










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.38 no.2 Santa Marta July/Dec. 2009
ENFERMEDADES CORALINAS Y SU INVESTIGACIÓN EN LOS ARRECIFES COLOMBIANOS*
CORAL DISEASES AND THEIR RESEARCH IN COLOMBIAN REEFS
Diego L. Gil-Agudelo1, Raúl Navas-Camacho1, Alberto Rodríguez-Ramírez2, María Catalina Reyes-Nivia3, Sonia Bejarano4, Jaime Garzón-Ferreira5 y Garriet W. Smith6
1Instituto de Investigaciones Marinas y Costeras-INVEMAR, Zona Portuaria, Cerro Punta Betín, Santa Marta, Colombia, diego.gil@invemar.org.co (D.L.G.A.), rnavas@invemar.org.co (R.N.C.)
2The University of Queensland, School of Earth Sciences, St. Lucia, Brisbane, QLD 4072, Australia, alberto.rodriguez@uq.edu.au
3The University of Queensland, Centre for Marine Studies, St. Lucia, Brisbane, QLD 4072, Australia. catalina.reyes@uq.edu.au
4University of Exeter. Marine Spatial Ecology Laboratory. Reino Unido, sb277@exeter.ac.uk
5Brewster Academy, 80 Academy Drive, Wolfeboro, NH, EE. UU. anisotremus@gmail.com
6University of South Carolina Aiken, Department of Biology and Geology, 471 University Parkway, Aiken, SC29801, EE. UU. smithres@usca.edu
RESUMEN
Los arrecifes coralinos, uno de los más bellos e importantes ecosistemas de nuestro planeta, han existido por más de 200 millones de años, sobreviviendo a grandes cambios durante su historia (glaciaciones, extinciones masivas, entre otros). Sin embargo, durante las últimas tres décadas se han registrado grandes y súbitos cambios en el medio marino que ajuicio de algunos investigadores ponen en peligro la supervivencia de corales y arrecifes coralinos. Uno de los mayores problemas que enfrentan los arrecifes coralinos hoy en día es la aparición de enfermedades que atacan a los corales, la base de estos ecosistemas. No existe un consenso sobre si estas enfermedades son realmente nuevas o no, pero lo que sí se sabe es que algunas de ellas están creando cambios en los arrecifes coralinos sin precedentes en más de cuatro mil años. Los arrecifes colombianos no son ajenos a la influencia de enfermedades. Desde la década de los años 80 varias de ellas han sido observadas y estudiadas e incluso una de ellas fue descrita por primera vez en áreas arrecifales de Colombia. Esta revisión presenta un compendio de las principales enfermedades coralinas observadas en el mundo con el fin de servir como guía para su identificación y como base para nuevos estudios en el tema. De igual forma, se presenta un resumen de los estudios en enfermedades coralinas que han sido llevados a cabo en Colombia y se discute sobre las perspectivas de investigación en este campo en el país.
PALABRAS CLAVES: Enfermedades coralinas, Arrecifes coralinos, Mortalidad, Colombia.
ABSTRACT
Coral reefs are one of the most beautiful and important ecosystems in the planet. These ecosystems have existed for over 200 million years and have survived extreme episodes such as glaciations and mass extinctions during their history. Nonetheless, during the last three decades, these ecosystems have registered sudden and dramatic changes that, according to some researchers, endanger their survival and persistence. One of the major problems coral reefs are facing nowadays is the outbreak of diseases that affect corals, which constitute the basic unit of this ecosystem. There is no consensus regarding whether these disease outbreaks are recent episodes; but what seems to be true is that some of these diseases have favored unprecedented changes in coral reefs. Coral reefs in Colombia have also been affected by disease events, and since the 1980's, several coral diseases have been observed and studied, and even one of them was first described in Colombian reefs. This work presents a compendium of the main coral diseases registered around the world and is meant to serve as a guide for new studies in this topic. Similarly, a summary of coral disease research carried out in Colombia is presented as well as a discussion on current perspectives for the study of this field in the country.
KEY WORDS: Coral diseases, Coral reefs, Mortality, Colombia.
INTRODUCCIÓN
Los corales escleractíneos (componentes principales de los arrecifes coralinos) aparecieron como grupo durante el Triásico (250 a 210 millones de años antes del presente). Durante este tiempo han experimentado períodos de gran diversidad (como por ejemplo durante el Jurásico), períodos de poca dominancia en el ambiente marino (por ejemplo desplazados por bivalvos rudistas durante el Cretáceo) y períodos de casi extinción (finales del Cretáceo y principios del Terciario) (Veron, 1995). En la actualidad, los arrecifes coralinos se encuentran distribuidos a lo largo de las costas tropicales del mundo, ocupando solamente el 1 % del ambiente marino (Birkeland, 1997), no obstante albergan una alta diversidad (calculada en más de nueve millones de especies sin contar microorganismos) y se constituyen en uno de los ecosistemas más altamente productivos del mundo (mayor que la de la mayoría de los cultivos que proveen alimento al hombre) (Dubinsky, 1990; Birkeland, 1997; Malakoff, 1997). Los arrecifes coralinos son igualmente importantes debido a sus significativas contribuciones en la economía mundial, aportando gran número de recursos (i.e. pesca) y servicios (i.e. turismo) y siendo la principal fuente de ingresos para numerosos países (Birkeland, 1997; Malakoff, 1997).
Debido a la estabilidad de la comunidad arrecifal y su cercanía al clímax ecológico, los arrecifes coralinos son ecosistemas muy frágiles ante la acción de agentes perturbadores externos; por esto existen muchas alteraciones, tanto de origen natural como antropogénico, que los afectan negativamente. Tormentas, huracanes, eventos como el fenómeno del Niño, contaminación, alta sedimentación por deforestación y sobrepesca, entre otros muchos factores, han provocado una considerable disminución de la salud coralina en las últimas décadas (Grigg y Dollar, 1990; Ginsburg, 1994; Birkeland, 1997). Sin embargo, uno de los fenómenos que parece estar afectándolos en mayor medida durante los últimos tiempos, es la aparición de nuevas y numerosas enfermedades coralinas. En algunos casos se cree que estas enfermedades han sido relativamente bien caracterizadas (como es el caso de la Aspergilosis y la Plaga Blanca), pero otras muchas han sido poco estudiadas hasta la fecha (como es el caso de la Banda Amarilla y la enfermedad de los Lunares Oscuros) a pesar de tener una gran incidencia en la mortalidad coralina de algunas áreas. Más aún, enfermedades que se creían relativamente bien caracterizadas, han mostrado nuevas facetas que revelan realmente lo poco que se ha progresado en estas tres décadas en el estudio de las enfermedades coralinas (Richardson, 1998; Frías-López et al, 2003; Sutherland et al, 2004).
Los arrecifes colombianos no han sido ajenos a esta problemática. Trabajos realizados desde hace varios años en Colombia han arrojado datos preocupantes sobre el grado de deterioro y la pérdida de la cobertura del tejido vivo coralino en sus arrecifes. Al igual que en otras partes del mundo, el retroceso de la salud en los arrecifes colombianos está causando alarma en la comunidad científica nacional. Dado el importante papel de las enfermedades coralinas en la dinámica de los arrecifes, se han implementado y desarrollado diferentes protocolos de monitoreo en los mares de Colombia y el mundo, incluyendo el Sistema Nacional de Monitoreo de Arrecifes Coralinos en Colombia (SIMAC) que desde 1998 ha monitoreado, entre otras variables, las enfermedades coralinas en los arrecifes coralinos del Caribe y Pacífico de Colombia (Garzón-Ferreira et al, 2002b).
Este trabajo incluye una síntesis de las enfermedades descritas hasta la fecha a nivel mundial (sin considerar el blanqueamiento no relacionado con procesos infecciosos). No se pretende reemplazar los trabajos en los que otros autores han realizado similares esfuerzos al compilar y revisar la información existente sobre enfermedades coralinas en el mundo (Richardson, 1998; Rosenberg y Loya, 2004; Sutherland et al, 2004, Weil et al, 2006), sino complementarlos con nuevos avances tanto en Colombia y en el mundo y resaltar la importancia de estas enfermedades en los ambientes arrecifales colombianos. De igual forma, se pretende proporcionar un marco de referencia para los investigadores interesados en generar nueva información sobre esta temática en nuestro país, a través de un análisis de las perspectivas de investigación, de manera que pueda asistir al diseño, desarrollo y ejecución de futuros estudios.
COMPENDIO DE LAS PRINCIPALES ENFERMEDADES CORALINAS REGISTRADAS EN EL MUNDO
A continuación se presenta una revisión de las enfermedades descritas en el ámbito mundial, incluyendo una descripción de las características para reconocimiento en campo de cada una, así como información etiológica. En la tabla 1 puede encontrarse una síntesis de los principales aspectos de cada tipo de enfermedad. Las abreviaturas utilizadas en el presente artículo se encuentran en español; pueden variar de aquellas usualmente utilizadas en idioma inglés.
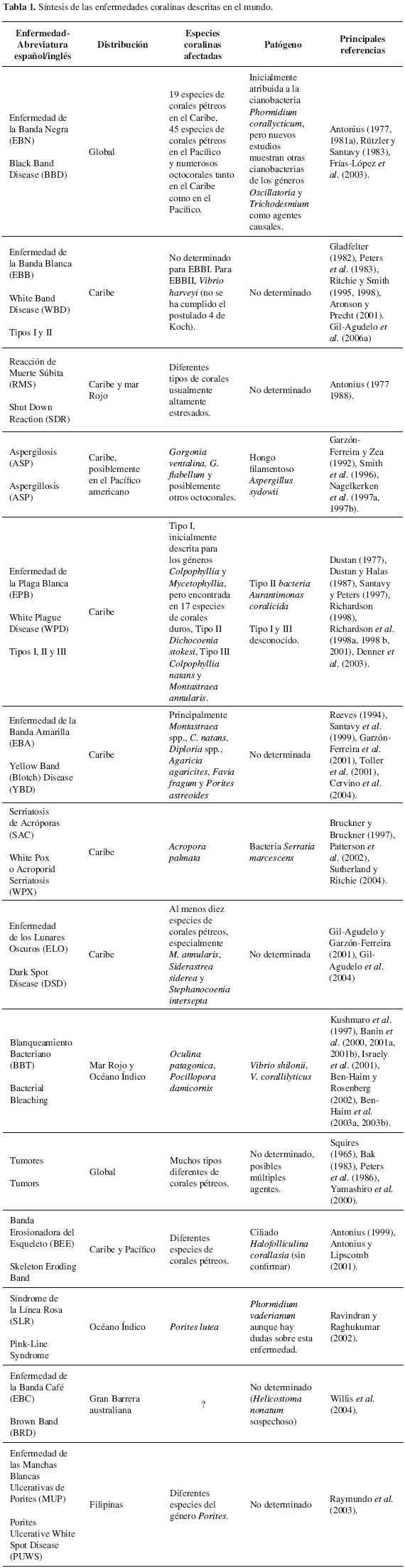
Enfermedad de la Banda Negra o Cyanobacterial (Microbial) Mat Disease (Black Band Disease) -EBN-
Observada y descrita por primera vez por Arnfried Antonius en 1973 en los arrecifes de Belice (Antonius, 1977, 1981a; Rützler et al, 1983) y complementada por Rützler y Santavy (1983). Esta afección se caracteriza por presentar una porción del esqueleto del coral desnudo, blanco y recién expuesto rodeado por una banda oscura de 5 a 30 mm de ancho que delimita el tejido sano (Figuras 1A, 1B) (Rützler y Santavy, 1983). Esta enfermedad fue detectada inicialmente en el mar Caribe, afectando principalmente a los corales pétreos de las especies Diploria strigosa y Montastraea annularis (Antonius, 1981b). En la actualidad al menos 19 especies de corales pétreos han sido observados con signos de esta enfermedad en el Caribe (Sutherland et al., 2004). La EBN ha sido también registrada en al menos 45 especies de corales pétreos del Indo-Pacífico y mar Rojo (Antonius, 1985b, 1988; Littler y Littler, 1996; Al-Moghrabi, 2001; Frías-López et al, 2002, 2003; Sutherland et al, 2004) y en numerosos octocorales tanto del Caribe como del Indo-Pacífico (Antonius, 1985a; Peters, 1993; Richardson et al, 1997; Sutherland et al, 2004).
Inicialmente Antonius (1981a, 1981b) identificó como componente principal de la EBN a la cianobacteria Oscillatoria submembranacea, posteriormente reclasificada como Phormidium corallyticum (Rützler y Santavy, 1983), la cual se creía que causaba necrosis en el tejido coralino por sus exudados (Peters, 1993). Estudios recientes, utilizando técnicas moleculares, han permitido establecer que esta enfermedad no es causada por un solo microorganismo sino que en la "banda" coexiste un consorcio microbiano constituido por al menos tres tipos de cianobacterias (Frías-López et al, 2003). Sin embargo, Frías-López et al. (2003) hallaron que, de las cianobacterias directamente relacionadas con la enfermedad, ninguna pertenece al género Phormidium, encontrando también que el consorcio de microorganismos asociados a la EBN en el Caribe se encuentra dominado por una especie desconocida de cianobacteria del género Oscillatoria, mientras que en el Pacífico es dominada por cianobacterias del género Trichodesmium. Aún se encuentra en discusión el papel de las cianobacterias en el proceso de infección de la enfermedad, así como el papel real de P. corallyticum en la misma. Otros microorganismos que también hacen parte del consorcio microbiano de la EBN son las bacterias oxidadoras de sulfuro Beggiatoa spp. (Ducklow y Mitchell, 1979), bacterias sulfato-reductoras Desulfovibrio spp. (Garrett y Ducklow, 1975), hongos, y otras bacterias heterotrófas (Ramos-Flores, 1983). Estudios realizados por Carlton y Richardson (1995) y Richardson et al (1997) muestran un cambio vertical abrupto en las concentraciones de oxígeno y sulfuro en la banda, alcanzando concentraciones de hasta 800 µM de sulfuro en la base de la banda a la vez que las concentraciones de oxígeno bajan prácticamente a cero, condiciones que se ha demostrado matan el coral. Richardson (1992) describió lo que llamó "enfermedad de la banda roja" en los cayos de la Florida, no obstante la investigación de esta enfermedad reveló como causante de la misma a dos cianobacterias del género Oscillatoria, por lo cual se considera como otro caso de la misma EBN. Por estas condiciones, se ha propuesto cambiar el nombre de la enfermedad a Cyanobacterial (Microbial) Mat Disease (Andrew Bruckner. NOAA. Washington D.C. 2009. Com. Pers.), ya que este nombre describe mejor la etiología de la enfermedad.
Se ha encontrado que la EBN no está distribuida por igual en todo el arrecife, ya que ha sido observada principalmente a poca profundidad (entre 0.5 y 4.0 m), avanzando a una tasa media de 3.1 mm/día (Rützler et al., 1983). También ha sido evidenciado un factor estacional en la aparición de la misma, avanzando más en algunas épocas del año que en otras (Rützler et al., 1983; Kuta y Richardson, 1996), al parecer por la existencia de una relación entre el crecimiento y la temperatura del agua (Kuta y Richardson, 1996). Ensayos en laboratorio muestran que la temperatura óptima de avance de la enfermedad es entre 28 y 32 oC (Rützler et al., 1983), deteniendo su desarrollo o muriendo por debajo de los 20 oC (Peters, 1993).
Enfermedad de la Banda Blanca (White Band Disease) -EBB-
Esta enfermedad, descrita inicialmente por Gladfelter (1982), ataca al parecer exclusivamente corales acropóridos (Acropora palmata y A. cervicornis) en el mar Caribe. El único registro de una afección similar fuera del Caribe corresponde a uno realizado por Antonius (1988) en el mar Rojo. Esta enfermedad se caracteriza por la presencia de una banda blanca de esqueleto recientemente desnudado de tejido, adyacente a un frente necrótico de tejido de aspecto normal (Figuras 1C y 1D). La banda blanca se origina en la base de las ramas y progresa hacia la parte superior de la colonia en una forma concéntrica (Gladfelter, 1982; Peters et al., 1983).
Ritchie y Smith (1995, 1998) encontraron colonias de A. cervicornis con una afección similar a EBB pero con ligeras diferencias en el modo de crecimiento, la cual llamaron enfermedad de la banda blanca tipo II. Ésta se caracteriza por la presencia de una banda de entre 2 y 20 cm de ancho de tejido vivo, pero blanqueado, entre el tejido de apariencia normal y el desnudado por la enfermedad (Ritchie y Smith, 1998). En algunas ocasiones la banda necrótica puede alcanzar el tejido sano, lo que hace imposible la diferenciación entre EBB tipo I y tipo II. La EBB tipo II progresa a una tasa que puede superar los 5 cm/día (Ritchie y Smith, 1998), mientras que la velocidad observada en la tipo I puede ser de tan solo 2 cm/día (Gladfelter, 1982).

Respecto a la identificación del organismo responsable de ocasionar la enfermedad, Peters et al. (1983) encontraron bacterias Gram negativas en el tejido de corales afectados por la EBB tipo I que no fueron encontrados en tejidos sanos. Sin embargo, estas agregaciones bacterianas no se han encontrado siempre asociadas con la enfermedad (Peters et al., 1983). Por esto, aún no se ha determinado el papel de tales bacterias en el proceso de la enfermedad (Sutherland et al., 2004). Ritchie y Smith (1995) estudiaron la composición bacteriana del mucus de corales con signos de EBB tipo II y encontraron que la bacteria Vibrio carchariae se encontraba presente en colonias enfermas pero no en las sanas, asignándola como posible causante de la enfermedad. No obstante, los postulados de Koch [1 el presunto patógeno debe estar presente en todos los casos de la enfermedad y ausente en organismos sanos; 2 el presunto patógeno debe ser aislado en un cultivo puro a partir de las lesiones de la enfermedad; 3 el presunto patógeno debe provocar los signos de la enfermedad en organismos susceptibles y sanos al ser inoculado; 4 el presunto patógeno debe reaislarse de las lesiones producidas en los organismos de experimentación (Madigan et al., 2003)] no fueron satisfechos durante este estudio, sin lo cual no se acepta su identificación como agente causal. Posteriormente, Gil-Agudelo et al. (2006a) identificaron a Vibrio harveyi (sinonimia de V. carchariae) como el agente causal de la enfermedad, reproduciendo por primera vez sus signos de desarrollo.
La incidencia de esta enfermedad en los arrecifes coralinos del mar Caribe ha sido muy amplia. Gladfelter (1982) la halló asociada con la muerte de cerca del 99 % de las poblaciones de corales del género Acropora en algunos arrecifes de la región, especialmente en St. Croix, Islas Vírgenes. De hecho, se atribuye a esta enfermedad la mortalidad masiva de estos corales a lo largo del Caribe y el subsiguiente cambio en las comunidades ecológicas de arrecifes someros a lo largo de toda la región (Aronson y Precht, 2001). Antes de la aparición de la EBB los arrecifes coralinos del Caribe presentaban una zonación característica. En la parte más somera del arrecife (cresta arrecifal entre 1 y 5 m de profundidad) se encontraba la zona de A. palmata y entre los 5-25 m se podía encontrar la de A. cervicornis, donde la especie dominaba el frente de arrecifes expuestos a oleaje (Goreau, 1959; Geister, 1977). Luego de la epizootia de banda blanca, los corales de estas franjas prácticamente desaparecieron (Hughes, 1994) y hoy se encuentran bajas coberturas de coral vivo, representadas básicamente por pequeñas colonias de Agaricia spp. y Porites astreoides. Estudios de paleo-reconstrucción de las barreras coralinas de Belice y las islas Vírgenes han mostrado que este fenómeno no tiene precedentes en aproximadamente 4000 años (Aronson y Precht, 2001). A pesar de los numerosos estudios realizados en EEB, no se ha podido establecer una relación entre su incidencia y factores de tipo antropogénico o natural (Gladfelter, 1982; Peters, 1984; Ritchie y Smith 1995, 1998).
Reacción de Muerte Súbita (Shut-Down Reaction) -RMS-
Antonius (1977) registró la denominada Muerte Súbita (Shut Down Reaction) en el Caribe, síndrome que provocaba la muerte rápida a corales altamente estresados, principalmente por condiciones ambientales extremas (tales como temperatura y sedimentación). La RMS comienza en cualquier punto de la colonia coralina a partir del cual se difunde hacia el resto de la colonia, despojando el tejido vivo a una tasa que puede alcanzar los 10 cm/hora. Se ha encontrado también que esta enfermedad es contagiosa, infectando a otros corales por contacto a través de la columna de agua, siempre y cuando se encuentren estresados (Antonius, 1977, 1988). La RMS ha sido registrada en corales del Caribe (Antonius, 1977) y del mar Rojo (Antonius, 1988), pero el agente causal no ha sido determinado (Sutherland et al, 2004).
Aspergilosis (Aspergillosis) -ASP-
Esta enfermedad hasta el momento sólo ha sido registrada afectando a los gorgonáceos Gorgonia flabellum y G. ventolina. Las lesiones ocasionadas por esta enfermedad se caracterizan por una regresión de la corteza (coenenquima) y una eventual exposición del esqueleto axial interno de estos abanicos de mar (Figuras 1E y 1F). Además, el área afectada se encuentra rodeada de un anillo color púrpura que indica la presencia de escleritos en abundancia y que podría ser un método de defensa (Smith et al, 1998). Durante la década de los ochenta se registraron mortalidades masivas de G. flabellum y G. ventolina en el Caribe sur que causaron la desaparición de más del 90 % de estos organismos en algunas zonas (Garzón-Ferreira y Zea, 1992). Hacia 1995 un nuevo evento de mortandad fue evidenciado y, aunque fue menos catastrófico, afectó los abanicos de mar en todo el mar Caribe (Nagelkerken et al, 1997a, 1997b).
Smith et al. (1996) identificaron como el patógeno responsable de esta enfermedad al hongo Aspergillus sydowii después de aislarlo de porciones enfermas de G. flabellum y G. ventolina y cumplir con todos los postulados de Koch. Las hifas del hongo se encuentran embebidas en el borde vivo de la lesión (Smith et al, 1996). Aspergillus sydowii es un hongo terrestre, poco común en ambientes marinos, inhabilitado para esporular pero capaz de germinar en el mar. Por esto, se ha discutido que la introducción de este hongo pudo darse a través de descargas de ríos (Smith et al, 1996). Si éste fuera el caso, podría indicar una conexión con perturbaciones de tipo antropogénico. De otro lado, Shinn et al (2000) propusieron que las esporas podrían estar siendo trasladadas desde el África sahariana por masas de viento y polvo. Al respecto, Weir-Brush et al (2004) aislaron esporas de A. sydowii de tormentas de polvo provenientes de África y que caían sobre las Islas Vírgenes e inocularon colonias sanas de G. ventalina, produciendo signos de ASP. Igualmente, realizaron experimentos similares con cepas de hongos de cultivos de referencia, las cuales no produjeron signos de la enfermedad. Aunque no son concluyentes, estos experimentos muestran la posibilidad que estas masas de polvo sean vectores de enfermedades.
Se ha encontrado que esta enfermedad es más frecuente en ambientes protegidos que en zonas de alta dinámica, donde se encuentra restringida a zonas más profundas del arrecife (Nagelkerken et al., 1997a, 1997b). También ha sido encontrada con mayor frecuencia en zonas de alta influencia antrópica (Kim y Harvell, 2002) y durante períodos cuando la temperatura del agua sobrepasa los 30 °C (Alker et al., 2001).
Autores como Kim et al. (2000a, 2000b), Dube et al. (2002) y Alker et al. (2004) han utilizado el modelo de infección A. sidowii-G. ventalina para estudiar la respuesta de corales en la creación de antibióticos y antifungales. De esta forma mostraron que los corales (en este caso corales gorgonáceos) producen sustancias químicas que reducen el daño en el tejido coralino e inhiben la acción del organismo patógeno y que estas sustancias pueden variar según el tamaño del organismo, encontrándose en mayores concentraciones en colonias pequeñas. Similarmente, Gil-Agudelo et al. (2006b) evaluaron el posible papel de bacterias asociadas a la capa de mucopolisacáridos de estos corales como posible determinante de la resistencia a patógenos. Ante esto, encontraron variaciones en las comunidades bacterianas de corales sanos y enfermos sugiriendo el posible papel protector ante enfermedades de las bacterias que viven en la capa de mucopolisacáridos.
Enfermedad de la Plaga Blanca (White Plague Disease) -EPB-
Dustan (1977) menciona esta enfermedad que luego fue denominada "White Plague" (Dustan y Halas, 1987) o plaga blanca (EPB) en corales de los géneros Colpophyllia y Mycetophyllia en arrecifes de Key Largo (Florida, EE. UU.). Esta enfermedad se caracteriza por el desprendimiento del tejido coralino que deja el esqueleto en perfectas condiciones (Santavy y Peters, 1997) y por una marcada línea entre el tejido sano y el esqueleto desnudo blanco (Figuras 1G y 1H) (Dustan, 1977). Posteriormente, con base en un brote de EPB en la Florida en 1995, se definió un nuevo tipo de EPB que se denominó "White Plague type II" -EPB tipo II- (Richardson et al., 1998a, 1988b). Las características para separar la EPB de los primeros informes (Dustan, 1977; Dustan y Halas, 1987) del tipo II, son las tasas de pérdida de tejido y las especies afectadas. Mientras las tasas inicialmente registradas no superaron los 3.1 mm/día, la EPB tipo II presentó tasas de hasta de 2 cm/día y afectó principalmente a Dichocoenia stokesi, una especie no mencionada como susceptible en los primeros registros (Richardson, 1998). En 1999, un diferente tipo de EPB fue encontrado en grandes colonias de M. annularis y C. natans, denudando tejido a una velocidad aun mayor que la encontrada en la EPB tipos I y II. A este síndrome se le denominó plaga blanca tipo III (Richardson et al., 2001), sin embargo, todavía no se han realizado estudios sobre esta aparente nueva enfermedad. En otras regiones diferentes al Caribe se han hecho observaciones de enfermedades en corales que muestran signos similares a los de EPB. Antonius (1985b) fue el primero en registrarla en el Indo-Pacífico, posteriormente Coles (1994) la registró en el golfo de Omán y más recientemente Riegl (2002) la registró para el golfo Arábigo. Sin embargo, en estos estudios se refieren a esta enfermedad como banda blanca.
De los tres tipos de EPB, sólo en el caso del tipo II se ha podido identificar el agente causal. Richardson et al. (1998a) aislaron cepas bacterianas a partir de colonias enfermas de D. stokesi en el evento de 1995 en la Florida (EE. UU.). Inicialmente, se identificó una bacteria cercana al género Sphingomonas, pero posteriores estudios mostraron que esta bacteria pertenecía a un nuevo género, y fue llamada Aurantimonas coralicida (Denner et al., 2003). Adicionalmente, el alga calcárea Halimeda opuntia fue identificada por Nugues et al. (2004) como posible vector y reservorio de la EPB tipo II. De otro lado, ensayos recientes utilizando marcadores fluorescentes no han mostrado la presencia de A. coralicida en muestras de corales afectados por EPB tipo I, lo cual es un indicativo que son enfermedades diferentes (Laurie Richardson. Florida International University. Miami, EE. UU. 2007. Com. Pers.).
Enfermedad de la Banda Amarilla (Yellow Band/Blotch Disease) -EBA-
Esta enfermedad se caracteriza por la presencia de parches circulares o irregulares de tejido de color amarillo brillante translúcido que pueden volverse bandas (Figuras 2A y 2B). El hecho que el tejido se torne amarillo la diferencia del palidecimiento o blanqueamiento coralino (Santavy et al., 1999). La EBA fue observada por primera vez afectando corales de la especie Montastraea faveolata en los cayos de la Florida (Reeves, 1994). Hacia 1996, un extenso brote de la enfermedad fue observado en las islas de San Blas, Panamá, nuevamente en corales de la misma especie. Al mismo tiempo, similares signos de esta enfermedad fueron observados en otros lugares del Gran Caribe como Key West, Jamaica, Cuba y Honduras, en diferentes especies de Montastraea (Santavy et al., 1999). Garzón-Ferreira et al. (2001), durante sus estudios en arrecifes colombianos, ampliaron el ámbito de especies que son afectadas por esta enfermedad, incluyendo C. natans, Diploria spp., Agaricia agaricites, Faviafragum y Porites astreoides. Recientemente James Cervino (University of South Carolina. Columbia, EE. UU. 2007. Com. Per.) observó signos similares a los de esta enfermedad en arrecifes del Pacífico (Bali, Indonesia). El crecimiento de la EBA ha sido determinado únicamente para M. faveolata, encontrando tasas de avance cercanas a 0.70 cm/mes (Gil-Agudelo et al., 2004).
Cervino et al. (2001) encontraron una reducción entre 41 y 97 % en el número de zooxantelas en corales con signos de banda amarilla. También estas zooxantelas tenían una significativa reducción en su índice mitótico llegando casi al 0 % y parecían vacuoladas. Por su parte, Toller et al. (2001) encontraron que las zooxantelas en el tejido afectado por esta enfermedad eran Symbiodinium pertenecientes a los clados A y E, en comparación con los clados B y C que usualmente dominan en las especies estudiadas (complejo M. annularis) en la zona. Esto explicaría el color amarillo de la banda (color de las zooxantelas del clado A) además que la presencia de estos clados sería la respuesta a condiciones de estrés que afectan a los simbiontes sensibles. Debido a estas observaciones, actualmente se cree que la EBA podría ser una enfermedad de las zooxantelas y no del coral (Cervino et al., 2001; Gil-Agudelo et al., 2004). Cervino et al. (2004) aislaron en laboratorio diferentes cepas bacterianas del género Vibrio sugiriendo que éstas podrían ser las responsables de producir la enfermedad, no obstante sus resultados no fueron concluyentes.
Korrubel y Riegl (1998) y Riegl (2002) registraron una enfermedad en el golfo de Omán que también denominaron banda amarilla. Sin embargo, esta afección es diferente a la encontrada en el Caribe debido a la presencia de una capa microbiana de color amarillo sobre el coral. Esta capa migra sobre la supericie del tejido coralino matándolo a su paso y dejando el esqueleto de color amarillo (Sutherland et al., 2004). La tasa de crecimiento de esta enfermedad se incrementa durante los meses de verano, alcanzando más de 19 mm/semana. También se ha encontrado que es transmisible entre colonias coralinas (Riegl, 2002). Las especies coralinas más afectadas son Acropora clathrata, A. pharaonis, A. tenuis, A. valida, A. florida, Porites lutea, P. lichen, P. nodifera, Turbidina reniformis y Cyphastrea microphthalma (Korrubel y Riegl, 1998).
Serratiosis de Acróporas (White Pox o Acroporid Serratiosis) -SAC-
Observada por primera vez en los cayos de la Florida (Holden, 1996), esta enfermedad se ha encontrado afectando únicamente corales de la especie A. palmata. Conocida comúnmente como white pox disease, también ha sido llamada patchy necrosis (Bruckner y Bruckner, 1997). Las lesiones ocasionadas por la SAC se distinguen por ser parches blancos irregulares de esqueleto recién expuesto, rodeados por un frente necrótico de tejido con aspecto y coloración aparentemente normal (Figuras 2C y 2D) (Sutherland et al., 2004). Las lesiones pueden ser pequeñas (algunos cm2) hasta 80 cm2 o más y pueden encontrarse varias lesiones en la misma colonia (Figura 2C) (Patterson et al., 2002). Se ha encontrado que esta enfermedad se desarrolla con mayor rapidez y es más prevalente en épocas de aguas cálidas y de lluvias abundantes (Patterson et al., 2002; Sutherland y Ritchie, 2004). La SAC ha sido relacionada con la mortandad masiva de corales A. palmata en el Santuario Natural de los Cayos de la Florida (Patterson et al., 2002). Entre los años de 1996 y 2002, se calculó la pérdida de A. palmata en los cayos de la Florida en un 87 %, alcanzando el 97 % en algunos lugares, debido principalmente a la incidencia de esta enfermedad (Patterson et al., 2002; Sutherland y Ritchie, 2004).
A esta enfermedad se le cambió el nombre de "white pox" a SAC (acroporid serratiosis) debido a que la bacteria Serraría marcescens fue determinada como el agente causal (Patterson et al, 2002); S. marcescens es una enterobacteria común en excrementos humanos y animales, en suelos y agua. Su prevalencia en el ambiente marino es desconocida, pero ha sido hallada en aguas marinas contaminadas con aguas residuales (Sutherland et al., 2004). De igual forma, S. marcescens ha sido determinada como agente patógeno en otros animales (vacas, cabras, peces, insectos) y plantas, y es un patógeno oportunista en humanos (Sutherland y Ritchie, 2004). En la actualidad se realizan estudios para conocer la fuente y los vectores de la enfermedad. Inicialmente, S. marcescens ha sido encontrada en las heces del pez Sparisoma viride, el cual podría ser un vector de la enfermedad al defecar sobre las colonias de A. palmata (Bruckner, 2002). Estudios preliminares han encontrado este mismo tipo bacteriano en las aguas residuales de la ciudad de Key Largo (Florida), (Kathryn P. Sutherland. Rollins College. Winter Park, EE. UU. 2007. Com. Per.). De comprobarse la relación entre los aportes de aguas residuales al medio marino y el desarrollo de la enfermedad, sería el primer caso demostrado de un vínculo directo entre contaminación humana y el desarrollo de una enfermedad coralina (Sutherland y Ritchie, 2004).
Enfermedad de los Lunares Oscuros (Dark Spots Disease) -ELO-
Esta enfermedad fue observada por primera vez en el archipiélago de Nuestra Señora del Rosario (Caribe colombiano) hacia 1991. Inicialmente fue descrita como "medallones mostaza" y se registró como un tipo de blanqueamiento (Solano et al., 1993). En 1994, fue nuevamente observada pero esta vez en el archipiélago de San Andrés y Providencia y fue llamada "enfermedad de los lunares oscuros" (ELO). Estas observaciones iniciales fueron realizadas en los corales del complejo M. annularis. Meses después, signos similares fueron observados en colonias de Siderastrea sidérea en el golfo de Urabá, Caribe colombiano (Garzón-Ferreira y Gil, 1998; Gil-Agudelo et al., 2004). Hoy en día esta enfermedad ha sido registrada a todo lo largo del mar Caribe (Goreau et al., 1998; Gil-Agudelo et al., 2004; Weil, 2004).
La ELO se caracteriza por la presencia de manchas redondas más oscuras que el tejido normal (de colores púrpura, marrón o café), que crecen en tamaño con el tiempo, presentando algunas de ellas depresión en el tejido y esqueleto coralino (Figuras 2E y 2F). Cuando se presenta tejido muerto al interior del lunar, el tejido afectado parece un anillo rodeando la porción muerta (Garzón-Ferreira y Gil, 1998; Goreau et al., 1998; Gil-Agudelo y Garzón-Ferreira 2001; Gil-Agudelo et al., 2004). Se ha encontrado que esta enfermedad afecta principalmente a corales de las especies S. siderea y M. annularis, pero signos similares han sido registrados en al menos otras 10 especies coralinas del Caribe (Garzón-Ferreira et al., 2001). Weil (2004) ha calculado que la prevalencia de esta enfermedad en corales del Caribe es de aproximadamente 1.2 % y esto la hace una de las tres enfermedades más prevalentes en la región, superada solamente por la Enfermedad de Banda Negra y Aspergillosis. Su prevalencia parece incrementarse con el aumento de la temperatura del agua (Gil-Agudelo y Garzón-Ferreira, 2001).
Se han realizado estudios microbiológicos con el fin de detectar el agente responsable de esta enfermedad. Como resultado, se han encontrado cambios en la composición microbiana del mucus de corales enfermos, y se han hallado bacterias posiblemente oportunistas como V. carchariae viviendo solamente en colonias enfermas. Aún no se ha podido inducir la enfermedad en corales sanos, para completar así los postulados de Koch (Gil-Agudelo et al., 2004, 2007; datos sin publicar). También se han encontrado hongos endolíticos relacionados con colonias con signos de lunares oscuros. Una nueva hipótesis señala a estos hongos como posibles responsables de la enfermedad (Obs. Pers.; Galloway et al., 2007).
Recientemente, Work et al. (2008) encontraron signos de una enfermedad similar a ELO en corales de las especies Pavona varians, P. maldivensis, Psamocora nierstrazi y Montipora sp. en Hawai y Samoa estadounidense en el óceano Pacífico. Análisis histopatológicos de las lesiones mostraron hongos endolíticos asociados, similar a los encontrados en corales con ELO en el mar Caribe.
Blanqueamiento Bacteriano (Bacterial Bleaching) -BBT-
El blanqueamiento coralino ha sido ligado usualmente a cambios en factores físicos o químicos del ambiente y no a factores bióticos. Sin embargo, Kushmaro et al. (1997) demostraron que el coral Oculina patagonica se blanqueaba debido a la acción de la bacteria Vibrio shilonii (V. shiloi, sic) en el mar Mediterráneo, siendo la primera investigación que relaciona el blanqueamiento con un agente biológico. Posteriormente, se ha argumentado que V. shilonii es una sinonimia de V. mediterranei (Thompson et al., 2001).

Esta enfermedad es posiblemente la mejor estudiada hasta la fecha desde el punto de vista de patogénesis. Se sabe que V. shilonii es atraída por el mucus de O. patagonica (Banin et al., 2001a; Rosenberg, 2004). Luego, la bacteria se adhiere a receptores en la superficie del coral que contienen p-galactosidasa. Un aspecto interesante es que la adhesión de la bacteria a la superficie del coral sólo se lleva a cabo cuando V. shilonii es cultivada a temperaturas elevadas (25 a 28 oC) (Toren et al., 1998; Rosenberg, 2004). Después, la bacteria penetra el tejido coralino y se multiplica, pasando a un estado viable pero no cultivable (Banin et al., 2000). A partir de este punto no se conoce mucho sobre lo que sucede, pero se sabe que V. shilonii produce toxinas extracelulares que bloquean la fotosíntesis de las zooxantelas. Una de estas toxinas es la llamada toxina P, debido a la abundancia de prolina en el péptido. Se cree que la toxina se adhiere a la membrana de las zooxantelas haciendo canales que permiten el ingreso de amonio a éstas, destruyendo su gradiente de pH y bloqueando la fotosíntesis (Banin et al., 2001b; Rosenberg, 2004). Recientemente se ha encontrado que las toxinas de V. shilonii, que producen daño en el aparato fotosintético de zooxantelas en O. patagonica, no tienen el mismo efecto en zooxantelas de otros corales (especialmente aquellos del Caribe). Esto podría mostrar que esta especie bacteriana no sería responsable de blanqueamiento masivo, el cual sucede en grandes áreas e involucra numerosas especies coralinas (observaciones recientes y datos sin publicar, Rosenberg, 2004) y el cual se ha atribuido a condiciones ambientales y de estrés.
La incidencia del blanqueamiento coralino puede llegar al 90 % de las colonias de O. patagonica durante los meses de verano (temperatura del agua cercana a los 30 oC) en el mar Mediterráneo, disminuyendo al 5 % o menos cuando la temperatura del agua se encuentra por debajo de 20 oC (Israely et al., 2001; Rosenberg, 2004). Se ha establecido también que durante el invierno no es posible encontrar a V. shilonii en colonias de O. patagonica, sin embargo, sí en abundancia en el gusano de fuego Hermodice carunculata, el cual serviría como reservorio durante esta época y posiblemente también como vector de la enfermedad (Israely et al., 2001; Rosenberg, 2004).
Ben-Haim y Rosenberg (2002) y Ben-Haim et al. (2003a) identificaron una nueva especie de Vibrio (V. coralliilyticus) produciendo signos similares a los del blanqueamiento bacteriano en corales de la especie Pocillopora damicornis en Zanzíbar, océano Indico. Además de producir blanqueamiento, V. coralliilyticus produce lisis del tejido coralino a temperaturas superiores a los 27 oC (Ben-Haim et al., 2003b). Al igual que V. shilonii, V. coralliilyticus produce toxinas extracelulares. Sin embargo el papel de estas toxinas, su modo de acción y sus características aún son desconocidas (Ben-Haim et al., 2003b). No se han realizado estudios de la incidencia de esta enfermedad en los arrecifes de ésta y otras regiones, pero se sabe que el aumento en la temperatura del agua incrementa la virulencia del patógeno o la susceptibilidad del hospedero (Ben-Haim y Rosenberg, 2002).
Debido al hallazgo de bacterias que producen signos similares a los de blanqueamiento coralino, Rosenberg y Ben-Haim (2002) y Rosenberg (2004) formularon la llamada hipótesis de bacterias como productoras de blanqueamiento coralino. En ésta, se establece la posibilidad que el blanqueamiento coralino sea producido por bacterias, debido a que éstas liberan toxinas como respuesta a cambios ambientales, tales como aumento de la temperatura del agua. De esta forma, el blanqueamiento si bien parece ser producido por incrementos en la temperatura, su causa real es la enfermedad bacteriana.
Tumores
Los tumores en corales han sido observados por varias décadas en corales. Posiblemente el primero de estos registros es el de Squires (1965) en colonias del coral Madrepora kauaiensis colectadas en las islas de Hawai en 1902. Estos tumores son formados por una proliferación anormal del tejido coralino y usualmente se encuentran asociados a formaciones anormales del esqueleto (Figuras 2G y 2H) (Peters etal., 1986; Sutherland et al., 2004). Los tumores se caracterizan por presentar un adelgazamiento del tejido coralino que recubre la parte afectada, incremento en la porosidad del esqueleto, pérdida de células secretoras de mucus y nematocistos, disminución o ausencia de zooxantelas, reducción de la fecundidad de la colonia y, más notorio, pérdida o degeneración de los pólipos (Bak, 1983; Peters et al., 1986; Yamashiro et al., 2000).
Los tumores han sido registrados en una gran variedad de corales pétreos y octocorales en arrecifes alrededor del mundo (Peters et al., 1986; Sutherland et al, 2004). Gateño et al. (2003) encontraron que los tumores poseen una menor densidad y mayor contenido de magnesio en sus esqueletos, favoreciendo la entrada de organismos erodadores. Estas anomalías pueden ser el resultado de interacciones con otros organismos como crustáceos (Grygier y Cairns, 1996), tremátodos (Aeby, 1998) y otros (Sutherland et al., 2004) o verdaderos tumores formados por simple multiplicación incontrolada de células (Peters et al., 1986).
A pesar de que muchos consideran el término neoplasia y tumores como sinónimos, los tumores pueden formarse por neoplasia (proliferación celular incontrolada), hiperplasia (proliferación celular de tejidos normales) o hipertrofia (incremento del tamaño de las células de tejidos normales) (Sutherland et al., 2004). Por su parte, Gateño et al. (2003) propone utilizar el término "Tumor del Esqueleto" (Skeletal Tumor) o Epitelioma Calicoblástico (Calicoblastic Epithelioma) por considerar que los nombres anteriormente mencionados no reflejan la patogénesis y etiología de este fenómeno.
Banda Erosionadora del Esqueleto (Skeleton Eroding Band) -BEE-
Observada por primera vez por Antonius (1999), esta enfermedad ha sido registrada en el mar Rojo, el océano Indico y la Gran Barrera australiana. Se caracteriza por una delgada banda de color negro, muy similar a la banda negra que avanza consumiendo el tejido coralino y dejando marcas de erosión en el esqueleto desnudo. Sin embargo, se cree que esta enfermedad es causada por el protozoario Halofolliculina corallasia (Antonius, 1999; Antonius y Lipscomb, 2001) a pesar de que los postulados de Koch aún no han sido satisfechos. Grupos de estos ciliados producen daño físico al tejido y al esqueleto con sus cilios, embebiendo la parte basal de su lorica en el tejido del coral; se cree que este proceso se ve favorecido por la acción de sustancias químicas secretadas por los protozoarios (Antonius y Lipscomb, 2001). Recientemente, se han observado signos similares de esta enfermedad en corales de la costa Caribe venezolana (Aldo Croquer. Universidad de Puerto Rico. Mayagüez, Puerto Rico. 2008. Com. Pers.) y Puerto Rico (obs. pers.). El avance de la banda erosionadora del esqueleto se da por la producción asexual de larvas de ciliados que migran para buscar un sitio para asentarse y secretar su lorica. Se ha calculado que este avance es de solamente 1 mm/semana, pero puede llegar a 1 mm/ día (Antonius y Lipscomb, 2001).
Síndrome de la Línea Rosa (Pink-Line Syndrome) -SLR-
Registrada por primera vez en el océano Indico en 2002, esta enfermedad que ataca el coral Porites lutea se caracteriza por una línea o banda de tejido de color rosa que separa el tejido sano del esqueleto donde el tejido ha muerto. Se ha encontrado que la cianobacteria Phormidium valderianum produce los signos característicos de esta enfermedad, satisfaciendo así los postulados de Koch. Sin embargo, esta enfermedad no forma una capa de cianobacterias que mata el tejido que se encuentra bajo éste (como en el caso de la banda negra). La coloración de esta enfermedad se debe, al parecer, a una reacción del tejido a la presencia de la cianobacteria (Ravindran y Raghukumar, 2002).
Se desconoce la incidencia de esta enfermedad en los arrecifes del Pacífico, pero Ravindran y Raghukumar (2002) encontraron cerca del 10 % de las colonias de P. lutea con signos de SLR en el arrecife de Kavaratti (India). Willis et al. (2004) argumentan que esta misma coloración se presenta en este coral como respuesta a variadas formas de estrés, tales como depredación, competencia, invasión por parásitos, entre otras, lo cual mostraría que esta línea no puede relacionarse con un único agente causal o patógeno.
Enfermedad de la Banda Café (Brown Band Disease) -EBC-
Esta enfermedad fue detectada por primera vez en el verano de 2003 en la Gran Barrera arrecifal australiana. La EBC presenta una zona de color marrón, de grosor variable, bordeada de tejido sano en el frente de progresión y esqueleto expuesto en la parte posterior. Ocasionalmente se encuentra una zona blanca entre el tejido sano y la banda café, la cual se puede deber a tejido blanqueado o esqueleto desnudo (Willis et al., 2004). Grandes densidades de organismos ciliados se han encontrado en esta banda, los cuales consumen las zooxantelas del coral y le dan la coloración café a la misma. Se cree que esta enfermedad es ocasionada por el ciliado Helicostoma nonatum, sin embargo aún no se han realizado experimentos para su comprobación (Willis et al., 2004).
Enfermedad de las manchas blancas ulcerativas en Porites (Pontes ulcerative white spot disease) -MUP-
Se observó por primera vez en las Filipinas a finales de los años 90. La enfermedad, que ataca a corales del género Porites comienza como focos de tejido blanqueado pero intacto, de forma redonda u ovalada y bien delimitados. Estos focos pueden llegar a formar úlceras en el tejido coralino, los cuales pueden unirse con otros focos. Ocasionalmente, la enfermedad produce la muerte de toda la colonia coralina (Raymundo et al., 2003). La incidencia de esta enfermedad en los arrecifes de las Filipinas es alta. Raymundo et al. (2003) encontraron la enfermedad en 80 % de los arrecifes muestreados, afectando a aproximadamente un 22 % de las colonias de Porites. En 2002, Longin Kaczmarsky (Florida International University. Miami. 2007 Com. Per.) encontró más del 57 % de las colonias de Porites spp. afectadas por la enfermedad. La tasa de progresión de la enfermedad parece ser lenta, con colonias coralinas mostrando lesiones por largo tiempo sin mostrar pérdida significativa de tejido. Sin embargo, la recuperación de la enfermedad ha sido raramente observada (Raymundo et al., 2003).
Experimentos de transmisión de la enfermedad realizados en campo mostraron que el 95 % de colonias sanas puestas en contacto con corales afectados por la enfermedad, presentaron lesiones luego de cinco semanas (Raymundo et al., 2003). Aunque no se han realizado estudios extensivos, parece haber una relación entre temperatura y aparición de la enfermedad, encontrándose mayor prevalencia de ésta por encima de los 26 °C (Raymundo et al., 2003).
INVESTIGACIÓN EN ENFERMEDADES DE CORALES EN COLOMBIA.
Historia
Antes de la década de los años 90, la investigación en corales y arrecifes coralinos en Colombia se enfocó principalmente en establecer una línea base de la composición, estructura y abundancia coralina de los arrecifes colombianos. Como ejemplo de este tipo de estudios están los trabajos de Erhardt y Werding (1975a, 1975b), Prahl y Erhardt (1985), Werding y Sánchez (1988, 1989), entre muchos otros. Sin embargo, sólo hasta 2000 se puede encontrar un estudio compendio con la línea base de todas las áreas coralinas de Colombia (Díaz et al., 2000). Con base en dichos estudios y debido al evidente deterioro sufrido por los arrecifes colombianos durante la década de los años 80 y principios de los años 90, se establecieron iniciativas para el monitoreo de las áreas arrecifales en el país. La primera de éstas fue la vinculación del país al programa CARICOMP (Caribbean Coastal Marine Productivity Program) y, luego en 1998, se inició el desarrollo del Sistema Nacional de Monitoreo de Arrecifes Coralinos en Colombia -SIMAC- (Garzón-Ferreira, 1999), un programa interinstitucional liderado por el Instituto de Investigaciones Marinas y Costeras (INVEMAR) y con el cual se implemento un protocolo específico para el seguimiento temporal de las enfermedades coralinas.
No obstante, antes del SIMAC a principios de los años 90, se realizaron evaluaciones enfocadas a la descripción de eventos específicos de enfermedades. Quizás el primero de estos estudios fue el de Garzón-Ferreira y Zea (1992), en el cual se describió la mortandad masiva del octocoral Gorgonia ventalina en el área de Santa Marta. Luego en 1993 se publicó un estudio por Solano et al. (1993) sobre la incidencia de blanqueamiento coralino en el archipiélago de Nuestra Señora del Rosario. Posteriormente, Díaz et al. (1995) publicaron el primer estudio en el que se evaluó salud arrecifal en Colombia y en el que se distinguieron claramente algunas enfermedades coralinas como factores de deterioro arrecifal. En ese estudio se acuñó el nombre de "Enfermedad de los Lunares Oscuros ELO (Dark Spots Disease)", a pesar que Solano et al. (1993) había descrito signos similares a esta enfermedad como "medallones mostaza" durante su estudio de blanqueamiento. Se convierte así esta enfermedad como la primera y única enfermedad coralina descrita en Colombia hasta el momento. Continuando con el estudio de la ELO, Gil-Agudelo y Garzón-Ferreira (2001) describieron la distribución e incidencia de esta enfermedad en los arrecifes del Parque Nacional Natural Tayrona, Santa Marta, y Garcés (2000) investigó el desarrollo de esta misma enfermedad en dos especies coralinas en la misma área. Finalmente, Gil-Agudelo et al. (2004) realizaron las primeras aproximaciones para establecer el patógeno causante de esta enfermedad (ver ELO en sección anterior).
También se han realizado trabajos sobre la incidencia de diferentes enfermedades coralinas en diversos arrecifes del país, como el Barrios (2000), en el cual se evaluaron conjuntamente las enfermedades y los principales agentes de deterioro de corales en el Caribe colombiano y el de Garzón-Ferreira et al. (2001), que registró enfermedades entre 1992 y 1998 en 13 arrecifes caribeños. Por su parte, Garzón-Ferreira y Pinzón (1999) realizaron los primeros registros de enfermedades coralinas para el Pacífico oriental en la isla de Malpelo, encontrando un síndrome similar a la EBB sobre corales del género Pocillopora y otro semejante a la EPB en Porites lobata. Además para el Pacífico colombiano, Zapata et al. (2001) hicieron la recopilación sobre el estado de salud de los arrecifes de la Isla Gorgona.
Monitoreo de enfermedades coralinas en Colombia en el marco del programa SIMAC
Con la creación del SIMAC, se comenzó con la evaluación y monitoreo de las diferentes enfermedades coralinas descritas, su incidencia, prevalencia y evolución en los arrecifes coralinos de cinco localidades del Caribe y tres en la costa pacífica colombiana (Garzón-Ferreira et al, 2002b). Tres de las estaciones del Caribe (isla de San Andrés, islas del Rosario y bahía de Chengue) han sido monitoreadas desde 1998. Las dos estaciones restantes (islas de San Bernardo y Urabá chocoano) se instalaron en 2002. En el Pacífico, la estación de la isla Gorgona fue establecida en 1998 y las otras dos (Utría y Malpelo) fueron instaladas en 2002 y 2003 respectivamente.
Durante el monitoreo SIMAC han sido observadas ocho enfermedades afectando los corales del Caribe colombiano: Plaga Blanca (EPB), Lunares Oscuros (ELO), Banda Negra (EBN), Banda Amarilla (EBA), Banda Blanca (EBB), Banda Roja (EBR), Reacción de Muerte Súbita (RMS) y la Serratiosis de Acropóridos (SAC). De éstas, la EMS y la SAC no habían sido registradas anteriormente (cf. Garzón-Ferreira et al., 2001). Para las áreas de monitoreo del Pacífico, se han encontrado colonias aisladas con síndromes similares a la EPB y la EBB descritos por Garzón-Ferreira y Pinzón (1999). Más recientemente, gorgonáceos del género Pacifigorgia fueron observados con signos de una enfermedad similar a la aspergillosis en la ensenada de Utría, durante la campaña de monitoreo de 2002. Por lo tanto, investigaciones conducentes para corroborar la presencia de enfermedades o patógenos asociados a la mortalidad de corales pétreos o blandos en los arrecifes del Pacífico colombiano deben ser implementados en el corto plazo.
Entre 1998 y 2005 en el SIMAC se han registrado diferentes especies de corales pétreos padeciendo enfermedades, de las cuales once no habían sido registradas por Garzón-Ferreira et al. (2001) e incrementaron a 36 el número total de especies afectadas por al menos una enfermedad en los arrecifes del Caribe colombiano (Tabla 2). Además, para varias especies se han detectado nuevas afecciones, siendo las de mayor susceptibilidad (con cuatro o cinco enfermedades): A. palmata, A. agaricites, C. natans, D. strigosa y las especies del complejo Montastraea annularis (Tabla 2). La EPB y la ELO son las enfermedades que más especies han afectado con 34 y 22 especies respectivamente (Tabla 2) y en conjunto son las responsables de la mayor prevalencia de enfermedades, así como las únicas registradas en todas las localidades de monitoreo (Garzón-Ferreira et al, 2004). Previamente se habían registrado sólo alrededor de 20 especies coralinas afectadas por EPB (Richardson, 1998; Richardson et al, 1998b; Gil-Agudelo et al, 2004) y hasta diez con ELO (Garzón-Ferreira et al, 2001; Gil-Agudelo et al, 2004). De otro lado, a lo largo del período de seguimiento (1998-2005) la prevalencia de enfermedades ha variado entre un máximo de 8.6 % (San Andrés 1999) y un mínimo de 0.6 % (islas del Rosario 2005 y bahía de Chengue 2000 y 2005), con cambios interanuales de diferente magnitud en cada localidad, pero tendiendo a presentar los mayores niveles en las estaciones de monitoreo de la isla de San Andrés (Figura 3). En relación con este hecho, desde hace algunos años San Andrés ha sido considerada como un centro de alta prevalencia de enfermedades coralinas (Weil et al, 2002), sin embargo no hay claridad sobre cuales son los factores que han contribuido a ello.
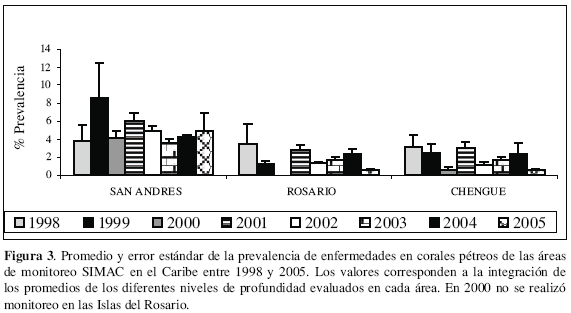
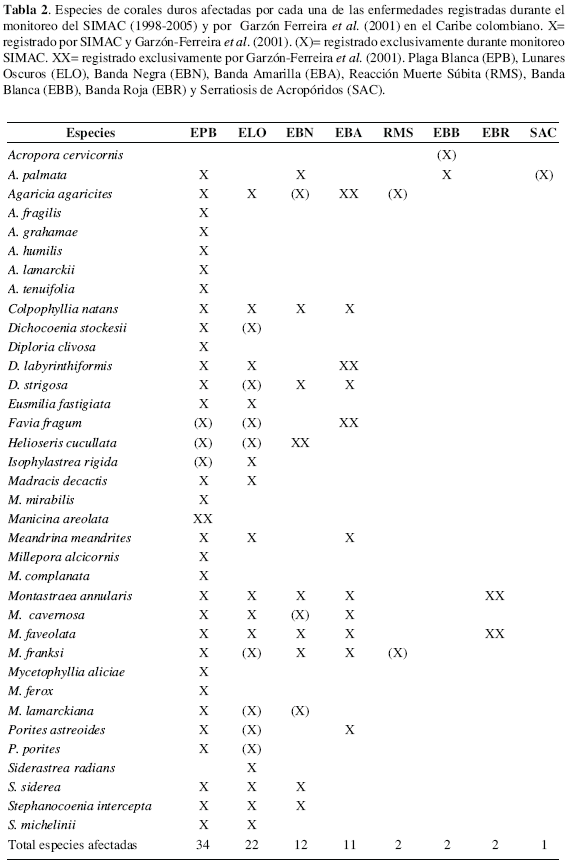
El incremento en la aparición de nuevas enfermedades coralinas y su más amplia dispersión geográfica en Colombia y el mundo es innegable (Goreau et al, 1998; Gil-Agudelo y Garzón-Ferreira, 2001; Rosenberg y Ben-Haim, 2002; Weil et al., 2002; Garzón-Ferreira et al., 2004; Sutherland et al., 2004). Hoy en día, las enfermedades son consideradas uno de los factores que ocasionan mayor pérdida de tejido coralino, convirtiéndose en un aspecto crítico en la ecología de los arrecifes de coral (Weil et al., 2002; Garzón-Ferreira et al, 2001; Sutherland et al., 2004). En consecuencia, los monitoreos sistemáticos de salud coralina en Colombia deben continuar y expandirse a otras áreas arrecifales para brindar información más sólida con el fin de explicar la variabilidad geográfica y temporal observada, involucrando, además, seguimientos particulares sobre la dinámica de la EPB y la ELO por su potencial papel en el proceso actual de deterioro arrecifal.
Perspectivas de investigación
Hasta el momento la investigación de enfermedades de corales en Colombia, al igual que el estudio de corales en general, ha sido dominada por las determinaciones en campo y estudios de línea base. En pocos casos se han realizado investigaciones que contemplen más allá de la descripción y cuantificación de estas enfermedades. Colombia posee un gran potencial para desarrollar investigaciones tendientes a resolver muchas de las incógnitas que rodean las enfermedades de corales, tales como son la identiicación de los organismos patógenos, vectores de transmisión, reservorios y modos de dispersión de estos patógenos. De otra parte, se desconocen los procesos de infección y resistencia de los corales a las enfermedades. En este marco y de acuerdo a la experiencia de los investigadores del país, los siguientes serían algunos de los posibles proyectos de investigación que deberían llevarse a cabo:
- Identificación y caracterización de los patógenos de las enfermedades coralinas de Lunares Oscuros (ELO), Banda Blanca (EBB) tipos I y II y Plaga Blanca (EPB) tipos I y III usando técnicas microbiológicas clásicas y moleculares.
- Diferenciación de los tipos I, II y III de la EPB mediante estudios moleculares y estudios detallados en campo sobre su incidencia y distribución.
- Estudios de los reservorios de EPB tipo II mediante el uso de técnicas moleculares y microbiológicas clásicas.
- Estudios de la presencia de V. shilonii y V. coralliilyticus en aguas del Caribe y Pacífico colombiano y su potencial papel en el blanqueamiento coralino en la región.
- Ensayos en laboratorio tendientes a determinar el papel de variables ambientales tales como temperatura, sedimentación y concentración de nutrientes en el proceso de enfermedades tales como EBB, ELO, Serratiosis de Acróporas (SAC), entre otras.
- Estudio de los posibles vectores de enfermedades cuyos patógenos ya han sido establecidos (EPB, EBN, SAC) mediante el uso de técnicas moleculares.
Debido a su situación geográfica y a que cuenta con arrecifes coralinos tanto en el océano Pacífico como en el mar Caribe, Colombia puede considerarse como un lugar privilegiado para realizar estudios de enfermedades en ambos lugares e incluso realizar investigaciones en enfermedades transoceánicas. Tal es el caso de:
- Profundización en el conocimiento de las enfermedades que afectan corales del Pacífico colombiano.
- Presencia de Aspergilosis en octocorales del género Pacifigorgia en el Pacífico colombiano.
- Corroborar la presencia de la EPB en corales de Malpelo y otras localidades del Pacífico, de las cuales se tienen registros iniciales (Garzón-Ferreira y Pinzón, 1999).
A su vez, deben fortalecerse los estudios de enfermedades coralinas en campo principalmente aquellos que implican el monitoreo de las zonas arrecifales colombianas para conocer el impacto a largo plazo en dichas áreas. La expansión geográfica de los programas de monitoreo permitirá también la detección de enfermedades en diferentes áreas coralinas, y debe ser acompañada de la implementación de protocolos rápidos de evaluación, así como de la toma de información complementarias para tratar de determinar los agentes causantes de las mismas. Dentro de este componente de monitoreo se recomienda:
- Evaluación de la prevalencia de enfermedades en áreas remotas del Caribe colombiano.
- Determinación de las tasas de pérdida de tejido asociadas con enfermedades como la EPB y el ELO.
- Determinar la relevancia de las enfermedades en la pérdida de cobertura coralina frente a otros agentes de mortalidad natural o antropogénica.
El desarrollo de estos temas necesita una estrecha interacción entre estudios de campo y de laboratorio. Para esto, Colombia ya posee una buena infraestructura cercana a zonas arrecifales que permite la ejecución de este tipo de investigaciones. En la costa Caribe, por ejemplo, se encuentran los laboratorios de INVEMAR y de la Universidad de Bogotá Jorge Tadeo Lozano en el área de Santa Marta. De otra parte, se cuenta con la Universidad de Cartagena, Universidad de Bogotá Jorge Tadeo Lozano y CARDIQUE en el área de Cartagena; Universidad de Córdoba y una estación remota de INVEMAR en el área del golfo de Morrosquillo y la Universidad de Antioquia en el golfo de Urabá. En el archipiélago de San Andrés y Providencia se cuenta con los laboratorios en CORALINA y la Universidad Nacional de Colombia, sede San Andrés. De otra parte, en el Pacífico colombiano están los laboratorios de la Universidad del Valle e INVEMAR. La Red de Parques Nacionales Naturales directa o indirectamente relacionados con zonas arrecifales en ambos océanos cuenta con la logística necesaria para apoyar este tipo de investigaciones en sus áreas de influencia. Los laboratorios y centros de investigación pueden servir no sólo para realizar investigaciones sobre enfermedades coralinas, sino que también pueden actuar como red de alerta temprana de posibles epidemias de enfermedades coralinas, dado que cuentan con personal capacitado. Con la creación del SIMAC se han dado los primeros pasos para integrar varias de estas entidades en torno a la investigación de arrecifes coralinos. Sin duda estos esfuerzos deben continuar en el futuro, con el ánimo de detectar cambios en la distribución e incidencia de las diferentes enfermedades coralinas hasta ahora observadas, pero también con el in de identiicar la aparición de nuevas enfermedades no detectadas con anterioridad.
De otra parte, es necesario canalizar esfuerzos conducentes a estudiar las causas de dichas enfermedades y los factores que exacerban su presencia en ciertas áreas. Para esto es imprescindible comenzar a cambiar en parte el enfoque de la investigación, pasando de los estudios descriptivos a estudios experimentales e involucrar en estas investigaciones a profesionales de otras ramas de la biología y de otras disciplinas de las ciencias. Es necesario vincular, por ejemplo, microbiólogos que se enfoquen en la identiicación de los patógenos causantes de estas enfermedades, biólogos moleculares capaces de encontrar reservorios y vectores de estos patógenos, biólogos celulares que investiguen los mecanismos de acción, químicos que identifiquen las toxinas producidas por los patógenos y su forma de acción, entre muchos otros. A pesar de que muchos de los laboratorios anteriormente mencionados no tienen las facilidades o el personal adecuado para hacer este tipo de investigaciones, ellos podrían servir como puntos de enlace y colecta de muestras para su posterior análisis en otras entidades que cuentan con este tipo de infraestructura. Este el caso de universidades como Andes, Javeriana, Jorge Tadeo Lozano, Nacional de Colombia, Antioquia, Valle, e institutos como INVEMAR y Humboldt, entre otros, los cuales cuentan con equipos y profesionales especializados en diferentes áreas del conocimiento que podrían realizar valiosos aportes a este campo de estudio.
La investigación de corales y enfermedades coralinas en Colombia tiene ya un gran reconocimiento internacional. Muestra de esto es la posición del país en la red CARICOMP, que agrupa a más de 25 laboratorios de 16 países del gran Caribe y en la que es uno de los miembros líderes. Esta colaboración ha arrojado importantes resultados no sólo desde el punto de vista del número de publicaciones realizadas por la red, sino también en la interacción permanente de los científicos colombianos con los de otros países. Como producto también de la posición de liderazgo de la investigación en arrecifes coralinos y sus enfermedades, Colombia (a través del INVEMAR) fue escogido como coordinador del nodo regional STA-GCRMN (Southern Tropical Atlantic, Global Coral Reef Monitoring Network) de la red global de monitoreo de arrecifes coralinos, en el cual están incluidos cinco países de la región (Costa Rica, Panamá, Colombia, Venezuela y Brasil).
El estudio de enfermedades de corales y en general de las de organismos marinos es un nuevo campo de mucho interés a nivel internacional, especialmente por el incremento de las mismas como consecuencia de procesos de contaminación y posiblemente por el cambio climático global. Muestra de esto es la reciente creación del grupo para estudios de enfermedades de corales del Banco Mundial y la creación de Reef Task por parte del gobierno de Estados Unidos. En consecuencia, la investigación en enfermedades de corales en Colombia debe cambiar su enfoque, pasando de estudios meramente descriptivos a una investigación multidisciplinaria e integral que involucre aspectos de ecología, microbiología y química. Además es importante buscar aliados tanto nacionales como internacionales que permitan a la comunidad científica nacional ser competitiva y líder de este tipo de investigaciones en la región.
AGRADECIMIENTOS
Los autores desean agradecer a todos los investigadores, estudiantes y pasantes que han colaborado con nuestros estudios de enfermedades coralinas en Colombia; igualmente a los dos evaluadores anónimos del presente artículo, los cuales ayudaron a mejorar significativamente la calidad del mismo. Los fondos para las investigaciones de enfermedades coralinas realizadas en Colombia han sido aportados en parte por el Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología COLCIENCIAS (Proyecto Colciencias-BID 2105-09-327-97), del Instituto de Investigaciones Marinas y Costeras INVEMAR, UNEP-CAR/ RCU, National Science Foundation (NSF OCE-0326269) y National Oceanic and Atmospheric Administration (NOAA-CRES 0648-0384). DLG-A quiere también agradecer la beca FULBRIGHT-COLCIENCIAS-DNP (081/2000). Agradecemos especialmente el apoyo logístico del CEINER, Corporación CORALINA, Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales UAESPNN, Universidad Nacional y Universidad del Valle.
BIBLIOGRAFÍA
1 Aeby, G. S. 1998. A digenean metacercaria from the reef coral, Porites compressa, experimentally identified as Podocotyloides stenometra. J. Parasitol., 84 (6): 1259-1261. [ Links ]
2 Alker A. P., G. W. Smith y K. Kim. 2001. Characterization of Aspergillus sydowii (Thom et Church) a fungal pathogen of Caribbean sea fan corals. Hydrobiologia, 460: 105-111. [ Links ]
3 Alker, A. P., K. Kim, D. H. Dube y C. D. Harvell. 2004. Localized induction of a generalized response against multiple biotic agents in Caribbean sea fans. Coral reefs, 23: 397-405. [ Links ]
4 Al-Moghrabi, S. M. 2001. Unusual black band disease (BBD) outbreak in the northern tip of the Gulf of Aqaba (Jordan). Coral Reefs, 19: 330-331. [ Links ]
5 Antonius, A. 1977. Coral mortality in reefs: a problem for science and management. Proc. 3rd Internat. Coral Reef Symp., 2: 617-623. [ Links ]
6 Antonius, A. 1981a. The "band" disease in coral reefs. Proc. 4th Internat. Coral Reef Symp., 2: 7-14. [ Links ]
7 Antonius, A. 1981b. Coral reef pathology: A review. Proc. 4th Internat. Coral Reef Symp., 2: 3-6. [ Links ]
8 Antonius, A. 1985a. Black band disease infection experiments on hexacorals and octocorals. Proc. 5th Internat. Coral Reef Symp., 6: 155-160. [ Links ]
9 Antonius, A. 1985b. Coral diseases in the Indo-Pacific: A first record. P.S.Z.N.I. Mar. Ecol., 6 (3): 197-218. [ Links ]
10 Antonius, A. 1988. Distribution and dynamics of coral diseases in the eastern Red sea. Proc. 6th Internat. Coral Reef Symp., 2: 293-298. [ Links ]
11 Antonius, A. 1999. Halofolliculina corallasia, a new coral-killing cilicate on Indo-Pacific reefs. Coral Reefs, 18 (3): 300. [ Links ]
12 Antonius, A. y D. Lipscomb. 2001. First protozoan coral-killer identified in the Indo-Pacific. Atoll. Res. Bull., 481-493: 1-21. [ Links ]
13 Aronson, R. B. y W. F. Precht. 2001. White band disease and the changing face of Caribbean coral reefs. Hydrobiologia, 460: 25-38. [ Links ]
14 Bak, R. P. M. 1983. Neoplasia, regeneration and growth in the reef-building coral Acropora palmata. Mar. Biol., 77 (3): 221-227. [ Links ]
15 Banin, E, T. Israely, A. Kushmaro, Y. Loya, E. Orr y E. Rosenberg. 2000. Penetration of the coral-bleaching Vibrio shiloi into Oculina patagonica. Appl. Environ. Microbiol., 66: 3031-3036. [ Links ]
16 Banin, E., T. Israely, M. Fine, Y. Loya y E. Rosenberg. 2001a. Role of endosymbiotic zooxanthellae and coral mucus in the adhesion of the coral-bleaching pathogen Vibrio shiloi to its host. FEMS Micriobiol. Lett., 199: 33-37. [ Links ]
17 Banin, E. T., K. H. Sanjay, F. Naider y E. Rosenberg. 2001b. A proline-rich peptide from the coral pathogen Vibrio shiloi that inhibits photosynthesis of zooxanthellae. Appl. Environ. Microbiol., 67 (4): 1536-1541. [ Links ]
18 Barrios, L. M. 2000. Evaluación de las principales condiciones de deterioro de los corales pétreos en el Caribe colombiano. Tesis Maestría Biol. Mar., Univ. Nal. Col., Santa Marta. 155 p. [ Links ]
19 Ben-Haim, Y. y E. Rosenberg. 2002. A novel Vibrio sp. pathogen of the coral Pocillopora damicornis. Mar. Biol., 141: 47-55. [ Links ]
20 Ben-Haim, Y., F. L. Thompson, C. C. Thompson, M. C. Cnockaert, B. Hoste, J. Swings y E. Rosenberg E. 2003a. Vibrio coralliilyticus sp. nov., a temperature-dependent pathogen of the coral Pocillopora damicornis. Int. J. Syst. Evol. Microbiol., 53: 309-315. [ Links ]
21 Ben-Haim, Y., M. Zicherman-Keren y E. Rosenberg. 2003b. Temperature-regulated bleaching and lysis of the coral Pocillopora damicornis by the novel pathogen Vibrio coralliilyticus. Appl. Environ. Microbiol, 69: 4236-4242. [ Links ]
22 Birkeland, R. (Ed.). 1997. Life and death of coral reefs. Chapman and Hall, Nueva York. 536 p. [ Links ]
23 Bruckner, A. W. 2002. Priorities for effective managment of coral diseases. Technical Memorandum. [ Links ]
24 Bruckner, A. y R. J. Bruckner. 1997. Outbreak of coral disease in Puerto Rico. Coral Reefs, 16 (5): 260. [ Links ]
25 Carlton, R. G. y L. L. Richardson. 1995. Oxygen and sulfide dynamics in a horizontally migrating cyanobacterial mat: Black band disease of corals. FEMS Microb. Ecol., 18: 155-162. [ Links ]
26 Cervino, J., T. J. Goreau, I. Nagelkerken, G. W. Smith y R. Hayes. 2001. Yellow band and dark spot syndromes in Caribbean corals: distribution, rate of spread, cytology, and effects on abundance and division rate of zooxanthellae. Hydrobiologia, 460: 53-63. [ Links ]
27 Cervino, J., R. L. Hayes, S. W. Poison, S. C. Poison, T. J. Goreau, R. J. Martínez y G. W. Smith. 2004. Relationship of Vibrio species infection and elevated temperatures to yellow blotch/band disease in Caribbean corals. Appl. Environ. Microbiol., 70: 6855-6864. [ Links ]
28 Coles, S. L. 1994. Extensive coral disease outbreak at Fahl Island, Gulf of Oman, Indian Ocean. Coral Reefs, 13: 242. [ Links ]
29 Denner, E. B. M., G. W. Smith, H. J. Busse, P. Schumann, T. Narzt, S. W. Poison, W. Lubitz y L. L. Richardson. 2003. Aurantimonas coralicida gen. nov. sp. nov., the causative agent of white plague type II on Caribbean scleractinian corals. Int. J. Syst. Evol. Microbiol., 53 (4): 1115-1122. [ Links ]
30 Díaz, J. M., J. Garzón-Ferreira y S. Zea. 1995. Los arrecifes coralinos de la isla de San Andrés, Colombia: estado actual y perspectivas para su conservación. Acad. Colomb. Cienc. Exac. Fís. Nat., 7: 150 p. [ Links ]
31 Díaz, J. M., L. M. Barrios, M. H. Cendales, J. Garzón-Ferreira, J. Geister, M. López-Victoria, G. H. Ospina, E. Parra-Velandia, J. Pinzón, B. Vargas-Ángel, F. A. Zapata y S. Zea. 2000. Areas coralinas de Colombia. Serie Publ. Esp. 5, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta. 176 p. [ Links ]
32 Dube, D., K. Kim, A. A. Alker y C. D. Harvell. 2002. Size structure and geographic variation in chemical resistance of sea fan corals (Gorgonia ventalina) against a fungal pathogen. Mar. Ecol. Prog. Ser., 231: 139-150. [ Links ]
33 Dubinsky, Z. (Ed.). 1990. Coral reefs. Elsevier Science Publishers B.V., Amsterdam. 550 p. [ Links ]
34 Ducklow, H. y R. Mitchell. 1979. Observations on naturally and artificially diseased tropical corals: a scanning electron microscope study. Microb. Ecol., 5: 325-223. [ Links ]
35 Dustan, P. 1977. Vitality of reef coral population off Key Largo, Florida: recruitment and mortality. Environ. Geol., 2: 51-58. [ Links ]
36 Dustan, P. y J. C. Halas. 1987. Changes in the reef-coral community of Carysfort Reef, Key Largo, Florida: 1974-1982. Coral Reefs, 6 (2): 91-106. [ Links ]
37 Erhardt, H y B. Werding. 1975a. Los corales (Anthozoa e Hidrozoa) de Granate, pequeña bahía al este de Santa Marta, Colombia. Cadalsia, 11 (53): 107-138. [ Links ]
38 Erhardt, H. y B. Werding. 1975b. Los corales (Anthozoa e Hidrozoa) de la bahía de Santa Marta, Colombia. Bol. Mus. Mar, 7: 3-50. [ Links ]
39 Frías-López, J., A. L. Zerkle, G. T. Bonheyo y B. W. Fouke. 2002. Partitioning of bacterial communities between seawater and healthy, black band diseased, and dead coral surfaces. Appl. Environ. Microbiol., 68: 2214-2228. [ Links ]
40 Frías-López, J. A., G. T. Bonheyo, Q. Jin y B. W. Fouke. 2003. Cyanobacteria associated with coral black band disease in Caribbean and Indo-Pacific reefs. Appl. Environ. Microbiol., 69: 2409-2413. [ Links ]
41 Galloway, S. B., T. M. Work, V. S. Bochsler, R. A. Harley, E. Kramarsky-Winter, S. M. McLaughlin, C. U. Meteyer, J. F. Morado, J. H. Nicholson, P. G. Parnell, E. C. Peters, T. L. Reynolds, D. S. Rotstein, L. Sileo y C. M. Woodley. 2007. Coral disease and health workshop: Coral histopathology II. NOAA Technical Memorandum NOS NCCOS 56 and NOAA Technical Memorandum CRCP 4. National Oceanic and Atmospheric Administration, Silver Spring, EE. UU. 84 p. [ Links ]
42 Garcés, A. 2000. Desarrollo de la enfermedad de los lunares oscuros los corales pétreos Montastraea annularis y Siderastrea sidérea en la bahía de Gayraca, Caribe colombiano. Tesis Biol. Mar., Univ. Bogotá Jorge Tadeo Lozano, Santa Marta. 101 p. [ Links ]
43 Garrett, P. y P. Ducklow. 1975. Coral disease in Bermuda. Nature, 253: 349-350. [ Links ]
44 Gateño, D., A. León, Y. Barki, J. Cortés y B. Rinkevich. 2003. Skeletal tumor formations in the massive coral Pavona clavus. Mar. Ecol. Prog. Ser., 258: 97-108. [ Links ]
45 Garzón-Ferreira, J. 1999. SIMAC: a new program to monitor coral reef condition in Colombia. Reef Encounter, 25: 10-15. [ Links ]
46 Garzón-Ferreira, J. y S. Zea. 1992. A mass mortality of Gorgonia ventalina (Cnidaria:Gorgoniidae) in the Santa Marta area, Caribbean coast of Colombia. Bull. Mar. Sci., 50 (3): 522-526. [ Links ]
47 Garzón-Ferreira, J. y D. L. Gil. 1998. Another unknown Caribbean coral phenomenon? Reef Encounter, 24: 10. [ Links ]
48 Garzón-Ferreira, J. y J. H. Pinzón. 1999. Rapid assessment of coral community structure and health at Malpelo island (Colombian Pacific). Bol. Invest. Mar. Cost., 28: 137-154. [ Links ]
49 Garzón-Ferreira, J., D. L. Gil-Agudelo, L. M. Barrios y S. Zea. 2001. Stony coral diseases observed in southwestern Caribbean reefs. Hydrobiologia, 460: 65-69. [ Links ]
50 Garzón-Ferreira, J., A. Rodríguez-Ramírez, S. Bejarano-Chavarro, R. Navas-Camacho y M. C. Reyes-Nivia. 2002b. Estado de los arrecifes coralinos. 30-40. En: INVEMAR (Ed.). Informe del estado de los ambientes marinos y costeros en Colombia: Año 2001. Serie Publ. Periód. 8, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta. 178 p. [ Links ]
51 Garzón-Ferreira, J. A., A. Rodríguez-Ramírez, S. Bejarano-Chavarro, R. Navas-Camacho, M. C. Reyes-Nivia, P. Herrón, F. Zapata, J. Rojas y O. Caucali. 2004. Estado de los arrecifes coralinos en Colombia. 79-135. En: INVEMAR (Ed.). Informe del estado de los ambientes marinos y costeros en Colombia: Año 2003. Serie Publ. Periód. 8., Instituto de Investigaciones Marinas y Costeras- INVEMAR, Santa Marta. 329 p. [ Links ]
52 Geister, J. 1977. The influence of wave exposure on the ecological zonation of Caribbean coral reefs. Proc. 3rd Internat. Coral Reef Symp., 1: 23-29. [ Links ]
53 Gil-Agudelo, D. L. y J. Garzón-Ferreira. 2001. Spatial and seasonal variation of the dark spots disease in coral communities of the Santa Marta area (Colombian Caribbean). Bull. Mar. Sci., 69 (2): 619-629. [ Links ]
54 Gil-Agudelo, D. L., G. W. Smith, J. Garzon-Ferreira, E. Weil y D. Petersen. 2004. Dark spots disease and yellow band disease, two poorly known coral diseases with high incidence in Caribbean reefs. 337-349. En: Rosenberg, E. y Y. Loya (Eds.). Coral health and disease. Springer-Verlag, Berlin. 488p. [ Links ]
55 Gil-Agudelo, D. L., G. W. Smith y E. Weil. 2006a. Fulfilling Koch's postulates for white band disease type II. Rev. Biol. Trap., 54 (Suppl. 3): 59-67. [ Links ]
56 Gil-Agudelo, D. L., C. Myers, G. W. Smith y K. Kim. 2006b. Changes in the microbial communities associated with Gorgonia ventalina during aspergillosis infection. Dis. Aquat. Org., 69: 89-94. [ Links ]
57 Gil-Agudelo, D. L., D. Fonseca, J. Garzon-Ferreira, E. Weil y G. W. Smith. 2007. Bacterial communities associated with the mucopolysaccharide layers of three coral species affected and unaffected with dark spots disease. Can. J. Microbiol., 53: 465-471. [ Links ]
58 Ginsburg, R. N. (Ed.). 1994. Proceedings of the colloquium on global aspects of coral reefs. Health, Hazards, and History. University of Miami, Rosenstiel School of Marine and Atmospheric Science, Miami. 420 p. [ Links ]
59 Gladfelter, W. B. 1982. White-band disease in Acropora palmata: implications for the structure and growth of shallow reefs. Bull. Mar. Sci., 32 (2): 639-643. [ Links ]
60 Goreau, T. F. 1959. The ecology of Jamaican coral reefs. I. Species composition and Zonation. Ecology, 40: 67-90. [ Links ]
61 Goreau, T. J., J. Cervino, M. Goreau, R. Hayes, M. Hayes, L. Richardson, G. W. Smith, K. DeMeyer, I. Nagelkerken, J. Garzön-Ferreira, D. Gil, G. Garrison, E. H. Williams, L. Bunkley-Williams, C. Quirolo, K. Patterson, J. W. Porter y K. Porter. 1998. Rapid spread of diseases in Caribbean coral reefs. Rev. Biol. Trap., 46 (Supl. 5): 157-171. [ Links ]
62 Grigg, R. W. y S. J. Dollar. 1990. Natural and anthropogenic disturbance on coral reefs. 439-452. En: Dubinsky, Z. (Ed.). Coral reefs. Elsevier Science Publisher B.V., Amsterdam. 550 p. [ Links ]
63 Grygier, M. J. y S. D. Cairns. 1996. Suspected neoplasm in deep-sea corals (Scleractinia: Oculinidae: Madrepora spp.) reinterpreted as galls caused by Petrarca madreporae n. sp. (Crustacea: Ascothoracida: Petrarcidae). Dis. Aquat. Org., 24 (1): 61-69. [ Links ]
64 Holden, C. 1996. Coral disease hot spot in the Florida Keys. Science, 274: 2017. [ Links ]
65 Hughes, T. P. 1994. Catastrophes, phase shift and large-scale degradation of a Caribbean coral reef. Science, 265: 1547-1551. [ Links ]
66 Israely, T., E. Banin y E. Rosenberg. 2001. Growth, differentiation and death of Vibrio shiloi in coral tissue as a function of seawater temperature. Aquat. Microb. Ecol., 24: 1-8. [ Links ]
67 Kim, K. y C. D. Harvell. 2002. Aspergillosis of sea fan corals: disease dynamics in the Florida Keys. 813-824. En: Porter, J. W. y K. G. Porter (Ed.). The Everglades, Florida Bay, and coral reefs of the Florida Keys: an ecosystem sourcebook. CRC Press, Nueva York. 1024 p. [ Links ]
68 Kim, K, C. D. Harvell, P. D. Kim, G. W. Smith y S. M. Merkel. 2000a. Fungal disease resistance of Caribbean sea fan corals (Gorgonia spp.). Mar. Biol., 136: 259-267. [ Links ]
69 Kim, K, P. D. Kim, A. P. Alker y C. D. Harvell. 2000b. Chemical resistance of gorgonian corals against fungal infections. Mar. Biol, 137: 393-401. [ Links ]
70 Korrubel, J. L. y B. R. Riegl. 1998. A new disease from the southern Arabian Gulf. Coral reefs, 17 (1): 22. [ Links ]
71 Kushmaro, A., E. Rosenberg, M. Fine y Y. Loya. 1997. Bleaching of the coral Oculina patagonica by Vibrio AK -1. Mar. Ecol. Prog. Ser., 147: 159-165. [ Links ]
72 Kuta, K. G. y L. L. Richardson. 1996. Abundance and distribution of black band disease on coral reefs in the northern Florida Keys. Coral reefs, 14 (1): 1-5. [ Links ]
73 Littler, M. M. y D. S. Littler. 1996. Black band disease in the South Pacific. Coral Reefs, 15 (1): 20. [ Links ]
74 Malakoff, D. 1997. Extinction on the high seas. Science, 277: 487-488. [ Links ]
75 Madigan, M. T., J. M. Martinko y J. Parker. 2003. Brock biology of microorganisms. Décima edición. Pearson Education Inc. Upper Saddle River, EE. UU. 1019 p. [ Links ]
76 Nagelkerken, I., K. Buchan, G. Smith, K. Bonair, P. Bush, J. Garzón-Ferreira, L. Botero, P. Gayle, C. Heberer, C. Petrovic, L. Pors y P. Yoshioka. 1997a. Widespread disease in Caribbean sea fans: I. Spreading and general characteristics. Proc. 8th Internat. Coral Reef Symp., 1: 679-682. [ Links ]
77 Nagelkerken, I., K. Buchan, G.W. Smith, K. Bonair, P. Bush, J. Garzón-Ferreira, L. Botero, P. Gayle, C.D. Harvell, C. Heberer, K. Kim, C. Petrovic, L. Pors y P. Yoshioka. 1997b. Widespread disease in Caribbean sea fans: II. Patterns of infection and tissue loss. Mar. Ecol Prog Ser., 160: 255-263. [ Links ]
78 Nugues, M. M., G. W. Smith, R. J. van Hooidonk, M. I. Seabra, y R. P. M. Bak. 2004. Algal contact as a trigger for coral disease. Ecology letters, 7: 919-923. [ Links ]
79 Patterson, K. L., J. W. Porter, K. B. Ritchie, S. W. Polson, E. Mueller, E. C. Peters, D. L. Santavy y G. W. Smith.2002. The etiology of white pox, a lethal disease of the Caribbean elkhorn coral Acropora palmata. Proc. Natl. Acad. Sci. USA, 99: 8725-8730. [ Links ]
80 Peters, E. C. 1993. Disease of other invertebrate Phyla: Porifera, Cnidaria, Ctenophora, Annelida, Echinodermata. 393-449. En: Couch, J. A. y J. W. Fournie (Ed.). Advances in fisheries science. Pathobiology of marine and estuarine organisms, CRC Press, Boca Ratón, EE. UU. 576 p. [ Links ]
81 Peters, E. C. 1997. Diseases of coral-reefs organisms. 114-139. En: Birkeland, C. (Ed.). Life and death of coral reefs. Chapman and Hall, Nueva York. 536 p. [ Links ]
82 Peters, E. C., P. Yevich y J. Oprandy. 1983. Possible causal agent of 'white band disease' in Caribbean acroporid corals. J. Invertebr. Pathol., 41: 394-396. [ Links ]
83 Peters, E. C. 1984. A survey of cellular reactions to environmental stress and disease in Caribbean scleractinian corals. Helgoländer Meeresuntersuchungen, 37: 113-137. [ Links ]
84 Peters, E. C., J. C. Halas y H. B. McCarty. 1986. Calicoblastic neoplasms in Acropora palmata, with a review of reports on anomalies of growth and form in corals. J. Natl. Cancer Inst., 76 (5): 895-912. [ Links ]
85 Prahl, H. v. y H. Erhardt. 1985. Colombia: corales y arrecifes coralinos. FEN Colombia, Bogotá, 295 p. [ Links ]
86 Ramos-Flores, T. 1983. Lower marine fungus associated with black line disease in star corals (Montastrea annularis , E. & S.). Biol. Bull. Mar. Biol. Lab. Woods Hole, 165 (2): 424-435. [ Links ]
87 Ravindran, J. y C. Raghukumar. 2002. Pink line syndrome (PLS) in the scleractinian coral Porites lutea. Coral Reefs, 21 (3): 252. [ Links ]
88 Raymundo, L. J. H., C. D. Harvell y T. L. Reynolds. 2003. Porites ulceraive white spot disease: description, prevalence, and host range of a new coral disease affecting Indo-Pacific reefs. Dis. Aquat. Org., 56: 95-104. [ Links ]
89 Reeves, L. 1994. Newly discovered: Yellow band disease strikes Keys Reefs. Underwater USA, 11 (8): 16. [ Links ]
90 Richardson, L. L. 1992. Red band disease: A new cyanobacterial infestation of corals. Proc. Am. Acad. Underwater Sci. 12th Annual Scientific Diving Symposium, 153-160. [ Links ]
91 Richardson, L. L. 1998. Coral diseases: what is really known? TREE. 13: 438-443. [ Links ]
92 Richardson, L. L., K. G. Kuta, S. Schnell y R. G. Carlton. 1997. Ecology of the black band disease microbial consortium. Proc. 8th Internat. Coral Reef Symp., 1: 597-600. [ Links ]
93 Richardson, L. L., W. M. Goldberg, K. G. Kuta, R. B. Aronson, G. W. Smith, K. B. Ritchie, J. C. Halas, J. S. Feingold y S, L. Miller. 1998a. Florida's mystery coral-killer identified. Nature, 392 (6676): 557-558. [ Links ]
94 Richardson, L. L., W. M. Goldberg, R. G. Carton y J. C. Halas 1998b. Coral disease outbreak in the Florida Keys: plague type II. Rev. Biol. Trop., 46: 187-198. [ Links ]
95 Richardson, L. L., G. W. Smith, K. B. Ritchie y R. G. Carlton 2001. Integrating microbiological, microsensor, molecular, and physiologic techniques in the study of coral disease pathogenesis. Hydrobiologia, 460: 71-89. [ Links ]
96 Riegl, B. 2002. Effects of the 1996 and 1998 positive sea-surface temperature anomalies on corals, coral diseases and fish in the Arabian Gulf (Dubai, UAE). Mar. Biol., 140: 29-40. [ Links ]
97 Ritchie, K. B. y G. W. Smith. 1995. Preferential carbon utilization by surface bacterial communities from water mass, normal, and white-band diseased Acropora cervicornis. Mol. Mar. Biol. Biotechnol., 4: 345-352. [ Links ]
98 Ritchie, K. B. y G. W. Smith. 1998. Type II white-band disease. Rev. Biol. Trap., 46: 199-203. [ Links ]
99 Rosenberg, E. 2004. The bacterial disease hypothesis of coral bleaching. 445-461. En: Rosenberg, E. y Y. Loya. Coral health and disease. Springer-Verlag, Berlin. 488 p. [ Links ]
100 Rosenberg, E. y Y. Ben-Haim. 2002. Microbial diseases of corals and global warming. Environ. Microbiol, 4 (6): 318-326. [ Links ]
101 Rosenberg, E. y Y. Loya. (Eds.). 2004. Coral health and disease. Springer-Verlag. Berlin. 488 p. [ Links ]
102 Rützler, K. y D. Santavy. 1983. The black band disease of Atlantic reef corals. I. Description of the cyanophyte pathogen. P.S.Z.N.I. Mar. Ecol, 4 (4): 301-319. [ Links ]
103 Rützler, K, D. Santavy y A. Antonius. 1983. The black band disease of Atlantic reef corals III. Distribution, Ecology and Development. P.S.Z.N.I. Mar. Ecol., 4 (4): 329-358. [ Links ]
104 Santavy, D. L. y E. C. Peters. 1997. Microbial pests: coral disease in the western Atlantic. Proc. 8th Internat. Coral Reef Symp., 1: 607-612. [ Links ]
105 Santavy, D. L., E. C. Peters, C. Quirolo, J. W. Porter y C. N. Bianchi. 1999. Yellow-blotch disease outbreak of reefs of the San Bias Islands, Panama. Coral reefs, 18 (1): 97. [ Links ]
106 Shinn, E. A., G. W. Smith, J. M. Próspero, P. Betzer, M. L. Hayes, V. Garrison y R. T. Barber. 2000. African dust and the demise of Caribbean coral reefs. Geophys. Res. Lett., 27: 3029-3032. [ Links ]
107 Smith, G. W., I. A. Nagelkerken y K. B. Ritchie. 1996. Caribbean sea-fan mortalities. Nature, 383: 487. [ Links ]
108 Smith, G. W., C. D. Harvell y K. Kim. 1998. Response of sea fans to infection with Aspergillus sp. (Fungi). Rev. Biol. Trop., 46: 205-208. [ Links ]
109 Solano, O. D., G. Navas-Suárez y S. K. Moreno-Forero. 1993. Blanqueamiento coralino de 1990 en el Parque Nacional Natural Corales del Rosario (Caribe colombiano). An. Inst. Invest. Mar. Punta Betín, 22: 97-111. [ Links ]
110 Squires, D. F. 1965. Neoplasia in a coral? Science, 148: 503-505. [ Links ]
111 Sutherland, K. P. y K. B. Ritchie 2004. White pox disease of the Caribbean elkhorn coral Acropora palmata. 289-300. En: Rosenberg, E. y Y. Loya. (Eds.). Coral health and disease. Springer-Verlag, Berlin. 488 p. [ Links ]
112 Sutherland, K. P., J. W. Porter y C. Torres. 2004. Disease and immunity in Caribbean and Indo-Pacific zooxanthellate corals. Mar. Ecol. Prog. Ser., 266: 273-302. [ Links ]
113 Thompson, F. L., B. Hoste, C. C. Thompson, G. Huys y J. Swings. 2001. The coral bleaching Vibrio shiloi Kushmaro et al. 2001 is a later synonym of Vibrio mediterranei Pujalte and Garay 1986. Syst. Appl. Microbiol., 24: 516-519. [ Links ]
114 Toller, W. W., R. Rowan y N. Knowlton. 2001. Repopulation of zooxanthellae in the Caribbean corals Montastraea annularis and M. faveolata following experimental and disease-associated bleaching. Biol. Bull., 201 (3): 360-373. [ Links ]
115 Toren, A., L. Landau, A. Kushmaro, Y. Loya y E. Rosenberg. 1998. Effects of temperature on adhesion of Vibrio strain AK-1 to Oculina patagonica and on coral bleaching. Appl. Environ. Microbiol., 64: 1379-1384. [ Links ]
116 Veron, J. E. N. 1995. Corals in space and time: The biogeography and evolution of the Scleractinia. Cornell University Press, Ithaca, EE. UU. 321 p. [ Links ]
117 Weil, E., I. Urreiztieta y J. Garzón-Ferreira. 2002. Geographic variability in the incidence of coral and octocoral diseases in the wider Caribbean. Proc. 9th Internat. Coral Reef Symp. Bali, 2: 1231-1237. [ Links ]
118 Weil, E. 2006. Coral reef diseases in the wider Caribbean. 35-68. En: Rosenberg, E. y Y. Loya (Eds.). Coral health and disease. Springer-Verlag, Berlín. 488 p. [ Links ]
119 Weil, E., G. Smith y D. L. Gil-Agudelo. 2004. Status and progress in coral reef disease research. Dis. Aqua. Org., 69 (1): 1-7. [ Links ]
120 Weir-Brush, J., V. H. Garrison, G. W. Smith y E. A. Shinn. 2004. The relationship between gorgonian coral (Cnidaria: Gorgonaceae) diseases and African dust storms. Aerobiologia, 20 (2): 119-126. [ Links ]
121 Werding, B. y H. Sánchez. 1988. Deterioro observado en las formaciones coralinas de la bahía de Santa Marta, Colombia. An. Inst. Inv. Mar. Punta Betín, 18: 9-16. [ Links ]
122 Werding, B. y H. Sánchez. 1989. The coral formations and their distributional pattern along a wave exposure gradient in the area of Santa Marta, Colombia. Medio Ambiente, 10 (2): 61-68. [ Links ]
123 Willis, B. L., C. A. Page y E. A. Dinsdale. 2004. Coral disease in the Great Barrier Reef. 69-104. En: Rosenberg, E. y Y. Loya (Eds.). Coral health and disease. Springer-Verlag, Berlín. 488 p. [ Links ]
124 Work, T. M., G. S. Aeby, F. G. Stanton y D. Fenner. 2008. Overgrowth of fungi (endolithic hypermycosis) associated with multifocal to diffuse distinct amorphous dark discoloration of corals in the Indo- Pacific. Coral Reefs, 27: 663. [ Links ]
125 Yamashiro, H., M. Yamamoto y R. van Woesik. 2000. Tumor formation on the coral Montipora informis. Dis. Aquat. Org., 41: 211-217. [ Links ]
126 Zapata, F. A., B. Vargas-Ángel y J. Garzón-Ferreira. 2001. Salud y conservación de las comunidades coralinas. 41-50. En: Barrios, L. M. y M. López-Victoria. (Eds.). Gorgona marina. Contribución al conocimiento de una isla única. Serie Publ. Esp. 7. Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta. 160 p. [ Links ]
FECHA DE RECEPCIÓN: 09/02/07 FECHA DE ACEPTACIÓN: 13/10/09
*Contribución No. 1037 del Instituto de Investigaciones Marinas y Costeras-INVEMAR.
