











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introduction
The Americas contribute 8.8% of the global burden of neglected infectious diseases (NIDs)1, An estimated 6-8 million people are infected with Trypanosoma cruzi (T. cruzi), the parasitic protozoan that causes Chagas disease (CD) (also known as American Trypanosomiasis), with 65-100 million people at risk worldwide. There are 30,000 new cases annually due to vector transmission and another 8,000 due to congenital transmission, causing approximately 14,000 deaths per year 1-5. The disease is endemic in 21 countries in Latin America and the Caribbean. An upward trend in the number of reported cases has been observed in Canada, Asia, the United States, and Europe, attributed mainly to migration and poverty 1,4,6,7. In 2021 poverty in Latin America affected more than 184 million people (32.3 % of the population) with another 12.9 % living in extreme poverty; these conditions remain higher than before the COVID-19 pandemic. In rural areas poverty and extreme poverty reach 44.1 % and 21.2 %, respectively, mainly affecting children and adolescents up to 17 years of age, women of productive age, people with low education, indigenous people, Afro-descendants and people with disabilities 8. Migration, access to travel, and expansion of international markets, especially to the United States and Europe, combined with poverty, limited access to health systems and deficiencies in housing, water and basic sanitation for migrants, have increased the risk of transmission of infectious diseases in non-endemic countries 1,6,9-11.
The success in the spread of the disease and the difficulties in its control and elimination can be attributed to several factors: the genetic diversity of T. cruzi (classified into seven different discrete typing units or DTUs: Tcl-TcVI and Tcbat 12) and the absence of rapid and specific tests for different genotypes, the parasite's capacity to infect various species of vertebrates and vectors (>100 and >150, respectively), the lack of knowledge about the prevalence and incidence of chronic disease, the low specificity of clinical symptoms, the absence of highly effective vaccines and drugs, the toxicity of existing treatments and poor adherence to them, the high costs that limit access to treatment and the resistance of vectors to insecticides used for their control 3,13-15
Chagas disease progresses through three phases: acute, indeterminate and chronic. The acute phase is characterized by mild, intermittent symptoms and abundant presence of parasitic forms in the bloodstream; followed by an indeterminate and a chronic phases in which the parasites invade nucleated muscle cells potentially leading to coronary (20 - 30 % of cases) or gastrointestinal (10 - 20 % of cases) disease, with the presence of megacolon or megaesophagus. During the acute phase the diagnosis is established mainly by the detection of trypomastigotes in blood, while in the chronic phase, serological tests for the determination of immunoglobulin G (IgG) of specific antibodies that bind to T. cruzi antigens (enzyme-linked immunosorbent assay (ELISA) and immunofluorescence assay (IFA)) are useful.
Nifurtimox and benznidazole, introduced in the l960s, remain the only drugs available for CD treatment. Pharmacological treatment should be initiated in the acute phase of the disease since its effectiveness decreases drastically and is even considered controversial when the chronic phase is reached. In addition, the safety profile of these drugs raises significant concerns due to frequent adverse effects, ranging from digestive disorders and psychic alterations to more severe conditions such as bone marrow depression, peripheral polyneuropathy, lymphadenopathy, thrombocytopenic purpura, and agranulocytosis. Pharmaceutical companies have shown very limited interest in developing new therapies against trypanosomatid parasites, due to the low expected return on investment as the target communities often have little to no purchasing power 3,4,6,16,17.
Recently, a meta-analysis that aimed to establish the in vitro susceptibility of the Y strain to benznidazole showed a wide variability in the IC50 values obtained, without clearly determining the cause of this heterogeneity, leading the authors to conclude that variability could be explained by the lack of uniformity in the experimental procedures for trypanocidal activity tests 18. Two in vitro studies using TcI, TcII and TcV DTUs failed to establish a correlation between benznidazole susceptibility and genetic distances among these DTU strains 19,20, additionally, a systematic review was unable to find statistically significant associations between T. cruzi genotype and clinical outcome 31. These studies employed different sources of parasite stages (e.g., trypomastigotes forms from infected mouse bloodstream or cell culture, amastigotes from different host cell lines, epimastigotes), various culture media, different benznidazole sources (pure active compound or commercial pills), and a variety of methods to determine parasite viability. This variability underscores the critical need for establishing standardized protocols in future research to ensure more consistent and comparable results across studies.
In vitro pharmacological models play a pivotal role as an initial filter in identifying substances with antiparasitic activity against T cruzi, being a fast, simple and inexpensive tool; these models employ a wide variety of cell lines, parasite strains and culture conditions, allowing us to expand our knowledge about the biology, mechanisms of parasitic infection and have been key to the discovery of molecules with antiparasitic activity; however, despite the advantages, the lack of consensus on in vitro screening protocols has emerged as a significant obstacle in the discovery of new drugs to treat Chagas disease.
The absence of well-defined experimental conditions for evaluating promising substances has resulted in poor reproducibility and reliability of results. This inconsistency becomes an obstacle when attempting to compare findings across different studies as a foundation for subsequent stages of the pharmacological research and development (R&D) process. Therefore it is not surprising that despite a large number of compounds with promising activity, in in vitro tests, are frequently reported, only four substances -allopurinol, itraconazole, posaconazole and fluconazole- have advanced to clinical studies 3,22,23; unfortunately, in clinical trials, none of these compounds have demonstrated superior efficacy to benznidazole, particularly when evaluated in patients with chronic CD. The drug R&D processes for Chagas disease are poor leaving an urgent need for new effective treatments, necessary to meet the World Health Organization (WHO)'s roadmap for neglected tropical diseases and to achieve the United Nations Sustainable Development Goals 4,11. The Open Science movement aims to promote research integrity, experimental and analytical reproducibility and full transparency. A major challenge in the research and development of drugs against trypanosomatids remains the lack of defined conditions/standards for the selection and validation of promising compounds. This deficiency often leads to conflicting results between different research teams 3.
Pharmacological models for trypanosome research have recently garnered growing interest, reflected in numerous reviews addressing various aspects such as: advances in Trypanosomatid biology from a drug R&D perspective and the identification of promising molecular targets 7,11, procedural approaches for assessing trypanocidal activity and toxicity in mammalian cells. Additionally, they explore infection characteristics and methodologies 15,22, the evolution of Chagas disease drug discovery techniques and current pharmacological models used in drug R&D cascades 3,24, medicinal chemistry, chemotherapy and therapeutic options in development 4,6,11. To address the challenge posed by Bhattacharya 3 and provide a comprehensive picture of in vitro antitrypanosomal screening conditions used in research and development of new therapeutic alternatives against CD, this scoping review aimed to identify and present available information regarding experimental conditions of epimastigotes models. Additionally, we aimed to propose criteria and conditions for homogenizing experimental protocols (Box 1). Through this approach, we hope to contribute to ensuring consistency and comparability of results across different laboratories.
Methodology
Search strategy
The review covered PubMed, Scopus and LILACS databases, focused on articles published between the years 2008 and July 2023, a time frame chosen based on preliminary analysis of publication trends. A preliminary search in PubMed for "in vitro antitrypanosomal screening" revelated low and intermittent scientific production (<4 articles / year) between l989 and 2007, which markedly increase from 2008 onwards (>4 articles / year). The search equation included the following terms MeSh (for English) and DeCS (for Spanish and Portuguese) ("Trypanosoma cruzi", "Parasitic Sensitivity Tests" and "Trypanocidal Agents"), in all cases the Boolean operator "AND" was used. To capture relevant articles potentially missed in database searches, reference lists of key sources were examined. Article review and data collection were conducted independently by at least two reviewers. Data were subsequently cross-checked, and discrepancies were resolved through consensus meetings. The study followed current, commonly accepted requirements and guidelines for reporting systematic reviews 25-27.
Study/source of evidence selection
This scoping review used the following inclusion criteria to select articles: English, Spanish or Portuguese language, studies reporting the conduct of in vitro antiparasitic activity against T. cruzi epimastigotes. Articles not specifying strain and parasitic form used and those that correspond to different parasite stages (e.g., trypomastigotes, amastigotes), immunological studies, genetic characterization studies of T. cruzi strains, biological studies, case reports, and non-primary sources were excluded. Article selection involved screening titles, abstracts, and keywords, followed by full-text review when necessary. This scoping review aimed to describe and map the literature to provide a comprehensive picture of in vitro antitrypanosomal screening conditions and to propose risk of bias assessment criteria for such studies.
Data extraction
A comprehensive database was constructed to record information in two main categories: regarding the pharmacological model, T. cruzi strain or isolate, culture media, serum supplementation (type and percentage), culture media enrichment, culture media replacement frequency (in days) and incubation temperature. From the antiparasitic pharmacological test information was extracted regarding infective inoculum (Parasites/mL), treatment exposure time (hours (h)), type of well culture plate and methodology to quantify parasite viability; finally, the active control used (reference drug), concentration tested, treatment duration and antiparasitic activity results were reviewed.
Results
Literature search
The Bibliographic search across all consulted databases, yielded a total of 392 records. After manually removing 189 duplicates and excluding l unavailable article, 202 articles remained for review (Figure 1). The analysis of database overlap revealed a concordance rate of 1.5 % between PubMed-Scopus-LILACS, 47.1 % between PubMed-Scopus, 4.1 % between PubMed-LILACS and 2.9 % between Scopus-LILACS. The remaining 202 articles were reviewed for title, abstract and keywords, of these 139 were discarded due to predetermined exclusion criteria. The remaining articles underwent full-text review by at least two independent readers yielding a final selection of 63 articles, encompassing 105 assays that met all inclusion criteria. The complete article selection process is illustrated in Figure 1.
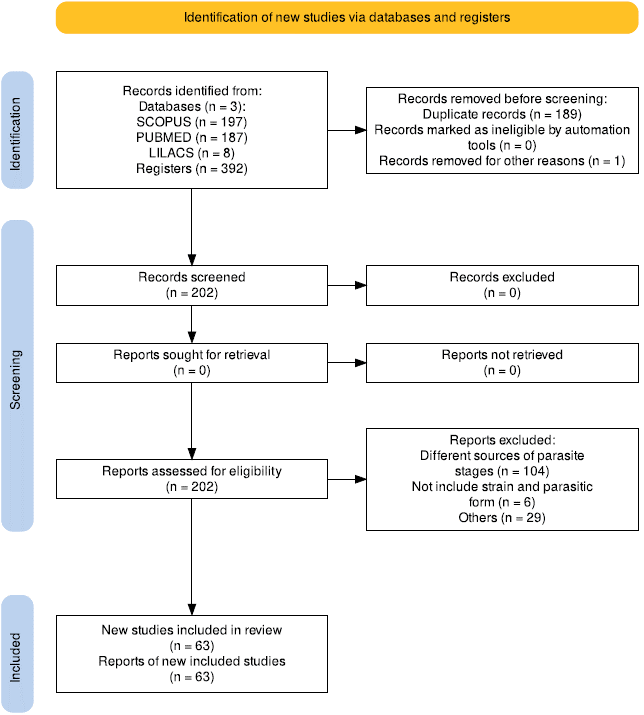
A global analysis of the parasitic forms used in the reports of in vitro trypanocidal activity (data not show). Out of 167 articles reviewed, the most frequently used parasitic forms were: amastigotes (104 reports), epimastigotes (63 reports), blood trypomastigotes (42 reports) and cell culture derived trypomastigotes (49 reports), Figure 2. The use of a single parasitic form was reported in 96 articles (57 %), 52 articles (31 %) reported the use of 2 parasitic forms, and in 19 articles (11 %) 3 parasitic forms were used.
Due to the variability that arises from analyzing different sources of parasite stages simultaneously, such as trypomastigote forms from the bloodstream of infected mice or cell cultures, amastigotes from various host cell lines, and epimastigotes; this scoping review focuses specifically on pharmacological models that assess antitrypanosomal activity in epimastigotes.
Reported conditions in pharmacological model of in vitro culture of T. cruzi epimastigotes
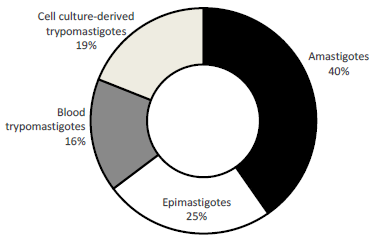
Antiparasitic activity against epimastigote forms was evaluated in 63 reports (106 assays) that informed the use of 28 different strains. Strain Y (Tc II) was the most frequently informed (33 assays), followed by strains CL (Tc VI) (15 assays), Tulahuen (Tc VI) (9 assays) and DM28c (Tc I) (8 assays), four strains were tested in 3-5 assays (SN3(IRHOD/CO/2008/SN3), Silvio X10, RA (Tc VI), Arequipa) and 20 others 1-2 assays (Esmeraldo cl3, Colombiana (Tc I), Maracay, YuYu (Tc I), Berenice 62, VL10, DA (MHOM/CO/01/07), MHOM/ES/2203/BCN590, M6241 cl6, 793, 893, 4166, NR cl3 (Tc V), SO3 cl5 (Tc V), 115, 294, 597, INC-5, NINOA, TH).
The use of liver infusion-tryptose culture medium (LIT) predominated with 58 assays (of which 14 assays correspond to Franco, J., et al.29 and 8 to Zingales, B., et al.30), followed by brain heart infusion (BHI), Roswell Park Memorial Institute (RPMI) 1640 media, Medium Trypanosomes Liquid (MTL) and Biphasic 3N medium (20, 7, 7 and 4 assays each, respectively), and the other media with two or one report (Diamond's monophasic medium, BHT, 3N + MEM, Grace's Insect Medium). Regarding the culture media enrichment with fetal bovine serum (FBS) predominated a concentration of 10% (79 tests of which 14 assays correspond to Franco, J., et al.29 and 8 to Zingales, B., et al.30), followed by FBS (5%) in 14 tests and FBS (20%) in three tests, 10 assays did not inform the FBS concentration used, Figure 3.
The culture media replacement frequency was not frequently reported (79 assays), in the remaining ones, replacement every 7d (9 assays) followed by every 3-4d (8 assays) and every 10d (7 assays) predominated; replacement every 5d and 8d was reported one and two times each respectively. Finally, the most reported incubation temperature, in parasite culture and in pharmacological tests was 28 °C (91 assays) followed by 26 °C (6 assays) and 27 °C (3 assays), 6 assays did not report the temperature used.
Strains with 1-2 assays (each): Esmeraldo cl3, Colombiana (Tc I), Maracay, YuYu (Tc I), Berenice 62, VL10, DA (MHOM/ CO/01/07), MHOM/ES/2203/BCN590, M6241 cl6, 793, 893, 4166, NR cl3 (Tc V), SO3 cl5 (Tc V), 115, 294, 597, INC-5, NINOA, TH. Strains with 3-5 assays (each): SN3(IRHOD/CO/2008/SN3), Silvio X10, RA (Tc VI), Arequipa. Culture media with one report (each): 3N + MEM and Grace's Insect Medium. Culture media with two reports (each): BHT and Diamond's monophasic.
* 14 assays correspond to Franco, J., et al.29. ** 8 assays correspond to Zingales, B., et al.30.
Not reported (NR).
Figure 3: Pharmacological model of epimastigote culture: strain, culture media, enrichment serum percentage and incubation temperature, most frequently used to obtain parasite forms.
Pharmacological test conditions for T. cruzi epimastigotes
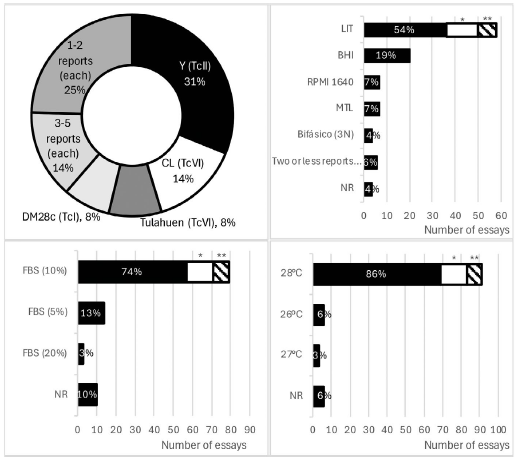
The initial infecting inoculum (parasites/mL) varies across three orders of magnitude, with a minimum value of 1 x 105 parasites/mL and a maximum value of 1 x 107 parasites/mL (100 times higher). This parameter is crucial for harmonizing test conditions, as it directly relates to the challenge faced by the substance being evaluated. A low initial inoculum may incorrectly declare inactive substances as active, while an excessively high infecting inoculum could lead to identify active substances as inactive. The latter scenario is known as a type II error, or false negative. Therefore, careful consideration of the initial inoculum is essential for accurate and reliable test results in parasitology studies.
Two outlier reports, involving 14 and 8 assays each 29,30 significantly distort the overall trend and are highlighted in Figure 4. Excluding these outliers, 97 % of the reports present 3 or fewer tests each. Among these reports, the initial infectious inoculum of 1 x 106 parasites/mL predominates in 27 % of the tests. This is followed by 5 x 105 parasites/mL (10 % of tests) 2 x 106 and 5 x 106 parasites/mL (7 % of tests each), and 1.5 x 106 parasites/mL (6 % of tests). Other infecting inoculum concentrations are reported in 5% or fewer of the cases. Notably, 13% of the tests reported parasite concentration as parasites/well without specifying the volume used in the well, making direct comparisons difficult. Additionally, 7 % of the test did not report units of parasite concentration, and one report failed to provide this important parameter altogether. When the two outlier reports 29,30 are included in the analysis, the proportion of test using I x 107 parasites/mL increase dramatically from 5 % to 25 % of all tests. This shift underscores the significant impact these outliers have on the overall distribution, Figure 4.
The addition of antibiotics in some test is concerning penicillin (5 tests), streptomycin (6 tests), penicillin-streptomycin (9 tests), and gentamicin (4 tests). This practice has been questioned in other pharmacological models, as good laboratory practices should ensure non-contamination of parasite cultures without antibiotic use. Regarding cell counting methods, microscopy with a Neubauer chamber was the predominant method, used in 58 of the reports, only I used microscopy with an Automated cell counter (TC20). Various other techniques were employed to determine parasite viability: colorimetric tests were described in 22 tests (MTT II, CPRG 3, Resazurin 8). Turbidimetry (referred to as absorbance) was informed in 16 tests. Six of the reports mentioned the use of fluorometry (Resazurin 5 and Propidium iodide I). Less frequently, radiometry using [3H]-thymidine was employed (3 reports), Figure 4.
The most frequently reported active control was Benznidazole, used in 70 tests, followed by nifurtimox in 13 tests. Two active controls were used in some tests: Benznidazole + Nifurtimox in 7 tests and Nifurtimox + Miltefosine in I test. However, there are notable omissions in the reporting of controls: 15 reports did not mention the use of any control drug, and 12 reports indicated the use of active control but failed to provide the resulting parasite inhibition data. These omissions are concerning, as the use of active control is essential for demonstrating adecuate experimental design and providing evidence of proper bias control in experimental pharmacology, Figure 4.
* 14 assays correspond to Franco, J., et al.29.
** 8 assays correspond to Zingales, B., et al.30.
***Neubauer (58), Automated cell counter (TC20) (1).
****MTT (II), Resazurin (8), Chlorophenol red-ß-d-galactopyranoside (CPRG) (3).
***** Resazurin (5), Propidium ioide (1).
Not reported (NR).
Figure 4: Epimastigotes screening conditions of in vitro trypanocidal tests: Infective inoculum (Parasites/mL), Active control, Treatment exposure time (Hours (h)) and Methodology to quantify parasite viability.
The inhibition of parasite multiplication achieved by the reference drug is reported in various terms: Inhibitory Concentration 50 (IC50) in 68 % of tests, fifty percent of Parasite Growth Inhibition (PG150) in 4 % of tests, and minorly so as Cytotoxic Concentration 50 (CC50) in 1 % of tests, Percentage of Parasite Death in 1 %, Inhibitory Dose 50 (ID50) in 1 %, fortunately, these last confusing interpretations of terms are rare. However, it is worth noting that 25 % of tests do not report the parasite inhibition generated by the reference drug. Despite the lack of uniformity in the experimental procedures for trypanocidal activity tests,
Table 1 Summarizes the pharmacological activity data reported for the active controls Benznidazole, Nifurtimox and Miltefosine.
| Reference | Author, Year | T. cruzi strain (DTU) | Active control | Treatment exposure time (h) | Results (μM) |
|---|---|---|---|---|---|
| 38 | Gusmão et al., 201 | Y (TcII) | Benznidazole | 72 | 30.78 |
| 42 | Salsi et al., 2019 | Y (TcII) | Benznidazole | 72 | 15.1 |
| 43 | Fonseca-Berzal et al., 2018 | Y (TcII) | Benznidazole | 48 | 28.11 |
| 24 | 218 | ||||
| 44 | Lima et al., 2018 | Y (TcII) | Benznidazole | 48 | 61.1 |
| 72 | 16.5 | ||||
| 45 | Cardoso et al., 2014 | Y (TcII) | Benznidazole | 264 | 6.6 |
| 47 | Velásquez et al., 2014 | Y (TcII) | Benznidazole | 72 | 34 |
| 48 | Franklim et al., 2013 | Y (TcII) | Benznidazole | 168 | 2.2 |
| 49 | Izumi et al., 2013 | Y (TcII) | Benznidazole | 96 | 6.53 |
| 50 | Jorge et al., 2013 | Y (TcII) | Benznidazole | 120 | 22.69 |
| 51 | Moreira et al., 2013 | Y (TcII) | Benznidazole | 72 | 34.5 |
| 52 | Carneiro et al., 2012 | Y (TcII) | Benznidazole | 72 | 11.5 |
| 53 | Izumi et al., 2012 | Y (TcII) | Benznidazole | 96 | 6.5 |
| 54 | Daliry et al., 2011 | Y (TcII) | Benznidazole | 24 | 168 |
| 56 | Veiga-Santos et al., 2010 | Y (TcII) | Benznidazole | 96 | 7.8 |
| 57 | Dos Santo Filho et al., 2009 | Y (TcII) | Benznidazole | 264 | 6.6 |
| 58 | Batista et al., 2008 | Y (TcII) | Benznidazole | 72 | 10.3 |
| 7 | Fonseca-Berzal et al., 2018 | CL (TcVI) | Benznidazole | 72 | 26.55 |
| 29 | Franco et al., 2015 | CL (TcVI) | Benznidazole | 72 | 13.2 |
| 30 | Zingales et al., 2015 | CL (TcVI) | Benznidazole | 72 | 13.2 |
| 67 | Vega et al., 2012 | CL (TcVI) | Benznidazole | 72 | 27.32 |
| 78 | Cardoso et al., 2019 | DM28c (TcI) | Benznidazole | 120 | 6.6 |
| 79 | Ceole et al., 2018 | DM28c (TcI) | Benznidazole | 24 | 59 |
| 81 | Da Silva et al., 2017 | DM28c (TcI) | Benznidazole | 96 | 48.8 |
| 83 | Gomes et al., 2016 | DM28c (TcI) | Benznidazole | 264 | 48.8 |
| 84 | Espíndola et al., 2015 | DM28c (TcI) | Benznidazole | 120 | 48.8 |
| 72 | Paucar et al., 2019 | Tulahuen (TcVI) | Benznidazole | 48 | 25.3 |
| 42 | Salsi et al., 2019 | Tulahuen (TcVI) | Benznidazole | 72 | 17.9 |
| 75 | Martín-Escolano et al., 2018 | Tulahuen (TcVI) | Benznidazole | 48 | 19.7 |
| 76 | Moreno-Viguri et al., 2016 | Tulahuen (TcVI) | Benznidazole | 72 | 18.7 |
| 55 | Álvarez et al., 2010 | Tulahuen (TcVI) | Benznidazole | 120 | 7.4 |
| 59 | Boiani et al., 2008 | Tulahuen (TcVI) | Benznidazole | 120 | 4.5 |
| 29 | Franco et al., 2015 | Silvio X10 (TcI) | Benznidazole | 72 | 26.1 |
| 30 | Zingales et al., 2015 | Silvio X10 (TcI) | Benznidazole | 72 | 26.1 |
| 30 | Zingales et al., 2015 | Esmeraldo cl3 (TcII) | Benznidazole | 72 | 26.7 |
| 32 | Muscia et al., 2020 | RA (TcVI) | Benznidazole | 72 | 5.8 |
| 31 | Beer et al., 2016 | RA (TcVI) | Benznidazole | 72 | 4.99 |
| 90 | Martín-Montes et al., 2021 | SN3(IRHOD/CO/2008/ SN3) (TcI) | Benznidazole | 72 | 15.8 |
| 72 | Paucar et al., 2019 | SN3(IRHOD/CO/2008/ SN3) (TcI) | Benznidazole | 48 | 15.3 |
| 75 | Martín-Escolano et al., 2018 | SN3(IRHOD/CO/2008/ SN3) (TcI) | Benznidazole | 48 | 36.2 |
| 76 | Moreno-Viguri et al., 2016 | SN3(IRHOD/CO/2008/ SN3) (TcI) | Benznidazole | 72 | 15.8 |
| 85 | Ramírez-Macías et al., 2012 | SN3(IRHOD/CO/2008/ SN3) (TcI) | Benznidazole | 72 | 15.89 |
| 29 | Franco et al., 2015 | Colombiana (TcI) | Benznidazole | 72 | 40.5 |
| 30 | Zingales et al., 2015 | Colombiana (TcI) | Benznidazole | 72 | 34.1 |
| 92 | Marín et al., 2011 | Maracay (TcVI) | Benznidazole | 72 | 15.8 |
| 91 | Maldonado et al., 2010 | Maracay (TcVI) | Benznidazole | 72 | 16 |
| 86 | Hamedt et al., 2014 | DA (MHOM/CO/0I/07) (TcI) | Benznidazole | 72 | 26.9 |
| 87 | Di Pietro et al., 2015 | MHOM/ES/2203/ BCN590 (TcI) | Benznidazole | 72 | 36.2 |
| 29 | Franco et al., 2015 | YuYu (TcI) | Benznidazole | 72 | 40.5 |
| 30 | Zingales et al., 2015 | YuYu (TcI) | Benznidazole | 72 | 40.5 |
| 29 | Franco et al., 2015 | Berenice 62 (TcII) | Benznidazole | 72 | 14.6 |
| 30 | Zingales et al., 2015 | Berenice 62 (TcII) | Benznidazole | 72 | 14.6 |
| 29 | Franco et al., 2015 | 793 (Tcbat) | Benznidazole | 72 | 16.6 |
| 29 | Franco et al., 2015 | 893 (TcIII) | Benznidazole | 72 | 74.9 |
| 29 | Franco et al., 2015 | SO3 cl5 (TcV) | Benznidazole | 72 | 10.6 |
| 30 | Zingales et al., 2015 | 115 (TcV) | Benznidazole | 72 | 7.6 |
| 29 | Franco et al., 2015 | VL10 (TcII) | Benznidazole | 72 | 30.4 |
| 30 | Zingales et al., 2015 | VL10 (TcII) | Benznidazole | 72 | 30.4 |
| 88 | Chacón-Vargas et al., 2017 | INC-5 (TcI) | Benznidazole | 24 | 42.34 |
| 89 | Domínguez-Díaz et al., 2021 | NINOA (TcI) | Benznidazole | 48 | 30.27 |
| 89 | Domínguez-Díaz et al., 2021 | TH (ITRI/MX/2018/TH) (TcI) | Benznidazole | 48 | 39.08 |
| 72 | Paucar et al., 2019 | Arequipa (TcV) | Benznidazole | 48 | 26.2 |
| 76 | Moreno-Viguri et al., 2016 | Arequipa (TcV) | Benznidazole | 72 | 15.8 |
| 75 | Martín-Escolano et al., 2018 | Arequipa (TcV) | Benznidazole | 48 | 16.9 |
| 41 | Espinosa-Bustos et al., 2020 | Y (TcII) | Nifurtimox | 120 | 7 |
| 46 | Tapia et al., 2014 | Y (TcII) | Nifurtimox | 120 | 7 |
| 50 | Jorge et al., 2013 | Y (TcII) | Nifurtimox | 120 | 3.78 |
| 59 | Boiani et al., 2008 | Y (TcII) | Nifurtimox | 120 | 9.7 |
| 62 | Scalese et al., 2021 | CL (TcVI) | Nifurtimox | 24 | 3.68 |
| 64 | Scalese et al., 2018 | CL (TcVI) | Nifurtimox | 120 | 2.8 |
| 66 | Santos et al., 2012 | CL (TcVI) | Nifurtimox | 72 | 3.16 |
| 67 | Vega et al., 2012 | CL (TcVI) | Nifurtimox | 72 | 3.6I |
| 69 | Rodríguez et al., 2009 | CL (TcVI) | Nifurtimox | 120 | 3.4 |
| 59 | Boiani et al., 2008 | CL (TcVI) | Nifurtimox | 120 | 4.9 |
| 77 | López-Lira et al., 2021 | DM28c (TcI) | Nifurtimox | 24 | 21.05 |
| 82 | Robledo-O'Ryan et al., 2017 | DM28c (TcI) | Nifurtimox | 24 | 17.4 |
| 84 | Espíndola et al., 2015 | DM28c (TcI) | Nifurtimox | 120 | 5.7 |
| 71 | Matiadis et al., 2021 | Tulahuen (TcVI) | Nifurtimox | 120 | 7 |
| 73 | Aguilera et al., 2019 | Tulahuen (TcVI) | Nifurtimox | 120 | 7 |
| 74 | Soares et al., 2019 | Tulahuen (TcVI) | Nifurtimox | 120 | 5 |
| 59 | Boiani et al., 2008 | Tulahuen (TcVI) | Nifurtimox | 120 | 7.7 |
| 88 | Chacón-Vargas et al., 2017 | INC-5 (TcI) | Nifurtimox | 24 | 8.74 |
| 89 | Domínguez-Díaz et al., 2021 | NINOA (TcI) | Nifurtimox | 48 | 7.09 |
| 89 | Domínguez-Díaz et al., 2021 | TH (ITRI/MX/2018/TH) (TcI) | Nifurtimox | 48 | 19.3 |
| 73 | Aguilera et al., 2019 | Tulahuen (TcVI) | Miltefosine | 120 | 8 |
In vitro normalized susceptibility indices (IC50, PGI50, CC50 ID50). DTU: Discrete typing unit, h: Hours.
The treatment exposure time, defined as the period from treatment addition to measurement of phenotypic changes, varies considerably. The minimum value is 24 h, while the maximum is 264 h (II times higher). 72 h predominates in 50 tests, followed by 120 h in 20 tests, 24 h and 48 h (II tests each), 96 h (8 tests) and finally 168 h and 264 h (3 tests each), Figure 4. On T. cruzi Y (TcII), the Benznidazole inhibition of parasite multiplication requires less quantity of drug when increasing exposure time (in the interval 24-72 h), a trend similar to that reported by Vela et al. However, this parameter stabilizes after 72 h, remaining consistent up to exposure times as long as 264 h. This preliminary evidence allows to visualize an optimal treatment exposure time so that this parameter can be harmonized, Table 1.
Treatment exposure time is a crucial parameter in pharmacological tests. It directly impacts the challenge faced by the test substance and can significantly influence the final results. Within timeframes that are realistic from chemical and pharmacokinetic perspectives, different exposure times for the same substance can lead to varying outcomes. This variability in results due to treatment exposure time can have important implications. It may lead to incorrectly rejecting the null hypothesis when it is actually true. These potential errors underscore the importance of harmonize and carefully considering the treatment exposure time in experimental design, Table 1.
A treatment exposure time of 72 h was most common among the reports meeting the selection criteria (even when refining the two references that bring together 22 tests, Figure 4), under these condition, various T. cruzi strains exhibited different susceptibilities to Benznidazole. The strain RA (Tc VI) showed he highest susceptibility, with an IC50 of 5,4 /μM 31,32. In contrast, strain 893 (TcIII) was the most resistant with an IC50 of 74.9 /μM 29 - a value 14 times higher than that of strain RA. This information can guide researchers in selecting appropriate challenge conditions when evaluating test substances. As efforts to harmonize conditions for pharmacological activity tests progress, this data will be further refined, potentially leading to more standardized and comparable results across studies, Figure 5.
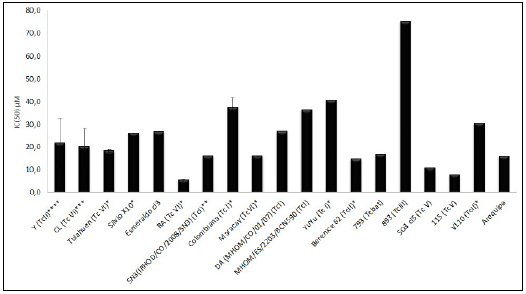
The results represent the mean ± SD of in vitro normalized susceptibility indices (IC50, PG150, CC50 ID50). *2 Independent tests, ** 3 Independent tests, *** 4 Independent tests, **** 7 Independent tests, the other strains I report.
Figure 5: Benznidazole as an active control: T cruzi multiplication inhibition at 72h treatment exposure.
The use of 96-well plates was most common (30 %, 32 tests), closely followed by 24-well plates (29 %, 31 tests). Notably, over a quarter of the test did not report the plate type used (28 tests). 15 reports used tubes or culture boxes (7 and 8 respectively). Regarding the number of parasite strains, the vast majority of reports (84 %) used a single strain. A small number of studies used two or three strains (5 % and 8 % respectively). Extreme values of eight and fourteen strains per article are reported on one occasion each.
Discussion
During its life cycle, T. cruzi alternates among different morphological and functional forms and involves both mammalian and triatomine bug hosts. The various parasitic stages of T. cruzi can be cultured in vitro using either axenic cultures or mammalian cells. Different forms of the parasite can be obtained or induced in laboratory conditions: epimastigote forms can transform into trypomastigote forms through nutrient or culture media suppression, epimastigote forms can be obtained from blood trypomastigotes, intracellular amastigotes or culture-derived trypomastigotes (by infecting mammalian cell cultures with trypomastigote forms) and axenic amastigotes can also be cultured 33,34. However, despite advances in maintaining these different evolutionary forms in the laboratory, it remains unfeasible to establish uniform culture conditions valid for all stages. This challenge arises from their distinct biological developmental contexts: trypomastigote and amastigote are found in the vertebrate host, and epimastigote and metacyclic trypomastigotes are found in the invertebrate host.
Furthermore, the morphogenetic and cell cycle modifications that occur during the progression of these three parasitic forms in their complex life cycle 13,34.
The determination of T. cruzi susceptibility in pharmacological activity tests is complex due to two main aspects: the first, mentioned above (related to the diversity of parasite evolutionary forms) confers wide variability in terms of morphology, cell tropism, infectivity, virulence, antigenic composition, metabolic activity, adaptation to the culture medium, drug susceptibility and natural resistance of the different known strains; and the second aspect is directly related to the experimental conditions under which the pharmacological activity tests is performed, which include the strain, the culture medium (and supplementation), the Infective inoculum and the interaction time between the parasite and the test substances, among others 18. Given these complexities, a systematic approach focusing on parasite forms is necessary. This approach aims to identify the most representative variables of both the biological model and the pharmacological test. The goal is to propose initial conditions for harmonization, enabling different research teams to develop equivalent and comparable tests.
The use of in vitro pharmacological models for screening trypanocidal activity offer several advantages related to costs and biosafety. These benefits include: In vitro T. cruzi cultivation allows for a significant reduction in the number of animals used in experimental infection models. This contributes to the ethical aspects of animal reducing and replacement in research (3 Rs principle). In vitro models generally require less laboratory infrastructure compared to in vivo methods. These models allow for reduced experiment duration compared to in vivo methods, potentially accelerating the research process. In vitro cultivation procedures present a lower risk of human infection during culture procedures, improving overall laboratory safety. Finally the combination of reduced animal use, lower infrastructure needs, and shorter experiment times can lead to significant cost savings in research 34.
In general, the Y strain of T. cruzi is the most frequently used in susceptibility testing, both in vitro and in vivo. This strain is characterized by high virulence and mortality in animal models, giving it clinical relevance and making it an appropriate biological model for pharmacological testing 35,36. However, it is crucial to recognize the biological diversity of T. cruzi. Whenever possible, it is essential to include several strains in anti-trypanosomal activity assays, this is particularly important for strains belonging to the DTU's: TcI, TcII, TcV and TcVI which are associated with human infection 7. This approach enhances the translational potential of preclinical studies, potentially leading to more effective treatments for Chagas disease.
Our global analysis found that amastigotes were the most commonly used parasitic form in in vitro pharmacological tests to determine T. cruzi susceptibility, followed by trypomastigotes and epimastigotes (Figure 2). Vela et al.18, reported epimastigotes as the most commonly used parasitic form, in pharmacological tests to determine sensitivity of different parasitic forms to benznidazole (47 %), followed by amastigotes (29 %) and trypomastigotes (24 %). The preference of amastigotes (104 tests) in drug sensitivity testing is highly relevant. This parasitic form is replicative in the human host and predominates in the late phase of the disease, when different organs may be affected, including cardiac complications leading to death. Moreover evidence about the effectiveness of benznidazole in this disease phase remains contradictory, driving research focus towards determining compounds efficacy against amastigotes 5,7.
Epimastigotes possess distinct metabolic, proteomic, and ultrastructural organization compared to trypomastigotes and amastigotes, suggesting that drug targets likely differ substantially between these parasite forms. While researchers currently discourage using extracellular epimastigote forms as primary pharmacological model for human and mammalian infections, these forms remain valuable for complementary screening, particularly as experimental evidence indicates potential oral transmission routes 96,97. Moreover, these forms allow for high parasite multiplication rates in a short time and at a low cost compared to intracellular forms, which require prior mammalian cell culture. However, it is important to note that test molecule activity may appear more promising in extracellular forms due to direct parasite-molecule interaction. This interaction may be reduced or absent when the substance must cross cell membranes to contact parasite. Therefore, a multistage testing is crucial: initial in vitro screening with intracellular amastigotes, complementary screening with extracellular forms, and then scaling up to in vivo models testing. This stepwise approach ensures a comprehensive evaluation of potential trypanocidal compounds 5,7.
Pharmacological model of in vitro culture of T. cruzi epimastigotes
For susceptibility testing of epimastigote forms, strain Y (TcII) is the most frequently used 33,37-60. Other commonly used DTUs include TcVI (strains CL, Tulahuen) 29,30,42,43,55,59,61-76 and TcI (strain DM28c) 77-84, Figure 3. These strains are associated with human infection, as recommended by Fonseca-Berzal et al.7. Our analysis of pharmacological tests using benznidazole as an active control against epimastigote forms revealed the use of 23 strains distributed across DTUs as follows: TcI 43.5 % 29,30,72,75,76,78,79,81,83-90, TcII 17,4 % 29,30,38,42-45,47-54,56-58, TcIII 4.3 % 29, TcV 13 % 29,30,72,75,76, TcVI 17.4 % 29-32,42,43,55,59,67,72,75,76,91,92 and Tcbat 4.3 % 28 -TcIV strains were not reported-. This information is summarized from 55 articles including 65 assays. Table 2. Vela et al.18, reported different frequencies in their analysis: TcI 69 %, TcII 12 %, TcIII 4 %, TcV 8 % and TcVI 6 % -TcIV strains were not reported-. The choice of strain for a pharmacological model should prioritize human infection-related DTUs (TcI, TcII, TcY and TcVI). This focus ensures that the testing is most relevant to clinical applications and potential drug development for human Chagas disease.
Table 2 Assays of epimastigote susceptibility to benznidazole across T. cruzi Discrete Typing Units (DTUs).
| T. cruzi DTU | No. articles | No. assays | No. strains |
| TcI | 16 | 21 | 10 |
| TcII | 18 | 23 | 4 |
| TcIII | 1 | 1 | I |
| TcV | 5 | 5 | 3 |
| TcVI | 14 | 14 | 4 |
| Tcbat | 1 | 1 | 1 |
| Total | 55 | 65 | 23 |
The most frequently used culture media for T. cruzi are LIT, BHI, RPMI 1640, and MTL. These media are typically enriched with FBS at various concentrations, with 10 % being the most common, followed by 5 % and 20 %. The FBS enrichment may vary depending on the state of the parasite culture. However, for consistent and comparable results, test conditions should be reported and harmonized when possible. A commonly recommended standard is the use of LIT medium enriched with 10% FBS.
Pharmacological test conditions for T. cruzi epimastigotes
In in vitro antitrypanosomal test for epimastigotes forms, a standardized and consistent parasite population is crucial for reliable and reproducible results. The infecting inoculum (parasites/mL) directly relates to the challenge faced by the substance to be evaluated. Therefore, this parameter should be homogenized to achieve reproducibility across different laboratories. Growth kinetic studies of various parasite strains, under standardized conditions, are necessary to identify the optimal density. This density should allow robust parasite growth without overcrowding the culture while enabling the detection of drug-induced effects. This review indicates that an initial infection inoculum of approximately 1 x 106 parasites/mL can serve as a basis for optimization and harmonization processes 37,41,44,46,49,52,56,57,59,64,69,71,73,74,89,93. It's important to note that two reports involving 14 and 8 assays each 29,30 significantly deviate this trend (Figure 4). Therefore, our refined analysis considered only the reports presenting three or less assays each, which represent 97% of the total reports. In the assessment of experimental anti-infectious pharmacology studies, the inoculum concentration should be part of a critical evaluation of methodological quality (assessment of risk of bias) of the evidence sources.
The size of the culture plate well influences the appropriate parasite density. Larger wells may require higher parasite densities to effectively cover the surface, and a consistent ratio of parasites per unit volume is required when using different types of plates. This review found the following distribution of culture plate types: 96-well plates predominated in 32 assays 31,37,43-45,51,61,62,64-67,72,75,78,79,83,84,86-89,94, while 24-well plates were used in 31 assays (though 22 of these derived from only two reports) 29,30,48,49,53,55,56,68,90. Additionally, 28 assays did not report this condition. The duration of the test (treatment exposure time) is another crucial factor in determining optimal parasite density. For extended pharmacological tests, researchers must consider parasite replication and growth over time. Among the studies reviewed, a treatment exposure time of 72 hours was the most commonly reported duration for antitrypanosomal test on epimastigotes.
The final parasite density levels must be compatible with the limits of the chosen quantification method. For example, if microscopy is involved, the final parasite density must be sufficient to allow easy visualization and quantification. The traditional method for parasite quantification is counting parasitic forms using a Neubauer chamber. Our review found this method to be the most used due its simplicity and low cost. However, its usefulness is compromised when evaluating a large number of samples, as it is time-consuming and may be subject to counting errors 95,99,100. Other methodologies that have gained popularity measure metabolic activity through colorimetric and fluorescence methods. These are efficient for high-throughput screening as they improve productivity in the number of compounds tested. However, in developing countries, few laboratories have the economic and technical capacity to implement these new methods 7,24,95. Turbidimetry (referred to as absorbance 600nm) has gained prominence as a quantification method. While turbidity-based quantification doesn't require the addition of any stain, it's essential to establish a standard curve relating turbidity to parasite density using a reference method (such as direct cell counting with a Neubauer chamber or flow cytometry) 99,100. This allows for accurate estimation of parasite concentration based on turbidity measurements. Additionally, factors such as culture conditions and the presence of debris in the culture medium can affect turbidity readings and should be taken into account during quantification. The choice of method should be based on the specific research needs, available resources, and the scale of the screening process.
Quality assessment of in vitro antitrypanosomal screening studies should include a comprehensive risk of bias evaluation. This assessment should consider several key design criteria: Presence of infection monitor groups and active control groups. Duration of follow-up (treatment exposure time). Experimental design bias (consistent time-period between treatment exposure and outcome for samples and controls). Performance of experiments in replicates necessary for adequate estimation of IC50. Measures to monitor assay performance (e.g. inter-plate and intra-plate variability). Adequate reporting (including detailed descriptions of methods, results, and statistical analyses). Criteria such as randomized assignment of treatments (plate positions) or blinding of the evaluators of the results are not commonly employed in this type of experimental pharmacology study. This structured approach to quality assessment ensures a thorough evaluation of the methodological rigor in in vitro antitrypanosomal screening studies. By considering these criteria, researchers can improve the reliability and reproducibility of their results, ultimately contributing to more effective drug discovery efforts against Trypanosoma cruzi.
Drug discovery pipelines for Chagas disease are sparse, and there remains an urgent need for new therapeutic options 10. Pharmacological models of in vitro phenotypic screening are essential tools in the search for substances with potential trypanocidal activity. These models have played a protagonist role in the recent development of anti-trypanosomatid drugs for human African Trypanosomiasis and Visceral Leishmaniasis. Drug discovery against T. cruzi is hampered by the small number of well-established targets. This limitation has led to the widespread adoption of phenotypic approaches, which have become the mainstay of Chagas disease R&D. The use of these phenotypic screening models is particularly crucial in the context of CD, where the complex lifecycle of the parasite and limited understanding of druggable targets make target-based approaches challenging.
The research and development of new drugs for CD have improved in the last years, thanks to the work of some organizations and institutions 98. However, the situation remains challenging, and new effective therapeutic alternatives for CD are lacking 4. Several promising approaches are being explored to address this need: Ethnopharmacology-derived natural products continue to be an important scaffold for affordable phytotherapeutic products. The discovery of new biologically active compounds offers potential for novel drug development. Drug repositioning presents a promising strategy to re-introduce known drugs with a history of use in humans. Considering the high attrition rate in the discovery and development of new drugs and the increasing cost of research, the harmonization of experimental conditions in pharmacological research models is a compelling need. By addressing these challenges and capitalizing on these promising approaches, the field of CD drug discovery can continue to progress towards the goal of developing effective, affordable, and accessible treatments for this neglected tropical disease.
Conclusions
Pharmacological models of in vitro phenotypic screening are an essential tool in the search for substances with potential trypanocidal activity, however harmonizing testing conditions is crucial for ensuring consistency comparability and confidence of results across different laboratories and research settings.
Scientific articles reporting the evaluation of antiparasitic activity against T. cruzi reveal a wide variety of parasite forms, parasite strains (and cell lines) and culture conditions, which makes it difficult to the effective selection of promising substances for advanced research stages. An initial approach focusing on specific parasitic forms is necessary to advance in the optimization of test conditions.
To ensure the viability and reproducibility of epimastigotes in in vitro culture models, several conditions need to be homogenized. These include strains, culture medium (and enrichment), and incubation parameters (temperature and culture medium replacement frequency). Similarly, pharmacological test conditions like infecting inoculum concentration, culture well plates specifications, duration of exposure to treatment, and method of parasite quantification must also be homogenous.
Quality assessment of in vitro antitrypanosomal screening studies involves experimental design bias (active and infection controls), duration of follow-up, measures to monitor assay performance, replicates necessary for adequate estimation of IC50, and adequate reporting. Harmonizing these conditions is crucial for improving the reliability and comparability of results across different studies, facilitating the reliable identification of potential drug candidates and supporting their further development. Moreover, it promotes the sharing of knowledge and data across the scientific community, fostering collaboration and accelerating drug discovery efforts.
Box 1 Recommended starting points for harmonizing experimental conditions in in vitro pharmacological models for assessing activity against t. cruzi epimastigotes
| Model of in vitro culture of epimastigote forms |
|
• Strains: Strain Y (TcII) is the most frequently used. Whenever possible include several strains, especially those belonging to the DTUs: TcI, TcII, TcV and TcVI, which are associated with human infection. Different T. cruzi strains show different susceptibility. • Culture medium: Liver infusion-tryptose culture medium (LIT) predominates. • Enrichment of Culture medium: Fetal bovine serum (FBS) at a concentration of 10 % is most common. FBS concentration may vary depending on the stage of the culture. • Incubation parameters: • Culture medium replacement frequency: Around every 7 d (range 3-10 d). • Incubation temperature at 28 °C. |
| Pharmacological test conditions |
|
• Infecting inoculum: Around I x 106 parasites/mL. • Culture well plates: 96-well plates are predominantly used. • Treatment exposure time: 72 h is most reported. On Benznidazole IC50 values generally decrease as incubation time increases but, stabilizing after 72 h. |
|
Method of parasite quantification: Microscopy through the Neubauer chamber is the method traditionally used for this purpose, however, limited usefulness for large sample numbers. Alternative methods correspond to colorimetric, turbidimetric (referred to as absorbance) and fluorescence-based. In turbidity measurements it's essential to establish a standard curve relating turbidity to parasite density using a reference method, culture conditions and the presence of debris can affect readings and should be considered during quantification. •Active control: The most frequently reported is Benznidazole 5.5 times more than nifurtimox. On Y (TcII) strain, at 72 h, Benznidazole shows IC50 of 21.8 +/-10.8 μM * DTU: Discrete typing unit, h: Hours. * The results represent the mean ± SD of in vitro IC50, 7 Independent reports. |

