006) conducted a growth analysis of the source-demand of two potato varieties (S. tuberosum), the results showed that the variety Alpha allocated 56% of its dry matter to the tubers. Meanwhile, Mora et al. (2005) evaluated five potato genotypes and determined the behavior of the accumulation and distribution of biomass.
Based on the above context, the aim of this study was to determine the behavior and distribution pattern of biomass in the diploid potato varieties Colombia, Guaneña and Latina during their crop cycle.
Materials and methods
Location
Three consecutive growing cycles of potato varieties in Santa Elena in eastern Antioquia (Colombia), at an altitude of 2,538 m a.s.l. (6°12'36'' N and 75°30'25'' O) and with average temperatures of 13.6, 13.81 and 14.6°C, were established in the field. According to the ecological conditions, this area is classifies as a lower montane forest zone (bh-MB) (Holdridge, 1967).
Experiment design
The field trials were established under a random complete blocks design with a 3x2 factorial layout and five replications. The first factor corresponded to the three varieties of potato and the second factor corresponded to the two fertilization levels: 7 and 21 g of fertilizer per planting site, representing 259.25 and 777.77 kg ha-1 of fertilizer; the higher dose of fertilizer corresponded to the one normally used by farmers. An NPK fertilizer compound was used (10 - 20 - 20 - 5(S)). There were 30 experiment units with an area of 18.27 m2 (seven 2.7 m furrows separated by 0.90 m); the distance between the plants was 0.30 m, for a density of 37,037 planting sites per hectare.
Variables evaluated
A fully functional plant was extracted weekly from each experiment unit to measure the variables listed below:
Dry matter: each plant was dissected, separating each organ individually and packing them in paper bags. These samples were placed in a drying oven at 80°C until constant dry weight was obtained. With this method, the dry weight of the leaves (DWL), stems (DWS), roots (DWR), stolons (DWST), tubers (DWTB), f lowers (DWF), and fruits (DWFR) and the sum of the weights of the total dry weight of the plant (DWT) (Eq. 6) was obtained.
Time: the development time was evaluated in terms of thermal time on accumulated degree-days (ADD), which takes into account the weather information of where the experiments are carried out. To obtain this information, a WatchDog 2900ET (Spectrum Technologies, Plansfield IL) weather station was used, which recorded the temperature, sunshine, rainfall and relative humidity every 15 min.
Statistical analysis
The incomplete gamma distribution model was used (Wood, 1967) (Eq. 1) since it is consistent with the modeling theory of crops based on models with lags (Manetsch, 1976; Vansickle, 1977; Gutierrez, 1996; Rodríguez et al., 2011).

This model can be linearized by the expression:
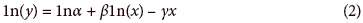
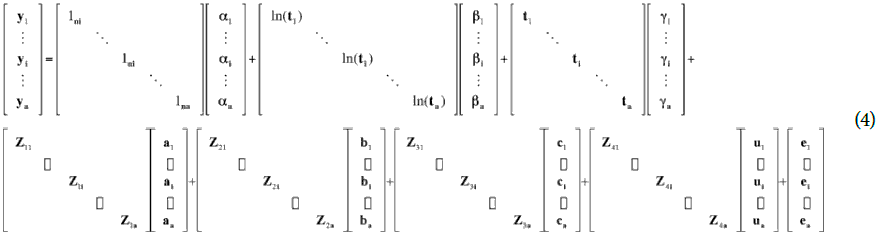
To analyze the data, a multilevel regression model varying regression coefficients (Gelman and Hill, 2007) from the Wood model for each treatment and trial was considered, generating the following mixed model (Sorensen and Gianola, 2002).
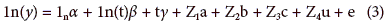
Where a, b and g are the average regression coefficient vectors for each treatment; a, b and c are the vector deviations for each trial of average regression coefficients, u is the vector of coefficients of error due to the effect of blocks within the sampling and trial, e is the vector of residual effects of the error, ln is a vector of each size "n", ln(t) is a vector with the logarithm of the thermal time, t is a vector with the thermal time and Z1, Z2 , Z3 and Z4 are the incidence matrices of the random effects, a, b, c and u respectively. It was assumed that the random effects and the residual error followed a normal distribution with zero mean and variance s2a, s2b, s2c, s2u and s2e (Searle et al., 1992).
Finally, y is the vector with the answer variable to consider, that is, LF, DWT, DWL, DWS, DWR, DWST, DWTB, DWF, DWFR.
Because variable measurements are made on the same individual, a multi-characteristic model was considered:
It was assumed that the vectors a, b, c, u and e, were normally distributed with zero mean and variance Ra 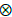 Ia, Rb Ia, Rc Ia, Ru Ia and Rc Ia respectively, with the covariance matrix R(·) expressed in the following form (Eq. 5) (Sorensen and Gianola, 2002).
Ia, Rb Ia, Rc Ia, Ru Ia and Rc Ia respectively, with the covariance matrix R(·) expressed in the following form (Eq. 5) (Sorensen and Gianola, 2002).
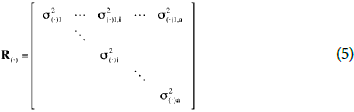
where, I is an Identity of a size, a is the number of variables to evaluate, n indicates the number of data of each variable i, i = {1, ... , a} and is the Kronecker product.
The Bayesian estimation methodology for the previous model was used with the Gibbs algorithm, generating a Markov chain of a length of one million for each parameter. From every 10 samples, 1 was selected and the first 10,000 considered to be the burn-in period were removed. The MCMCglmm package (Hadfield, 2010), of the R statistical environment (R Core Team, 2013), was used. The highest posterior density intervals of 0.90 were calculated with the CODA package (Plummer et al., 2006) and, with the Bayes estimator, the average marginal posterior distribution was obtained, which minimizes the quadratic loss function.
Based on the Markov chain of obtained parameters for the original variables (TTDW, LDW, SDW, RDW, STDW, TBDW, FDW, FRDW), the ratio (PP) of dry matter was obtained in each organ for the evaluated time with the expression:

where, (j) corresponds to LDW, SDW, R DW, STDW, TBDW, FDW, FRDW.
Results and discussion
To establish the existence of significant differences between the curves obtained for the leaf area and dry matter accumulation, a comparison of the highest posterior density intervals was made of the fitted model for each variety in each of the levels of fertilization. In this regard, each graph allowed for the determination of these statistical differences with the curves of the highest posterior density intervals of 0.90.
The organs with the greater amount of dry matter accumulated over the crop cycle are the leaves, stems and tubers; however, the proportion of total plant biomass for each organ changes according to the variety, nutritional conditions and phenological stage; for example, in the vegetative phase, the leaves and stems demand assimilates, which changes to the f lowers and fruits in the reproductive phase. In addition, in crops like the one evaluated in this study, the greatest filling capacity corresponds to the tubers as the starch storage organ and the one of main economic interest.
Criolla Colombia variety
The fertilizing effect over this variety is not noticeable; however, it is possible to perceive differences in the proportion of stem in relation to the total of the plant, which increases with the highest dose. With the 7 g/site dose, in the early stages of the crop cycle, the roots were the ones that represented the greatest amount of accumulated biomass (52.34%) at 255 ADD, a decreasing value over time. This proportion decreased to half at 390.18 ADD (with a rapid decline) and by the end of the cycle at 1,252 ADD the roots represent only 1.98% total of the plant. Stolons, throughout the cycle, represented a fraction of the total plant, reaching the maximum proportion at 643.66 ADD (2.66%), and, at the end of the cycle, the proportion of this organ was less than 1%.
In the early stages of growth at 255 ADD, the leaves and stems were 33.99 and 13.65%, respectively. The highest proportion of leaves to total plant was registered at 508.47ADD (52.71%), but this time did not coincide with the maximum dry matter accumulation in the leaves or the maximum leaf area as recorded above 600 ADD. From that moment of maximum proportion leaves/total plant this begins to decrease, becoming almost 35% at 829.54 ADD and continued to slowly decrease until the end of the cycle, reaching 6.88% of the total. The largest proportion of stems was reached at 694.35 ADD (28.47%) and, as seen in the leaves, after the maximum ratio, it declined until the end, with 10.11% of the total plant (Fig. 1).
Decreased dry matter proportion of the leaves and stems started when the emergence of the tubers at 440.88 ADD (0.61%). This organ rapidly increased its share in the plant, achieving a maximum of 72.32% at 1,252 ADD.
In the corresponding dose of 21 g/site, the behavior of the proportions of the roots, stolons and stems was similar. In the leaves, the maximum rate was reached at 423.98 ADD with a value of 54.77% and, at the end of the cycle, this organ represented 7.56%. The emergence of the first tubers was at 440.88 ADD (0.69%) and, at the end of the cycle, their ratio was 71.32%.
Although the behavior of the ratios of the organs in this study matches Santos (2010), the proportions were lower in their experiment, reporting final proportions for the tubers at 50% of the total plant.
In a study that determined the behavior of the distribution of biomass in varieties of S. tuberosum, Ñústez et al. (2009) found similar results, where the highest proportion of leaves in the Betina variety corresponded to a value close to 40% five weeks after emergence. But values above 80% were reported at the end of the cycle in tubers because the species we evaluated ended their cycle with total plant death, unlike the varieties of this study that ended their cycle alive since they did not have senescence. Likewise, Aguilar et al. (2006) reported a similar behavior in the Alpha and Milagros varieties.
Criolla Guaneña variety
The behavior for the assimilate distribution of the Criolla Guaneña variety did present differences regarding the fertilizer rate. At the beginning of the life cycle of the crop (255 ADD), the organ that occupied the largest proportion was the roots, with values of 52.70 and 50.98% for the doses of 7 and 21 g/site of fertilizer, respectively. The leaves followed the roots with a higher proportion at this time of the cycle, with values of 33.24 and 34.05%, respectively (Fig. 2).
The differences were noticed when the organs, such as the stems, reached their maximum proportion. For the low fertilizer dose, the stems reached their greatest proportion at 626.76 ADD, corresponding to 25.96%, while, in the highest dose, the maximum proportion (31.65%) was achieved 84.49 ADD later and this proportion was maintained longer. 152 ADD later, the proportion began to decrease until it reached 11.05% at 1,252 ADD and, in the low dose, at the end of the cycle the stems correspond to 25% of the plant.
The leaves achieved the maximum proportion at 491.57 ADD with both fertilizer doses, 49.75 and 54.05% for the lowest and highest doses, respectively; at the end of the cycle at 1,252 ADD with the lower dose, the leaves occupied 19.73% and, with the higher dose, they occupied 7.41%. The most obvious difference was observed in the distribution of biomass in the tubers. With the 7 g/site dose, this organ emerged at 440.88 ADD at a rate of 1.09%. With the 21 g/site dose, the tubers emergence was recorded at 508.47 ADD at 1.36% of the plant. The increase in the proportion of this organ was a constant for the higher dose of fertilizer, which did not stop its progress and which corresponded to 74.38% of the plant at 1,252 ADD. At low doses, an increase in the proportion of the tubers went up to 40.81% at 1,066.11 ADD; this ratio dropped until the end of the cycle where it registered a value of 36.81%.
In the low dose of fertilizer in this variety, the significant increase in the proportion of fruit in the final stage of the crop, which went from 0.42% (at 812.64 ADD) to 12.98% (at the end of the cycle), was noticable. This led to the conclusion that, with the lower fertilizer dose, tubers in the Guaneña variety had to lose some of the filling capacity, leading to the sexual reproduction organs; in this case, the fruits have won this important total percentage of the plant.
For the same variety, Santos (2010), although not as clear as in this study, also reported an increase in the proportion of fruit and a stability in the increase of the proportion of tubers in the plant. The distribution of biomass in terms of other organs also differed since the author reported the highest proportion of leaves in the initial phase, with a 50% and continuously decreases until the end and the maximum proportion of tubers was recorded at the end of cycle, 77 d after sowing with a value of 50% as well.
Ñústez et al. (2009), for the Pastusa Suprema variety, reported a similar behavior where the leaves and stems gradually increased their share until the emergence of the tubers when they started to decrease.
Criolla Latina variety
This variety has a similar behavior to the Guaneña variety at low and high doses of fertilizer.
Starting the cycle, at 255 ADD in the low dose of fertilizer the organ representing the largest proportion of the plant is the root (58.83%) followed by leaves and stems with values of 30.07 and 10.48% respectively. Both the leaves and stems increased their proportion to reach a maximum and then decreased until the end of the cycle. The largest proportion of leaves, 57.16%, occurred at 423.98 ADD and the maximum proportion of stems, 25.46%, was seen at 694ADD. The decline in the proportion of these organs was constant until 1,252 ADD, with values of 10.27% in the leaves and 14.32% in the stems (Fig. 3).
In the high dose of fertilizer, at 255ADD, the organ that had the biggest proportion was the leaves (44.00%), followed by the roots (42.74%) and then the stems with a ratio of 12.77%. The maximum proportion of leaves was reached at 423.98 ADD, with a value of 57.16%, which dropped until 1,252 ADD, finishing with 4.90%. The stems reached the maximum proportion at 694.35 ADD with a 29.93% value and ended the cycle with 7.46% of the total plant.
Similar to the Guaneña variety, differences in the distribution of biomass in the tubers can be seen when comparing the fertilizer dose. The emergence of the organ was recorded in both doses at 440.88 ADD, with a proportion of 1.56% with the 7 g/site dose and 0.21% with the 21 g/ site dose. At the high dose, the behavior of the distribution of the dry matter to the tubers progressively increased the proportion of this organ until the end of the cycle, with a value of 83.13% (increase that occurs until 1,184 ADD where it stabilizes). In this case, the fruits also showed an increase in the proportion, ending the cycle with a value of 13.57%.
According to the results of Santos (2010), a higher proportion of tubers was obtained at the end of the cycle. This author reported a maximum proportion 77 d after sowing, with a value of 60%. However, in the behavior and progress of the distribution over time, these results agree with the findings of Aguilar et al. (2006), Mora et al. (2005), and Ñústez et al. (2009) for each of the varieties of the S. tuberosum species.
In general terms, the results of this study agree with Meena et al. (2014), where the maximum tuber formation and tuber bulking rate were recorded from 60 to 90 d after crop sowing, during the large growth phase. This occurred because of the availability of nutrients for crop growth and development, as well as for partitioning of the dry matter production. Similarly, the results of Silva et al. (2013) showed that, in the Bintje variety, tuber filling starts 60 d after sowing and the volume of the tubers is greater when applying more calcium than nitrogen, while in the Atlantic variety, the largest volume of tubers occurs when there is more nitrogen than calcium.
Conclusion
The expected results according to literature were obtained. In the vegetative phase of the plants, photosynthates are used for the formation of new stems and leaves and, in the reproductive phase, they tend to accumulate dry matter in the f lowers and fruits; in the particular case of this species, the reproductive organs had a lower filling capacity than the tubers, which are the organs used for the storage of starch.
Literature cited
Aguilar, M.G., J. Carrillo, A. Rivera, and V. González. 2006. Growth analysis and sink-source relationships in two potato (Solanum tuberosum L.) varieties. Rev. Fitotec. Mex. 29, 145-156. [ Links ]
Fourcaud, T., X. Zhang, A. Stokes, H. Lambers, and C. Körner. 2008. Plant growth modelling and applications: the increasing importance of plant architecture in growth models. Ann. Bot. 101, 1053-1063. Doi: 10.1093/aob/mcn050 [ Links ]
Foyer, C. and M. Paul. 2001. Source - Sink relationships. pp. 1-11. In: Encyclopedia of life sciences. Nature Publishing Group, London. Doi: 10.1038/npg.els.0001304 [ Links ]
Gelman, A. and J. Hill. 2007. Data analysis using regression and multilevel/hierarchical models. Cambridge University Press, New York, NY. [ Links ]
Gutierrez, A.P. 1996. Applied population ecology: a supply-demand approach. Wiley, New York, NY. [ Links ]
Hadfield, J.D. 2010. MCMC methods for multi-response generalized linear mixed models: the MCMC glmm R package. J. Stat. Softw. 33, 1-22. Doi: 10.18637/jss.v033.i02 [ Links ]
Holdridge, L. 1967. Life zone ecology. Tropical Life Science, San Jose. [ Links ]
Kooman, P.L. and R. Rabbinge. 1996. An analysis of the relation between dry matter allocation to the tuber and earliness of a potato crop. Ann. Bot. 77, 235-242. Doi: 10.1006/anbo.1996.0027 [ Links ]
Larcher, W. 2003. Physiological plant ecology. Ecophysiology and stress physiology of the functional groups. 4th ed. Annals of Botany.Springer, Berlin. Doi: 10.1007/978-3-662-05214-3 [ Links ]
Manetsch, T.J. 1976. Time-varying distributed delays and their use in aggregative models of large systems. IEEE Trans. Syst., Man, Cybern. SMC-6, 547-553. Doi: 10.1109/TSMC.1976.4309549 [ Links ]
Meena, B.P., A. Kumar, M.L. Dotaniya, N.K. Jat, and B. La l. 2014. Effect of organic sources of nutrients on tuber bulking rate, grades and specific gravity of potato tubers. Proc. Natl. Acad. Sci., India, Sect. B Biol. Sci., 1-7. Doi: 10.1007/s40011-014-0398-4 [ Links ]
Mora, R., J. Ortiz, A. Rivera, M. Colinas, H. Lozoya, and J. Rodríguez. 2005. Comportamiento de la acumulación y distribución de biomasa en genotipos de papa establecidos en condiciones de secano. Rev. Chapingo Ser. Hortic. 11, 135-142. [ Links ]
Ñústez, C., M. Santos, and M. Segura. 2009. Dry matter allocation and partitioning of four potato varieties (Solanum tuberosum L.) in Zipaquirá, Cundinamarca (Colombia). Rev. Fac. Nal. Agr. Medellin 62, 4823-4834. [ Links ]
Plummer, M., N. Best, K. Cowles, and K. Vines. 2006. CODA: convergence diagnosis and output analysis for MCMC. R News 6, 7-11. [ Links ]
R Core Team. 2013. A language and environment for statistical computing. www.r-project.org [ Links ]
Rodríguez, D., J.R. Cure, J.M. Cotes, A.P. Gutierrez, and F. Cantor. 2011. A coffee agroecosystem model: I. Growth and development of the coffee plant. Ecol. Model. 222, 3626-3639. Doi: 10.1016/j.ecolmodel.2011.08.003 [ Links ]
Santos, M. 2010. Evaluación del crecimiento, desarrollo y componentes de rendimiento de cuatro cultivares de papa criolla en dos loca lidades del depar ta mento de Cundina marca. MSc thesis. Faculty of Agronomy, Universidad Nacional de Colombia, Bogota. [ Links ]
S earle, S., G. Casella, and C.E. McCulloch. 1992. Variance components. Jhon Wiley and Sons, New York, N Y. Doi: 10.1002/9780470316856 [ Links ]
Silva, J.G., M.G.C. França, F.T.F. Gomide, and J.R. Magalhaes. 2013. Different nitrogen sources affect biomass partitioning and quality of potato production in a hydroponic system. Amer. J. Potato Res. 90, 179-185. 10.1007/s12230-012-9297-5 [ Links ]
Sorensen, D. and D. Gianola. 2002. Likelihood, bayesian, and MCMC methods in quantitative genetics (statistics for biology and health). Springer, Madison, MS. Doi: 10.1007/b98952 [ Links ]
Taiz, L. and E. Zeiger. 2010. Plant physiology. 5th ed. Sinauer Associates, Sunderland, MA. [ Links ]
Tekalign, T. and P.S. Hammes. 2005a. Growth and productivity of potato as influenced by cultivar and reproductive growth. I. Stomatal conductance, rate of transpiration, net photosynthesis, and dry matter production and allocation. Sci. Hortic. 105, 13-27. Doi: 10.1016/j.scienta.2005.01.029 [ Links ]
Tekalign, T. and P.S. Hammes. 2005b. Growth and productivity of potato as influenced by cultivar and reproductive growth. II. Growth analysis, tuber yield and quality. Sci. Hortic. 105, 29-44. Doi: 10.1016/j.scienta.2005.01.021 [ Links ]
Van Heemst, H.D.J. 1986. The distribution of dry matter during growth of a potato crop. Potato Res. 29, 55-66. Doi: 10.1007/BF02361981 [ Links ]
Vansick le, J. 1977. Attrition in distributed delay models. IEEE Trans. Syst., Man, Cybern. SMC-7, 635-638. Doi: 10.1109/TSMC.1977.4309800 [ Links ]
Warren, J. 1981. Analysis of growth, photosynthesis and light interception for single plants and stands. Ann. Bot. 48, 507-512. [ Links ]
Wood, P. 1967. Algebraic model of the lactation curve in cattle. Nature 216, 164-165. Doi: 10.1038/216164a0 [ Links ]


{kind=link}