











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
In Colombia, snakebites have been considered events of public health interest and established as a notifiable event since October 2004 1,2. However, consistent reporting only began in 2007 2. Due to the number of events registered since then, snakebites are considered a public health problem in Colombia 1. During 2023, 6,184 cases were registered in the National Public Health Surveillance System 3. In Colombia, snakebites involving snakes of the Bothrops genus represent the most significant number of reported cases, being considered the most epidemiologically relevant accidents in the country, with 80-85% of the cases 2. The local tissue damage observed in these envenomations can lead to several sequelae, such as limb amputation or loss of muscle mass 4-7. Currently, the only available treatment for this health problem is antivenom administration. However, it cannot neutralize local effects induced by snake venoms 1,5,6.
The venom of Viperidae family snakes can be divided into three main components: enzymes, toxins, and a non-protein component 8. Within the enzymatic group, the most representative components are phospholipases A2 (PLA2s), snake venom metalloproteinases (SVMPs), and snake venom serine proteinases (SVSPs) 5. PLA2 causes hydrolysis of the bond at the sn-2 position of a cell membrane glycerophospholipids, resulting in a free fatty acid and a lysophospholipid. This reaction triggers a series of events in the organism, including local effects (such as local myotoxicity and edema) and systemic effects (such as systemic myotoxicity, hypotension, hemolysis, platelet aggregation modulation, cardiotoxicity, and direct organ damage, mainly liver, kidney, and lung) 5,7.
Traditionally, some plants have been used to treat snakebites, and these plants have been shown to have positive effects such as anti-inflammatory activity and inhibition of snake venom’s PLA29,10. Ethnopharmacology has attempted to study these plants and isolate the compounds with the greatest benefits in these envenomations to evaluate them and determine their pharmacodynamics 9-12. Pharmacologically active compounds that have been described with beneficial effects in snakebites are secondary metabolites, mainly phenolic compounds (like flavonoids, phenolic acids, coumarins, and coumestanes), alkaloids, steroids, terpenoids, and polyphenols 9,11-14.
The inhibitory capacity of pentacyclic triterpenes against the main enzymes found in snake venoms has been previously assessed 12,13,15. Indeed, some of these molecules have shown inhibition on a snake venom metalloproteinase isolated from Bothrops atrox from Colombia 16,17. Some of these evaluations have utilized bioinformatics tools, such as molecular modeling, to identify chemical compounds in plant extracts that exhibit inhibitory effects on the PLA2 enzyme 12,13. The results have shown that some of these compounds inhibit the activity of venom PLA2s; in addition, other pentacyclic triterpenes have been found to inhibit the coagulant activity induced by the serine proteinases 15; among these triterpenes are madecassic acid, oleanolic acid, ursolic acid, betulinic acid, betulin, 3-epilupeol formiate, α-amyrin acetate, 3-epilupeol acetate, lupenone, 3-epilupeol, and α-amyrin 12,13,15.
Based on these results and others reported in the literature of similar compounds with inhibitory activity on PLA215, this study aimed to evaluate the inhibitory activity of four pentacyclic triterpenes (betulinic acid, madecassic acid, ursolic acid, and oleanolic acid) on a myotoxic PLA2 purified from Crotalus durissus cumanensis venom, and using molecular docking tool identify the possible interaction between these compounds and the active site of the enzyme.
Methods
Chemical and reagents.
Betulinic acid, oleanolic acid, ursolic acid, and madecassic acid were purchased from Santa Cruz Biotechnology (Dallas, TX, USA). Other reagents used in this work were of the highest purity available from Sigma Aldrich (St. Louis, MO, USA) and Merck (Kenilworth, NJ, USA).
Purification of PLA2
PLA2 was purified from the lyophilized venom of C. durissus cumanensis, obtained from four specimens from the Department of Meta (southeast region of Colombia), and kept in captivity at the serpentarium of the University of Antioquia (Medellín, Colombia). PLA2 was purified following the method described by Pereañez et al. 18 with modifications. Briefly, a bioguide fractionation was performed by quantifying the PLA2 activity of each fraction. The enzyme was purified by molecular exclusion chromatography on Sephadex G-75, followed by reverse-phase HPLC on a C18 column eluted at 1.0 mL/min with a gradient from 0% to 100% acetonitrile in 0.1% trifluoroacetic acid (v/v). A micro-UV/VIS NanoDrop Spectrophotometer was used to quantify the toxin. Absorbance in effluent solution was recorded at 215 nm.
Purity Evaluation of the obtained PLA2
RP-HPLC was used to determine protein purity, on a C18 column eluted at 1.0 mL/min with a gradient from 0% to 70% acetonitrile in 0.1% trifluoroacetic acid (v/v) for 25 minutes. A micro-UV/VIS NanoDrop Spectrophotometer was used to quantify the toxin. Absorbance in effluent solution was recorded at 215 nm.
Inhibition of PLA2 activity using 4-nitro-3-octanoyloxybenzoic acid (4N3OBA) as a synthetic monodisperse substrate.
Measurements were performed according to the method described by Holzer et al.19, using a 96-well ELISA plate method with 25 μL of 4N3OBA (Santa Cruz Biotechnology, Dallas, TX, USA) (1 mg/mL) each. Four pentacyclic triterpenes were used to evaluate their inhibitory capacity of PLA2 activity: betulinic, oleanolic, ursolic, and madecassic acids. For the wells carrying the compounds, 80 μg of PLA2 and a 500 μM solution of each compound dissolved in buffer (10 mM Tris-HCl, 10 mM CaCl2, 100 mM NaCl, pH 8.0) were used. Positive (PLA2 + buffer), negative for each compound (compound + buffer), and negative (only buffer) controls were performed. The mixture was incubated for 60 min at 37 ºC, and absorbance was measured at 425 nm, n=4. Results are described in percentage of inhibition, taking toxin and PBS as 100% and 0% of activity, respectively.
Molecular docking
Molecular docking was performed using AutoDock Vina 20. The structure of phospholipase A2 from the venom of Crotalus durissus terrificus (PDB code 2QOG) was used as a model structure for the computational studies, which was prepared using the programs UCSF Chimera 1.14 21 and AutoDockTools. In UCSF Chimera 1.14, foreign elements such as water molecules and residual solvent present from crystallization were removed from the protein, whereas in AutoDockTools, polar hydrogen atoms were added, atoms were assigned the AD4 type, and atomic charges were assigned. A charge of +2 was set to the calcium cofactor of phospholipase A2. Likewise, the structures of the four compounds (betulinic acid, madecassic acid, ursolic acid, and oleanolic acid) were downloaded from the MolView database 22 and prepared using the Avogadro program 23, and hydrogen atoms were automatically added according to the ionized form found under physiological conditions. The coupling was performed in AutoDock Vina, using the calcium ion as the center of the grid (X= 61.179, Y= 42.712, and Z= 47.465), an exhaustivity of 20 and a mesh size of 16 Å3. Finally, the interactions between the compounds and the enzyme active site were visually inspected using the UCSF Chimera 1.14 21, and PLIP 24 was used to generate the docking images.
Statistical analysis.
A Shapiro-Wilk test was used to determine whether the data sets adjusted to a normal distribution. The data obtained were subjected to a one-way ANOVA, followed by a Bonferroni post hoc test. The significance level was 0,05. Data are shown as mean ± standard deviation, n=4. Analysis was performed with Python (Version 3.10).
Results
Purification of PLA2
Lyophilized venom of C. durissus cumanensis was first subjected to molecular exclusion chromatography on Sephadex G-75. The fraction with PLA2 activity on 4N3OBA eluted in about 10 minutes and is consistent with the previous isolation 18. This fraction was concentrated in a SpeedVac. Then, 6 mg of this fraction was submitted to a RP-HPLC, under the conditions described in methodology, and the average yield of PLA2 obtained was around 20%. Figure 1 shows the molecular exclusion chromatography and RP-HPLC profiles corresponding to the purification of the toxin, in which the peak obtained around 60 min with a concentration of acetonitrile at 75% belongs to PLA2, which is also the major compound in the fraction.
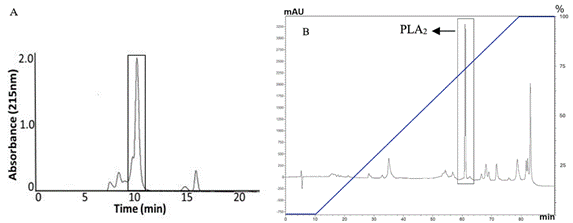
Purity Evaluation of the obtained PLA2
An RP-HPLC run was performed to verify the purity of PLA2 obtained, in which a single peak was observed around 18 min with a concentration of acetonitrile at 52%, indicating that the toxin had been successfully isolated from the venom of C. durissus cumanensis (Figure 2).
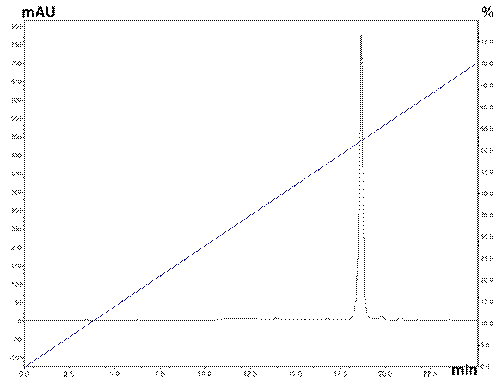
Inhibition of PLA2 activity using 4-nitro-3-octanoyloxybenzoic acid (4N3OBA) as a synthetic monodisperse substrate.
In this method, p-nitroanilide is released from the substrate after the hydrolytic action of the PLA2. This chromophore has its maximum absorbance at 425 nm. Therefore, it is expected that when inhibitors are tested, the absorbance decreases. Absorbance obtained in the test with the synthetic substrate for each compound, including the positive control (PLA2), are recorded Figure 3. The best inhibitory activity was those for ursolic and madecassic acids (p<0.001, regarding positive control). In contrast, oleanolic acid (p<0.01) and betulinic acid (p<0.05) showed lower inhibitory ability on PLA2 of the toxin.
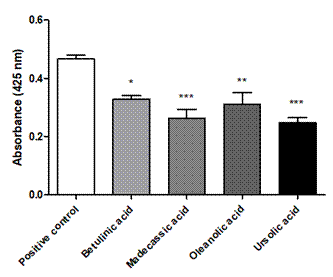
Figure 3 Absorbance results for the test with the synthetic substrate, the triterpenic compounds, and the positive control (80 μg PLA2). Negative for each compound (compound + buffer), and negative (only buffer) controls were also performed. (*) Statistically significant differences compared with the absorbance induced by the positive control with a p-value ˂ 0.05. (**) P-value ˂ 0.01 (***) P-value ˂ 0.001.; n=4.
Molecular docking
A molecular docking study was performed to suggest a mode of action for the pentacyclic triterpenes. The results indicated that all compounds could bind to the PLA2 (Figure 4). Three of the four compounds had a similar conformation and binding site; however, ursolic acid had a different mode of action and binding site.
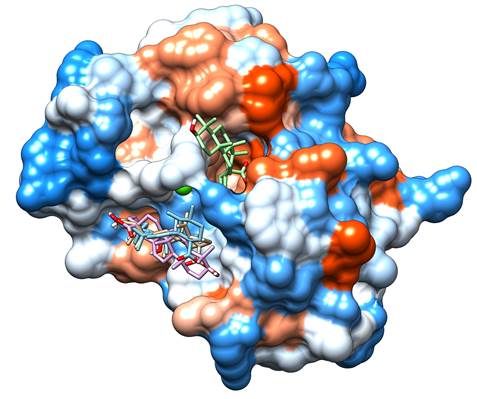
Figure 4 Molecular docking results. Betulinic acid is shown in beige, madecassic acid in blue, oleanolic acid in purple and ursolic acid in green. The red areas of the surface represent the acid regions, the white areas represent the neutral, and the blue areas are basic regions. The green sphere represents Ca2+. Images were obtained using UCSF Chimera 21.
When evaluating the type of interactions between the inhibitors and the enzyme, it is observed that most of these interactions are Van der Waals; however, hydrogen bonds are observed between madecassic acid and Tyr28 and Asp49; betulinic acid and Tyr28; and oleanolic acid and Gly33 and Thr68 (Figure 5 and Table 1).
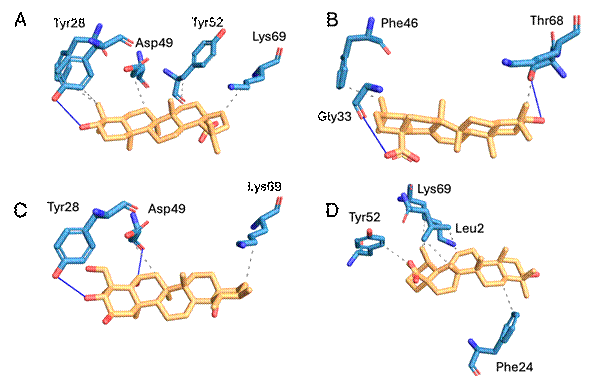
Figure 5 Binding modes of triterpenic acids into PLA2 binding pocket. A: Betulinic acid. B: Oleanolic acid. C: Madecassic acid. D: Ursolic acid. Gray dashed lines represent Van der Waals interactions; blue lines represent hydrogen bonds. Images were obtained with PLIP 24.
Table 1 Suggested interaction for each evaluated compound and PLA2.
| Interaction | Van der Waals | Hydrogen bond | Affinity (kcal/mol) |
|---|---|---|---|
| Madecassic acid | Asp49 and Lys69 | Tyr28 and Asp49 | - 5,8 |
| Betulinic acid | Tyr28, Phe46, Asp49, Tyr52 and Lys69 | Tyr28 | - 6,6 |
| Oleanolic acid | Phe46 and Lys69 | Gly33 and Thr68 | - 6,2 |
| Ursolic acid | Leu2, Phe24, Tyr52, Lys69 | - 6,8 |
Discussion
The PLA2s are the most abundant enzymes in Viperidae snake venoms 7,25. In addition, they contribute to the local tissue damage that can derive in sequelae, such as amputations and tissue loss 4,7. Therefore, an important goal in snakebites is to inhibit or neutralize the catalytic activity of the PLA2s 25,26. An enzyme from C. durissus cumanensis was isolated to test the inhibitory potential of triterpenic acids on snake venom PLA2. According to the results, the purification of the PLA2 enzyme was consistent with the previously reported procedure in terms of enzyme activity and purification parameters (retention times and acetonitrile concentration) 18.
The tested triterpenic acids inhibited the PLA2 activity of the enzyme in different percentages, 47,01% (ursolic acid), 43,77% (madecassic acid), 33,57% (oleanolic acid), and 29,60% (betulinic acid). These results may be explained by the binding modes suggested by molecular docking results. All compounds exhibited a Van der Waals interaction with Lys69, which is a conserved residue involved in the interfacial binding surface of the enzyme that contributes to the activation in the lipid-water interface 26,27. Other important interactions are Van der Waals and hydrogen bonds of betulinic and madecassic acids with Asp49, respectively. This amino acid is responsible for Ca2+ coordination, which is required for the oxyanion stabilization during catalytic cycle 26,27.
The most active compound was the ursolic acid, which showed several Van der Waals interactions with Leu2, Phe24, Tyr52, and Lys69. Phe24 is involved in the interfacial binding surface, therefore, affecting substrate recognition by the enzyme 26,27. In contrast, Leu2 is located in the hydrophobic channel of the PLA2, which is involved in the stabilization of the glycerophospholipid in the active site of the enzyme 26,27. The other interaction of ursolic acid was with Tyr52, which is responsible for maintaining the hydrogen bond network of the catalytic site of the enzyme 26,27. Finally, the interaction with Lys69 was described previously.
Results obtained in this work agree with those reported by Dharmappa et al., who reported the ability of oleanolic acid to inhibit the PLA2 activity of Naja naja and Vipera russelli venoms. The authors determined that the inhibition process is independent of the amount of substrate or the presence of calcium ions, also demonstrated that oleanolic acid interacts directly with the enzyme, forming a complex, that irreversibly inhibits PLA2 activity 28. On the other hand, Nataraju et al., reported similar findings for ursolic acid by identifying that this compound has inhibitory action against phospholipase A2 isolated from the venoms of Vipera russelli and Naja naja snakes 29.
Triterpenic acids also have been reported by their inhibitory effects on SVMPs, by inhibiting the catalytic, hemorrhagic, myotoxic, and edema-forming activities of a PI-SVMP from B. atrox venom 16. In addition, it was demonstrated that the carboxylate ion interacts with the zinc ion, an essential cofactor for SVMP activity 17. Therefore, these compounds are promising molecules that can find active leads against local tissue damage induced by snake venoms.
Conclusion
The method used for the extraction, purification, and evaluation of the toxin was effective, allowing us to determine the inhibitory activity of different triterpenic compounds against the toxin. Through the method using the synthetic substrate to evaluate the inhibition of PLA2 enzymatic activity, it was determined that ursolic acid presented greater inhibitory capacity compared to the other compounds evaluated, with a percentage of 56.79% (p < 0.001), being the most promising compound to reduce or inhibit the local effects that PLA2 can cause. With the results obtained in the molecular docking, a hypothesis can be established as to why ursolic acid presents a greater inhibitory capacity against the enzymatic activity of PLA2, since it presents the highest affinity with the enzyme and shows interactions with the amino acids Leu2, Phe24, Tyr52, Lys69, all involved in important functions on the enzyme structure and catalysis (hydrophobic channel, active site, and interfacial binding surface).
Further studies should be conducted to determine other potential benefits of these compounds against other biological actions of PLA2 that could not be addressed in the present project, as well as formulations studies to develop therapeutic strategies as coadjutants of antivenoms.

