











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink1 Introducción
El melanoma es una neoplasia causada por la transformación maligna de los melanocitos, encargados de la producción de melanina como barrera de protección frente a los rayos UV, que desencadena la pérdida de control de los mecanismos de crecimiento y proliferación celular [1]. En los últimos años, la incidencia del melanoma a nivel mundial ha sido del 1,7 % y la población más afectada es la caucásica, que presenta una incidencia del 3 al 5 %. Aunque el melanoma representa menos del 5 % de los casos de cáncer de piel, alcanza mortalidades cercanas al 80% [2].
Este tipo de cáncer se caracteriza por la presencia de la mutación BRAF V600 y alteraciones en los genes MEK, NRAS, c-KIT, etc., involucrados, junto con genes como CDKAN2A y MCR1, en la activación de la vía de señalización MAPK/ERK en melanoma familiar [3]. Adicionalmente, sobreexpresión de receptores de puntos de control inmune como la molécula de muerte programada 1 (PD-1/CD279) y el Antígeno-4 asociado al Linfocito T Citotóxico (CTLA-4/CD152), han sido asociados con el escape de las células tumorales a la inmunovigilancia mediante la manipulación del microambiente local y sistémico [4, 5]. Lo anterior conduce a la destrucción de las respuestas inmunitarias innatas y adaptativas, debido a la capacidad de las células tumorales de enviar señales al sistema inmunológico para ralentizar estas reacciones [6].
En pacientes con melanoma avanzado, la terapia dirigida con inhibidores BRAF e inhibidores de MEK, junto con terapias inmunitarias y anticuerpos monoclonales (mAb) de anti-CTLA-4 y de anti-PD-1, han sido el centro de atención para el desarrollo de estrategias terapéuticas de alta eficacia y baja toxicidad [7-10]. A su vez, las terapias combinadas empleando inhibidores del punto de control inmunitario (CPI por sus siglas en inglés) anti-PD-1, más anti-CTLA-4 o anti-PD-L1, han mostrado mejores resultados en la supervivencia total, en comparación con la monoterapia anti-PD-1, que presentó resistencia a la terapia con progresión de la enfermedad en el 40 % de los pacientes [11, 12]. Con el objetivo de superar tal resistencia, han sido identificados potenciales blancos inmunoterapéuticos, que incluyen al gen de activación de linfocitos 3 (LAG-3) y al receptor inhibitorio con dominios Ig e ITIM (TIGIT), los cuales actúan como receptores de los puntos de control inmune que desempeñan un rol fundamental en la inmunología tumoral [13-16].
LAG-3 es un receptor que actúa como regulador negativo de la respuesta inmunitaria, expresado en células T reguladoras (Tregs), linfocitos infiltrantes de tumores (TLI) CD4+, CD8+, Natural Killer (NK) y células B [17]. El complejo principal de histocompatibilidad de clase II (MHC II) es considerado el ligando canónico de LAG-3 y junto con ligandos como la proteína similar al fibrinógeno 1 (FGL1) y Gal-3 bloquea la proliferación, activación y homeostasis de las células T [18], promoviendo el crecimiento tumoral al inhibir la respuesta antitumoral mediada por CDK2, CDK6 y CDK4 cuando se une con LSECtina [19]. Recientes estudios han indicado que el bloqueo conjunto de LAG-3 y PD-1, además de restaurar la función antitumoral de las células T [20, 21], conduce a una mayor supervivencia libre de progresión (47 %) en melanoma no resecable o metastásico en contraste con la inhibición de PD-1 [16,22].
El inmunoreceptor de células T (TIGIT) es un potencial blanco inmunoterapéutico, debido a que constituye un punto de control de la respuesta inmune innata y adaptativa. Este receptor se expresa en células T, NK activadas, Tregs, CD8+ y CD4+, y se coexpresa con LAG-3 y con la inmunoglobulina de células T mucina-3 (TIM-3). Dentro de los ligandos para este receptor se encuentran Nectina-4, CD155, CD112, entre otros, los que generalmente se sobre-expresan, siendo indicadores de mal pronóstico en melanoma [13, 23].
A pesar de la gran información existente sobre los receptores del punto de control inmunitario LAG-3 y TIGIT en cáncer, el rol de estos en la respuesta inmune de las células presentes en el microambiente tumoral en melanoma, no está claro. Considerando lo anterior, el objetivo de esta revisión es abordar el conocimiento actual alrededor de la influencia de los receptores LAG-3 y TIGIT/Nectina-4 en la respuesta inmune antitumoral y los avances en monoterapia y terapia combinada dirigida a estos receptores en melanoma.
2 LAG-3 Y TIGIT en melanoma
2.1 LAG-3 como punto de control en melanoma
LAG-3 (CD223) pertenece al grupo de la superfamilia de las inmunoglobulinas. Su expresión en células T CD4+ y CD8+ es inducida por el interferón beta (IFN-ß) y la interleucina-27 (IL-27), a diferencia de TIGIT, el cual disminuye su expresión en las mismas condiciones [24]. De igual manera, otras citoquinas como IL-2, IL-7, IL-12 e IL-15 promueven la expresión de LAG-3 [25].
El receptor LAG-3 presenta 4 dominios extracelulares similares a Ig (D1-D4) que interactúan con ligandos específicos. Los dominios extracelulares tienen afinidad al ligando MHC-II tanto en LAG-3 como su homólogo, el receptor CD4 [26], los cuales presentan 20 % de similitud estructural, debido a un evento de duplicación de genes [25, 27, 28]. Particularmente, la variación en la conformación estructural de D1-D4 es la responsable de la unión específica de MHC-II y FGL1, así como las diferentes funcionalidades de LAG-3 al interactuar con sus ligandos [29, 30]. En la región intracelular de LAG-3, el dominio KIEELE es necesario para que se produzca la transducción de la señal inhibitoria [31]. Sin embargo, recientes estudios identificaron que dos señales inhibitorias se transducen independientemente en los motivos FXXL en la región proximal a la membrana y la repetición EX en el extremo C-terminal en la sección citoplasmática del receptor [32].
Ha sido indicado que la expresión de LAG-3 y MHC-II se relaciona positivamente en melanoma [33]. De hecho, la señalización producida por la interacción LAG-3/MHC-II en células dendríticas reveló un patrón de fosforilación en tirosina que induce la activación de las proteínas PLCy2 y p72syk, y de las vías de señalización PI3K/Akt, p42/44ERK y p38MAPK [34]. Así mismo, ha sido reportado que las vías MAPK/Erk y PI3K/Akt activadas por fármacos que dañan el ADN y la proteína Fas respectivamente, están involucradas en la limitada actividad apoptótica de las células de melanoma con expresión LAG-3/MHC-II (Figura 1) [35]. De resaltar que recientes estudios, demostraron que la unión de LAG-3 al ligando MHC-II es selectiva y se produce a través de complejos estables de péptidos antigénicos unidos a MHC clase II (pMHC-II) específicos. Razón por la que la asociación de MHC-II al receptor CD4 no se ve interrumpida. Dicha interacción LAG-3/MHC-II en células tumorales y células presentadoras de antígeno (APC), suprime la actividad de células T [36]. LAG-3/MHC-II también promueve la activación de células dendríticas plasmocitoides (pDC) asociadas a melanoma, las cuales incrementan la secreción de IL-6 y disminuyen la producción de IFNa, generando un microambiente tumoral inflamatorio exacerbado que promueve la progresión del tumor [37]. Además, en ensayos in vivo se demostró que la pérdida de la unión estable de pMHCII a LAG-3, incrementó la actividad inmunitaria contra el cáncer [38, 39]. Sin embargo, es de resaltar que incluso LAG-3 sin interacción con MHC-II es capaz de limitar la activación de células T CD4+ y CD8+. Lo anterior debido a que la estructura de la cola citoplasmática de LAG-3 presenta una repetición en tándem de ácido glutámicoprolina, la cual provoca una disminución del pH en la sinapsis inmunológica (SI) donde se reclutan las proteínas que intervienen en la interacción entre células T y APC [40, 41]. Esto produce la disgregación del complejo tirosina quinasa Lck/CD4 y, genera una alteración en la señalización de TCR (Figura 1). Así, LAG-3 se desplaza hacia la SI para asociarse con el complejo TCR/CD3 [41], interrumpiendo la señalización aguas abajo de CD3+.
Dentro de los ligandos principales de LAG-3, se encuentra la proteína FGL1. Esta proteína es producida en el hígado y en el páncreas, y es secretada a órganos periféricos por medio de la sangre. De resaltar, que la proteína FGL1 también se sobreexpresa en melanoma metastásico. En especial, la forma oligomérica de FGL1 se une con alta afinidad a LAG-3 por medio de su dominio similar a fibrinógeno independientemente de MHC-II, evitando la actividad inmune de células T infiltrantes de tumor activadas en modelos de ratón [42].
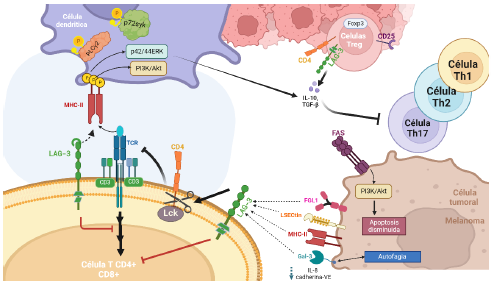
Figura 1 Efecto de la sobreexpresión de LAG-3 por la inducción de diversas citoquinas, en el comportamiento del microambiente tumoral en melanoma. La asociación de LAG-3/MHC- II conduce a la activación de varias vías de señalización, promoviendo la producción de citoquinas inmunosupresoras que contribuyen al microambiente tumoral inflamatorio, a la sobreexpresión de LAG-3 en células T CD4+ y CD8+, a la disminución de la autofagia, a la reducción de la apoptosis y a la progresión del melanoma.
Por otro lado, diferentes estudios han demostrado la asociación de la expresión transcriptional de LAG-3 con otras moléculas y su influencia en la respuesta inmune contra el melanoma. Por ejemplo, un estudio reciente realizado en pacientes con melanoma uveal (MU), mostró que la expresión de ARNm de ligandos como LSECtina y Galectina-3 (Gal-3) están asociados positivamente con la expresión de LAG-3 y con baja tasa de supervivencia [33]. De destacar que la sobreexpresión de LSECtina ha sido evidenciada en células de melanoma humano, en donde su interacción con LAG-3 inhiben la funcionalidad secretora de interferón gamma (IFN/) [19].
En lo que respecta a Gal-3, ensayos in vitro han demostrado que este ligando se asocia con LAG-3 en células T CD8+ activadas, impidiendo su funcionalidad en el microambiente tumoral. Esta asociación sugiere la inhibición de la actividad de pDC inflamatorias, las cuales impulsan la activación de células T específicas de antígeno [43], lo cual se relaciona con la alta expresión de LAG-3 en pDC de ganglios linfáticos invadidos por metástasis de melanoma, así como en el melanoma metastásico cutáneo [37]. Dentro de las funciones descritas hasta la fecha de Gal-3 se encuentran: la disminución de la actividad autofágica, la progresión de melanoma mediante la regulación de la IL-8 y la cadherina VE, la regulación positiva de la expresión de autotaxina (ATX) a través del factor nuclear de células T activadas (NFAT1) y, la progresión de melanoma metastásico [44, 45]. A su vez, niveles elevados del transcrito de LAG-3 en MU han sido correlacionados con pérdida del gen BPA1 [33,46], implicado en el desarrollo de metástasis a través de la activación de la vía de señalización NFkB que se asocia con factores reguladores inmunes MHC-I [47].
Estudios adicionales en una cohorte de pacientes con melanoma cutáneo, indicaron que la regulación transcripcional mediada por la hipometilación en la región promotora del gen LAG-3, se correlaciona significativamente con la expresión de su ARNm en tipos celulares proinflamatorios como: células NK activadas, células T CD8+ y células T CD4+ activadas, lo que contribuye a un microambiente inmunosupresor [48].
De igual manera, en pacientes con melanoma en etapa III, la expresión de LAG-3 fue identificada en células Treg de ganglios linfáticos invadidos por el tumor y linfocitos infiltrados por el tumor, que a su vez expresan CD25+, Foxp3+ y CD4+ [49] (figura 1). Lo anterior, conduce a la alteración de la proliferación y la homeostasis de las células T, al incrementar la producción de citoquinas inmunosupresoras IL-10 y TGF-ß 1 en células dendríticas inducidas por el tumor, limitando la respuesta inmune de células T CD4+, T CD8+ y NK, contra el cáncer [50] y disminuyendo el fenotipo efector de linfocitos cooperadores como Th1, Th2 y Th17 [51]. Los anteriores hallazgos, han conducido al establecimiento de asociaciones entre la expresión de LAG-3 en células Treg con la producción de citoquinas inmunosupresoras en tumores de melanoma avanzado [49] (figura 1).
2.2 TIGIT y su rol en melanoma
El receptor inhibitorio con dominios Ig e ITIM (TI-GIT) pertenece a la familia de las super inmunoglobulinas, junto con LAG-3 y TIM-3. Este receptor está formado por cuatro dominios: un dominio variable de inmunoglobulina extracelular (IgV), un dominio transmembrana tipo I, una cola citoplásmica con un motivo inhibidor basado en tirosina de inmuno-receptor (ITIM) y un motivo similar a la tirosina de cola de Ig (ITT) [52]. Dentro de sus ligandos se encuentran dos miembros de la familia de las nectinas, CD155 (PVR, Necl-5) y CD112 (Nectina-2), ligandos que generan coestimulación o coinhibición y que son compartidos con otros receptores como CD226. Recientemente otro ligando fue identificado, el ligando Nectina-4 cuya afinidad es específica de TIGIT [53]. Dentro de las funciones asociadas a este inmunoreceptor se encuentran: la supresión de la proliferación de células CD8+ al unirse a los dominios del receptor de células T (TCR) [54], la reducción de la señalización de p-ERK inducida por TCR en células T CD8+ [55], la inhibición de la citotoxicidad regulada por las células NK y el bloqueo de la unión de receptores homólogos (como CD266) a los respectivos ligandos [56].
TIGIT ha sido relacionado con una regulación positiva en diferentes tipos de cánceres como el carcinoma hepatocelular [57], adenocarcinoma de pulmón [58], carcinoma de células renales [59], melanoma [60], entre otros, dónde niveles altos de CD155 o TIGIT han sido asociados con progresión tumoral, incluida la invasión tumoral y la recurrencia, e indican resultados clínicos desfavorables [53].
En melanoma, TIGIT se expresa en >70 % de los microambientes tumorales [61] y su locus específico está hipometilado en las células Tregs. Las células Tregs, que expresan una alta proporción de TIGIT en los sitios del tumor, evidencian niveles más altos de genes como FOXP3, CD25 y CTLA-4, los que contribuyen a la capacidad invasiva del melanoma [62,63]. Por el contrario, tanto las células T efectoras (Teff) como las CD4+, regulan al alza tanto a TIGIT como a CD226 e incrementan la liberación de la citoquina proinflamatoria IL-10 [60].
De la misma manera, ha sido indicado que el ligando CD155, regulado al alza en células de melanoma, suprime la activación de CTL específicos a través de la interacción con TIGIT en la fase efectora [64]. La estimulación tumoral induce la disminución de la expresión de CD226 con la consecuente regulación positiva de TIGIT en los CTL productores de IFN-7 específicos de melanoma, incrementando por ende la resistencia inflamatoria en el microambiente tumoral [14]. Además, el alza de TIGIT/CD155, reduce la citotoxicidad, la desgranulación y la secreción de citoquinas en células NK. Además, TIGIT/CD155 permite reclutar el dominio SH que contiene inositol-5-fosfatasa (SHIP1), y de esta forma inhibir las vías de señalización PI3K, MAPK y NF-kB [65, 66].
Adicionalmente, un estudio reciente reportó asociaciones entre los receptores inmunitarios, indolamina 2,3-dioxigenasa (IDO1) y TIGIT, con un aumento significativo en el número de células TIGIT+ en pacientes con MU metastásico. Cabe resaltar que los efectos de la metástasis aumentaron 18 veces más en comparación con las células TIGIT negativas [67].
Como se indicó anteriormente, el ligando Nectina-4, fue recientemente identificado como un ligando con afinidad específica para TIGIT [53]. Nectina-4 se sobreexpresa en pacientes con melanoma con mutación en BRAF V600. La sobreexpresión de esta molécula ha sido asociada con la sobreactivación de la vía PI3K/Akt en los melanocitos, constituyéndose, por ende, como un indicador de mal pronóstico [68]. Adicionalmente ha sido indicado que, en melanoma avanzado, los cambios en la expresión de Nectina-4 alteran la adhesión celular, favoreciendo el crecimiento de las células cancerosas y el aumento de la resistencia a los medicamentos, al afectar la vía de señalización de ERK [69] (Figura 2).
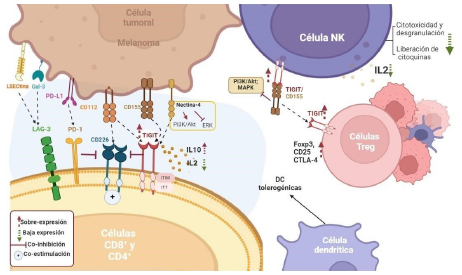
Figura 2 Efectos de la sobreexpresión de TIGIT en el microambiente tumoral en Melanoma. La unión de TIGIT a CD155 y CD112 conduce al aumento de IL-10, a la disminución de IL-2, y a la inhibición de células T. Adicionalmente, el alza de este complejo, reduce la citotoxicidad, desgranulación y secreción de citoquinas en células NK conduciendo a la inhibición de vías de señalización como la de PI3K, MAPK y NF-kB. Por su parte, en células T reguladoras (Tregs) el alza de TIGIT favorece la capacidad invasiva del tumor por el incremento en la expresión de las vías Foxp3, CD25 y CTLA-4.
3 Inmunoterapia LAG-3
Las terapias dirigidas al bloqueo de LAG-3 se han centrado en el desarrollo tanto de monoterapias (anticuerpos monoclonales, proteínas de fusión solubles LAG-3-Ig y fármacos bioespecíficos anti-LAG-3 [70], como de terapias combinadas.
3.1 Monoterapia
Diversos anticuerpos anti-LAG-3 han sido desarrollados con el objetivo de restablecer la actividad de las células T mediante la interrupción de la unión del receptor con sus ligandos. Dentro de estos mAb se encuentran LBL-007, Eftilagimod alfa y C25, entre otros. LBL-007 es un mAb desarrollado actualmente, el cual corresponde a un aislado de una biblioteca de presentación de anticuerpos humanos mediante fagos que es capaz de bloquear la unión de LAG-3 a MCH-II y LSECtina simultáneamente. Este anticuerpo permite una mejor internalización celular que Relatlimab e inhibe mejor las funciones de LAG-3/MHC-II y lectina de células endoteliales sinusoidales hepáticas. Lo anterior conduce a la disminución del crecimiento tumoral, debido al incremento en la secreción de IL-2 y del NFAT [71].
Eftilagimod alfa es un péptido soluble LAG-3 que se une a MHC-II e interviene en la activación de APC y células CD8 [72]. Estudios recientes han demostrado que la activación de células T con Eftilagimod alfa, exhibe una respuesta prolongada al tratamiento. Específicamente, se observó qué en pacientes con melanoma no resecable o metastásico, el incremento sostenido en el número de células T CD4+ y CD8+ luego del tratamiento se mantuvo durante 3 a 6 meses [73].
C25, es un nuevo péptido cíclico desarrollado por "biopanning" de visualización de fagos. Este péptido presenta una alta afinidad de unión con LAG-3, conduciendo a la interrupción de la interacción de HLA-DR (una molécula del MHC-II). Adicionalmente, ha sido asociado con la ralentización de la degradación enzimática, y con el aumento en la proporción de células T CD4+, CD8+ infiltrantes de tumor y la producción de IFN-7, en modelos de ratón [74].
3.2 Terapia combinada
La expresión de LAG-3 en células CD4+, CD8+ y pDC es considerada como biomarcador de mal pronóstico. Por lo que diferentes terapias inhibidoras han sido desarrolladas para interrumpir la unión LAG-3/MHC-II. Teniendo en cuenta que LAG-3 se coexpresa en melanoma con otros CPIs, incluyendo a PD-1, IDO-1 y CTLA-4 [33], se han desarrollado terapias combinadas que involucran el uso del bloqueo conjunto entre inmunoterapia con otros CPIs. El bloqueo conjunto de PD-1 y LAG-3 es el más ampliamente estudiado (Tabla 1) y ha mostrado mejores resultados en cuanto a la disminución del crecimiento tumoral en ensayos con ratones [71]. Por ejemplo, en ensayos de melanoma acral (metastásico), mostró una alta tasa de control de la enfermedad, mediante la activación de células T efectoras en el microambiente tumoral y una supervivencia libre de progresión (PFS por sus siglas en inglés) más duradera [75]. Estudios adicionales, mostraron que el uso de anticuerpos de doble bloqueo (que se dirigen a PD-L1 y LAG-3), permitieron el restablecimiento de la actividad prolongada de células T (en modelos murinos) y, la optimización de la respuesta inmune antitumoral mediante la secreción de IFN-ү e IL-2 [76-78].
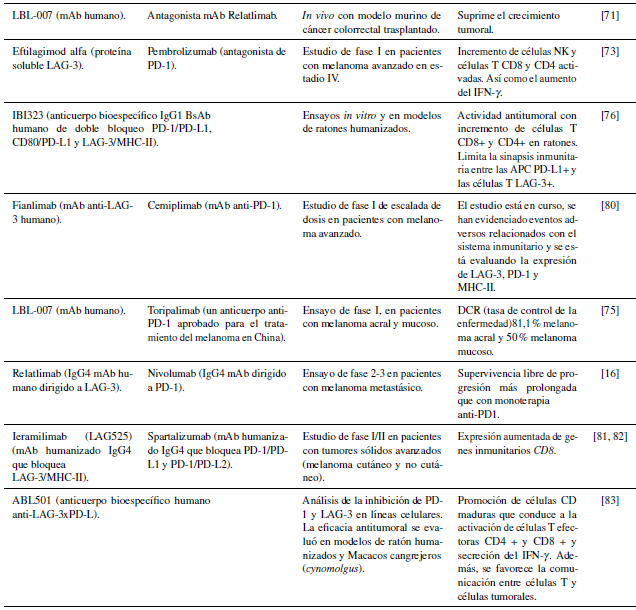
Es importante resaltar que la inhibición dual de PD-1 y LAG-3 mediante relatlimab y nivolumab respectivamente, ha sido la inmunoterapia de bloqueo conjunto más recientemente aprobada por la administración de alimentos y medicamentos de los Estados Unidos (FDA) para el tratamiento de personas mayores de 12 años con melanoma no resecable o metastásico [79]. De hecho, un reciente estudio reportó que relatlimab-nivolumab condujo a una PFS a los 12 meses (47,7% de los pacientes) más amplia que la monoterapia con nivolumab (36 % de los pacientes con una mediana de PFS de 4,6 meses). De igual forma, identificaron que la PFS fue más duradera en los pacientes que expresaron LAG-3 en 1 % o más [16]. Estos resultados son similares a los publicados en ensayos clínicos previos [79].
4 Inmunoterapia TIGIT
La inmunoterapia dirigida a TIGIT, se ha basado en el uso de monoterapia, terapia combinada y la combinación de anticuerpos-fármacos dirigidos a ligandos específicos [56, 84].
4.1 Monoterapia
La monoterapia ha sido dirigida al bloqueo de la unión de TIGIT a sus ligandos, mediante el uso de mAb. En melanoma el desarrollo clínico de mAb anti-TIGIT como monoterapia se encuentra en fases tempranas. Actualmente, se llevan a cabo estudios del anticuerpo anti-TIGIT vibostolimab (VIB), como monoterapia junto con el ensayo clínico NCT04335253 para evaluar el mAb anti-TIGIT EOS884448 (EOS-448). Este ensayo clínico corresponde a un estudio multicéntrico, abierto, de fase 1/2 en pacientes con tumores malignos sólidos avanzados previamente tratados. Como resultados preliminares de la fase 1 en un paciente, previamente tratado con un inhibidor de BRAF-MEK y pembrolizumab (PEM), se evidenció una reducción de los tumores intrabdominales del 50% [85]. Además, se ha empleado la desmetilación farmacológica de la línea celular de melanoma A375 como estrategia terapéutica reciente, en la cual se observó una reducción en la expresión de TIGIT constitutiva [86].
4.2 Terapia combinada
En estudios preclínicos ha sido evidenciado que la terapia conjunta dirigida al bloqueo de PD-1 y TI-GIT, presenta efectos favorables [13]. De hecho, ha sido mostrado que el bloqueo simultáneo de la vía PD-1 y la vía CD155-TIGIT, restablece la función efectora de las células T citotóxicas in vitro, lo cual fundamenta el uso de mAb combinados en el tratamiento de melanoma avanzado [64]. Sin embargo, al ser terapias recientes, la mayoría de ensayos clínicos se encuentran en fase de reclutamiento. Dentro de tales ensayos clínicos se encuentran:
4.2.1 Ensayo de fase I/II (NCT04305041)
Este ensayo busca analizar el efecto combinado de mAb de TIGIT (VIB) con mAb PD-1 (PEM) y anti-CTLA-4, Quavonlimab, (QUA) en 200 pacientes con melanoma refractario positivo para PD-1. Se estima obtener resultados preliminares de este ensayo clínico en el año 2030 [87].
4.2.2 Dos ensayos aleatorizados con 315 y 60pacientes con melanoma avanzado de primera línea (1L) y con melanoma en etapa III respectivamente
Este ensayo contempla evaluar la seguridad y eficacia del inhibidor P mediante el uso de mAb-antiPD-1, PEM, en combinación con agentes en investigación, incluyendo mAb dirigidos a TIGIT (VIB) [88, 89].
4.2.3 Ensayo clínico NCT05130177 de fase II del inhibidor de PD-1 zimberelimab (AB122) con inhibidor de TIGIT domvanalimab (AB154)
El objetivo de este ensayo es evaluar la desaparición completa y hasta en un 30% de eliminación de las lesiones del melanoma, junto con la supervivencia libre de progresión (PFS) y supervivencia general (OS) a los 6, 24,48 y 60 meses luego de iniciado el tratamiento, en un grupo de 26 pacientes con melanoma avanzado previamente tratados con inhibidores de PD-1. En este ensayo se tiene en cuenta la enfermedad progresiva en las 12 semanas posteriores a la última dosis de mAb anti-PD-1/L1 [90].
4.2.4 Ensayo clínico NCT05060432
Este ensayo corresponde a un estudio multicéntrico, abierto, de fase I/II de EOS884448 (EOS-448) en 376 pacientes de tumores sólidos como el melanoma. En este ensayo se pretende evaluar la respuesta a seis combinaciones de mAb de TIGIT, PD-1 y el antagonista del receptor de adenosina A2A [91].
5 Combinación de anticuerpos-fármacos dirigidos a ligandos específicos
Enfortumab vedotin, actúa como inhibidor de microtúbulos, y consiste en un anticuerpo monoclonal ligado a la monometil auristatina (MMAE) dirigido al bloqueo de Nectina-4. Los buenos resultados del uso de Enfortumab vedotin, han sido demostrados en carcinoma urotelial metastásico o localmente avanzado, en donde se observó gran eficacia antitumoral, evidenciada por un aumento de la supervivencia general libre de progresión en comparación con pacientes tratados con quimioterapia estándar e inhibidores de PD-1 y PD-L1 [92-94]. Estudios posteriores han recomendado el uso de Enfortumab vedotin en el tratamiento de melanoma avanzado [68], debido a la inducción de apoptosis en células de melanoma con mutación BRAF o resistentes a BRAFi (Figura 3).
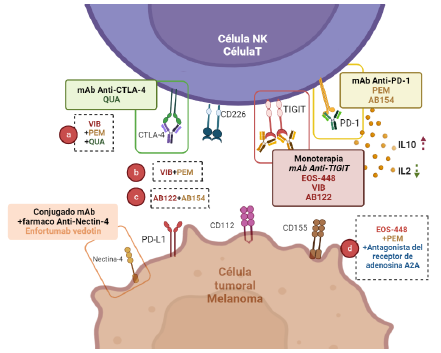
Figura 3 Ensayos clínicos de inmunoterapia con mAb anti- TIGIT, PD-1 y CTLA-4 en melanoma. Actualmente, los tratamientos usados en melanoma incluyen el uso de monoterapia y terapia combinada. En monoterapia se usan anticuerpos monoclonales anti-TIGIT como EOS-448, VIB y AB154. La terapia combinada incluye el uso conjunto de: a) VIB + PEM + QUA; b) VIB + PEM; c) AB122 + AB154; d) EOS-448 + PEM + antagonista del receptor de adenosina A2A. Adicionalmente, para el bloqueo de Nectina-4, se desarrollan ensayos clínicos de conjugados anticuerpo-fármaco como Enfortumab vedotin. VIB, vibostolimab; PEM, pembrolizumab; QUA, Quavonlimab; AB122, zimberelimab; AB154, domvanalimab.
6 Interacción entre LAG-3 y TIGIT en la respuesta inmune antitumoral
LAG-3 y TIGIT son moléculas coinhibitorias que tienen funciones reguladoras en la respuesta inmune del ambiente inflamatorio tumoral con patrones de expresión similares. Sin embargo, las colas de señalización de estos son únicas y definen las funciones específicas y los efectos sinérgicos de la terapia dirigida y combinada a estas moléculas en cáncer [95]. Se ha evidenciado que TIGIT se correlaciona con un aumento en la expresión de otros puntos de control inhibidores como LAG-3, TIM-3 y expresión disminuida de DNAM-1, lo cual reduce la capacidad del sistema inmune frente a las células del melanoma [96]. LAG-3, TIM-3 y TIGIT se expresan en gran medida en células T disfuncionales o agotadas y evidencian una correlación positiva con la producción de IL-10 [97].
Adicionalmente, ha sido indicado que TIGIT y LAG-3 tienen mayor expresión conjunta en la zona peritumoral que en el área intratumoral en células de melanoma cutáneo, lo cual ha sido relacionado con variables clinicopatológicas de peor pronóstico (mayor grosor de Breslow, aumento en la tasa de mitosis en el tumor y afectación ganglionar). Estos CPI se expresan en CD3+ y están correlacionados con la alta expresión de PD-1 y la densidad de TIL en el 30% de casos de melanoma, aunque dicha asociación no se presentó en células tumorales [98].
Recientemente, un estudio demostró que inhibir la glucógeno sintasa quinasa-3 (GSK-3) bloquea la transcripción de PD-1 y regula negativamente la expresión del gen de activación de linfocitos 3 (LAG-3) en células T CD4+ y CD8+ citolíticas para reducir el crecimiento tumoral de melanoma subcutáneo B16 en ratones. Ensayos posteriores en un paciente con melanoma metastásico demostró que Elraglusib un inhibidor de GSK-3, redujo la progresión del cáncer y disminuyó la expresión no solo de PD-1 y LAG-3, sino también de TIGIT en TIL CD8+ [99, 100]. Por lo anterior, TIGIT y LAG-3 se han convertido en biomarcadores pronósticos y posibles dianas de la inmunoterapia combinada antitumoral.
7 Conclusiones y perspectivas
TIGIT y LAG-3 son inhibidores de puntos de control inmunitarios que están regulados al alza en melanoma y cuya expresión sinérgica ha sido asociada con peor pronóstico de la enfermedad. Actualmente, mientras que el desarrollo clínico de monoterapias y terapias combinadas entre mAb anti-TIGIT con mAb anti-PD-1, y antagonistas del receptor de adenosina A2A se encuentra en etapas tempranas, la terapia de anticuerpos-fármacos como Enfortumab vedotin solo se encuentra reportada en ensayos preclínicos. Lo anterior evidencia la necesidad de la realización de estudios adicionales que permitan caracterizar el funcionamiento y la eficacia de terapias que han sido aprobadas para otros tipos de cáncer como el cáncer urotelial avanzado. Dada la alta implicación de LAG-3 y TIGIT/Nectina-4 en melanoma metastásico, estos receptores son actualmente considerados como blancos prometedores de terapia combinada con moléculas inhibitorias específicas como mAb anti-TIGIT y anti-LAG-3.

