










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.40 no.1 Santa Marta Jan./June 2011
FORMACIONES CORALINAS DE PROFUNDIDAD: CRITERIOS BIOLÓGICOS PARA LA CONFORMACIÓN DE ÁREAS MARINAS PROTEGIDAS DEL MARGEN CONTINENTAL (100-300 m) EN EL CARIBE COLOMBIANO*
DEEP-SEA CORALS FORMATIONS: BIOLOGIC CRITERIA FOR THE ESTABLISHMENT OF MARINE PROTECTED AREAS ON A CONTINENTAL MARGIN (100-300 M) IN THE COLOMBIAN CARIBBEAN
Juan Diego Urriago1,2, Nadiezhda Santodomingo1,3 y Javier Reyes1
1 Instituto de Investigaciones Marinas y Costeras-INVEMAR. Cerro Punta Betín, Santa Marta, Colombia. j_reyes_forero@hotmail.com
2 Hong Kong Baptist University. The Swire Institute of Marine Science, The University of Hong Kong, Cape d'Aguilar Road, Shek O. Hong Kong (dirección actual). urriago@gmail.com
3 Natural History Museum, Cromwell Road, SW7 5BD, Londres, Reino Unido (dirección actual). nsantodomingo@gmail.com
RESUMEN
Esta investigación se realizó con el fin de establecer criterios biológicos evaluables, que sirvan de herramienta para incluir a las comunidades de corales azooxantelados de profundidad como ecosistemas estratégicos a conservar en el marco de los sistemas de Áreas Marinas Protegidas (AMP) en Colombia. Se hace énfasis en la recientemente creada AMP de los Archipiélagos del Rosario y de San Bernardo (ARSB). La información utilizada en este análisis fue extraída de los proyectos MACROFAUNA I-II (1998-2001). A través de análisis ecológicos se pudo establecer que los corales profundos formadores de hábitat Anomocora fecunda, Eguchipsammia cornucopia, Madracis asperula, M. myriaster y Madrepora carolina favorecen el desarrollo de una alta biodiversidad de fauna acompañante en el AMP ARSB. Los parámetros ecológicos evaluados en estas comunidades (riqueza, abundancia y uniformidad) se enmarcaron dentro de los criterios de biodiversidad, naturalidad, representación y heterogeneidad de hábitats, calidad de único y de exclusión, resaltando la importancia de estas comunidades en el contexto del AMP ARSB.
PALABRAS CLAVE: Biodiversidad, Corales de profundidad, Criterios biológicos, Áreas Marinas Protegidas, Archipiélagos del Rosario y de San Bernardo.
ABSTRACT
We analyzed data from previous research in the Rosario and San Bernardo Archipelagos (MACROFAUNA I-II, 1998-2001) to evaluate whether the deep-sea azooxanthellate coral communities fulfill the criteria for a Marine Protected Areas System in Colombia. The azooxanthellate corals Anomocora fecunda, Eguchipsammia cornucopia, Madracis asperula, Madracis myriaster and Madrepora carolina were identified as habitatforming corals that create particular conditions favoring increased faunal diversity in the ARSB-MPA. These deep-sea coral communities had increased species richness, abundance and uniformity. The biological parameters assessed for these coral communities were considered within the biological criteria of biodiversity, naturalness, habitat representation and heterogeneity, uniqueness and exclusion, highlighting the importance of deep sea coral communities in the context of the ARSB-MPA.
KEY WORDS: Biodiversity, Azooxanthellate corals, Biological criteria, Marine Protected Areas, Rosario and San Bernardo Archipelagos.
INTRODUCCIÓN
En los últimos años se ha descubierto que extensas construcciones principalmente formadas por corales azooxantelados de profundidad poseen diversidades análogas a la de los arrecifes coralinos someros (Tabla 1; Roberts et al., 2006). Estas comunidades coralinas proporcionan un hábitat esencial para muchas especies de peces e invertebrados (estrellas de mar, crinoideos, nudibranquios, pulpos, caracoles, esponjas y anémonas, entre otros), los cuales encuentran en ellas alimento, áreas de desove, cría y protección ante depredadores (Krieger, 2001; Tunesi et al., 2001). Como las principales especies de corales que construyen el hábitat no poseen simbiosis con algas zooxantelas, sus colonias pueden desarrollarse a grandes profundidades marinas (hasta 4000 m), ya que no están restringidas a la zona fótica de la columna de agua. Incluso en las zonas templadas llegan a colonizar hábitats someros, desde 50 hasta 1000 m de profundidad, donde adicionalmente se presentan bajas temperaturas (4 a 12 °C), un régimen de corrientes fuertes y alto contenido de materia orgánica, con el fin de cumplir con los requerimientos nutricionales que estos corales necesitan para su crecimiento y desarrollo (Roberts et al., 2009).

A pesar de que el primer registro de estas comunidades data del siglo XVIII, sólo durante los últimos años se ha reconocido la importancia de las comunidades coralinas de profundidad y ya se cuenta con algunas iniciativas para su protección (Roberts y Hirshfield, 2004; King y Beazley, 2005). Se ha identificado que las principales actividades humanas que amenazan la preservación de estos ecosistemas son la pesca de arrastre, extracción de hidrocarburos y cableado submarino (Davies et al., 2007). Del estudio de estos factores de riesgo han surgido numerosas iniciativas con el fin de minimizar sus impactos. Con respecto al establecimiento de Áreas Marinas Protegidas (AMP) que contienen estos ecosistemas, en Canadá se tienen como ejemplos al Basin Head (Prince Edward Island), Gilbert Bay (Labrador) y Eastport (Newfoundland), cuyas AMP están enfocadas a la protección e investigación de los corales de profundidad como un esfuerzo en la conservación de las comunidades bentónicas en estas áreas (Butler, 2005). Algunas de las AMP que aún están en proceso de declaración, donde los corales de profundidad han sido considerados como el principal objeto de conservación son Rockall Bank y Darwin Mounds en el Reino Unido, Galicia Bank en España, Josefine Bank en Portugal, Skagerrak en Suecia, Faroe Bank en las islas Faroe, Rost Reef y Sula Ridge en Noruega (UNEP, 2006). Es claro que todos estos esfuerzos de protección se han concentrado principalmente en países localizados en zonas templadas de Europa y Norteamérica, donde además de contarse con las tecnologías para el estudio de ecosistemas profundos, los impactos causados por las pesquerías y la explotación de hidrocarburos han llamado la atención de científicos y conservacionistas (Roberts y Hirshfield, 2004).
Con la poca información que se cuenta, en Latinoamérica ya se han iniciado esfuerzos similares en la búsqueda de alternativas para la protección de los ecosistemas de profundidad, y es así como mediante la Resolución 679 del 2005 (mayo 31) el Ministerio de Ambiente, Vivienda y Desarrollo Territorial de Colombia (MAVDT, 2005) declaró el AMP de los Archipiélagos del Rosario y de San Bernardo (ARSB), la cual amplió tanto en área como en disposiciones legales al Parque Nacional Natural Corales del Rosario y de San Bernardo y dio un nuevo carácter de conservación y manejo de la región. Una de las justificaciones importantes para la ampliación de esta área hasta la isóbata de los 200 m de profundidad, estuvo basada en el registro preliminar de comunidades coralinas profundas realizado por Reyes et al. (2005). Por lo cual, estas comunidades fueron incluidas bajo el principio de "precaución" en la declaración del AMP ARSB, dado el limitado conocimiento que se tenía aún de ellas.
Durante las investigaciones realizadas en los proyectos INVEMAR MACROFAUNA I-II, se listaron cerca de 900 especies de peces e invertebrados para el Caribe colombiano (Reyes et al., 2005). El hallazgo de una gran cantidad de corales azooxantelados en tres estaciones, La Guajira, Santa Marta y San Bernardo, con una riqueza de especies mayor que las demás estaciones maestreadas, permitió sugerir que dichas áreas coralinas constituyen comunidades bentónicas diversas en el Caribe colombiano (Reyes et al, 2005; Santodomingo et al., 2007). En la estación ubicada costa afuera del archipiélago de San Bernardo (155-160 m), se encontraron 19 especies de corales escleractinios y una alta diversidad de especies de crustáceos, moluscos, equinodermos y peces asociados. Dichos resultados sugirieron que las comunidades coralinas profundas son importantes ya que en su conjunto sostienen una tercera parte de la biodiversidad marina presente en la plataforma y talud continental del Caribe colombiano (Reyes et al., 2005).
La conservación y el manejo sostenible de los recursos son los dos objetivos principales para el establecimiento de las AMP (Roberts et al., 2003a). Todas las AMP se definen con base en diferentes criterios de selección y metas de conservación. Para la identificación y selección de nuevas AMP se requiere de un proceso de evaluación previo que implica la aplicación de criterios que orienten el proceso adecuadamente (Salm et al., 2000). En el caso del AMP ARSB se seleccionaron 17 criterios con el fin de orientar su delimitación (INVEMAR et al., 2003). El presente estudio pretende identificar cuáles de estos criterios biológicos son evaluables para las comunidades de profundidad, de modo que sirvan como herramienta para la zonificación y declaración de los corales azooxantelados como comunidades estratégicas a conservar en el sistema de AMP en Colombia.
ÁREA DE ESTUDIO
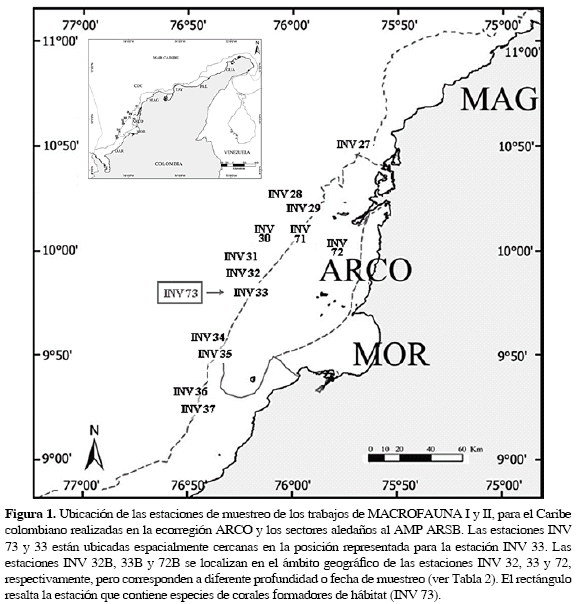
El área de estudio se localiza en el sector suroriental del Caribe colombiano. Los datos fueron obtenidos en 17 estaciones de la ecorregión Archipiélagos Coralinos (ARCO) y sus áreas adyacentes, la cual incluye el AMP ARSB, con profundidades que oscilan entre 70 y 520 m, localizadas entre los 9° 16' 21" N, 76° 29' 37" W y los 10° 28' 07" N, 75° 42' 34" W (Figura 1, Tabla 2). La ecorregión ARCO se ubica paralelamente frente a la ecorregión Golfo Morrosquillo (MOR), a partir de la isóbata de 40 m y hasta el límite externo de la plataforma continental del Caribe colombiano, de acuerdo con la zonificación propuesta por Díaz y Acero (2003). Esta región, que comprende las islas coralinas del Rosario y el archipiélago de San Bernardo, está localizada en una dinámica zona de interacción de tres placas tectónicas, Nazca, Suramericana y Caribe, cuyos desplazamientos generan fenómenos de compresión y cizallamiento que han dado origen a los cinturones de San Jacinto y Sinú. En este sector el diapirismo de lodo aparece como el factor predominante sobre la geomorfología, permitiendo el establecimiento de comunidades coralinas someras (Vernette, 1985; Vernette et al., 1992), y se ha sugerido que estos fenómenos también favorecen la presencia de comunidades de profundidad en el área (Santodomingo et al., 2007).
MATERIALES Y MÉTODOS
Con base en la información de los proyectos MACROFAUNA I y II para el Caribe colombiano, se extrajo un listado de las especies de corales, crustáceos, equinodermos, moluscos y peces recolectadas en 15 estaciones de la ecorregión ARCO y dos estaciones adyacentes localizadas en la ecorregión Darién (DAR), para un total de 17 estaciones de muestreo (Figura 1). Todas las muestras fueron colectadas a bordo del B/I Ancón del INVEMAR, mediante una red demersal (tipo semi-balón, apertura 9 x 1 m) durante 20 min, con una velocidad promedio de 3 nudos (aprox. 5.5 Km/h). Una vez en cubierta las muestras fueron lavadas sobre un tamiz (ojo de malla 2 mm), separadas por grupos taxonómicos principales y posteriormente fijadas en etanol al 70 ó 96 %. En el laboratorio fueron fotografiadas e identificadas por expertos, cuyos principales resultados taxonómicos han sido publicados en Saavedra-Díaz et al. (2000), Lattig y Reyes (2001), Borrero-Pérez et al. (2002a, 2002b), Cruz et al. (2002), Gracia et al. (2002), Roa-Varón et al. (2003), Borrero-Pérez y Benavides-Serrato (2004), Campos et al. (2004), Gracia et al. (2004), Reyes et al. (2005).
En el análisis de la estructura de la comunidad presente en el archipiélago de las islas del Rosario y San Bernardo, se emplearon indicadores ecológicos tales como las técnicas univariadas riqueza de Margalef, uniformidad de Pielou, diversidad de Shannon-Wiener y predominio de Simpson con base en los datos de abundancia de especies (Gray, 2000), con el fin de establecer si los sitios con corales de profundidad difieren de otros sectores de la plataforma. En dichos análisis de biodiversidad se consideró la abundancia de las especies de moluscos, peces, crustáceos, equinodermos y cnidarios; dentro de este último grupo se excluyeron las abundancias de las especies de coral Anomocora fecunda, Eguchipsammia cornucopia, Madracis asperula, Madracis myriaster y Madrepora carolina, debido a que éstas se tuvieron en cuenta como formadores de hábitat y sustento para las demás especies de invertebrados y peces. Además se realizó un análisis de clasificación aplicando el índice de similaridad de Bray-Curtis, con base en datos de abundancia transformados log (x+1), y posteriormente se efectuó un análisis de ordenación mediante la técnica del NMDS (Non-metric Multidimensional Scaling; Clarke y Warwick, 2001).
Criterios biológicos
Los criterios biológicos para el establecimiento de las AMP han sido propuestos y compilados en las publicaciones de Salm y Clark (1984), Kelleher y Kenchington (1991), Kelleher et al. (1995), Roberts y Hawkins (2000), Salm et al. (2000), Airamé et al. (2003) y Roberts et al. (2003a, 2003b). Con base en esta información y la presentada en el informe técnico del INVEMAR et al. (2003), se escogieron los criterios biológicos que pueden ser aplicados a las comunidades de corales azooxantelados en el marco del establecimiento del AMP en cuestión. Como punto de partida se escogieron los criterios de diversidad, naturalidad, representación y heterogeneidad de hábitats, calidad de único, presencia de poblaciones o especies de interés especial, presencia de etapas de vida vulnerables, vulnerabilidad, productividad, de exclusión y de selección (conectividad). De esta manera, con base en los valores de diversidad e importancia ecológica, y con la información secundaria recopilada para el área se describió cada uno de los criterios preestablecidos y se determinó cuáles de éstos pueden ser usados como herramienta para priorizar el (los) sector(es) más importante(s) en el área de estudio que conlleven a la preservación de estas comunidades. En este caso, para los criterios identificados también se tuvieron en cuenta las posibles amenazas, factibilidad, influencias, protección existente, entre otras. Por último, se identificaron los vacíos de información en el conocimiento de estas comunidades y sobre los cuales sería interesante realizar nuevas investigaciones que soporten el manejo y conservación de estos ecosistemas únicos.
RESULTADOS Y DISCUSIÓN
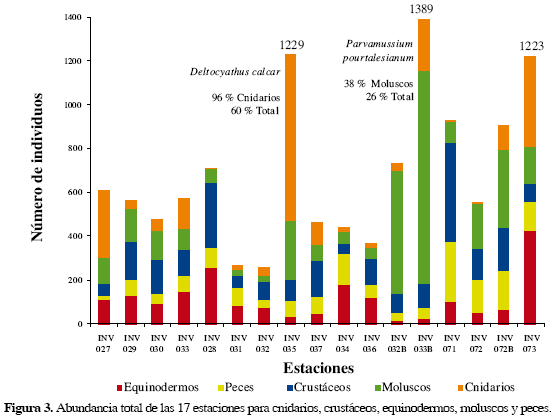
Se recolectaron un total de 11685 ejemplares de todos los taxa, distribuidos en 528 especies, pertenecientes a 238 familias. En general, la composición observada fue la siguiente (Figuras 2 y 3):
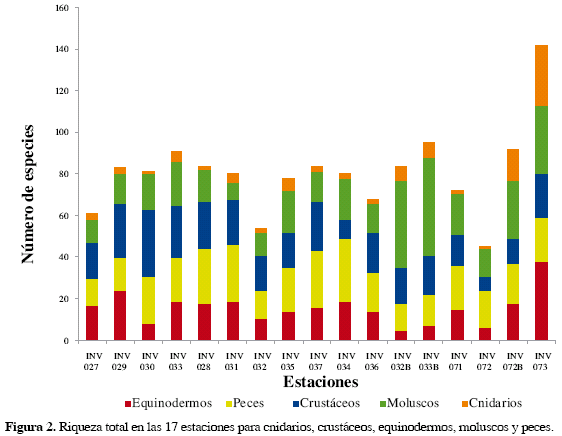
Los moluscos fueron el grupo con mayor cantidad de familias (81) y especies (154). Las familias Cocculinidae (ocho especies) y Turridae (ocho especies) fueron las de mayor riqueza. Los peces representaron el segundo grupo con mayor abundancia y riqueza con 63 familias y 129 especies, siendo la familia Scorpaenidae con 11 especies, la de mayor riqueza. Los crustáceos presentaron 110 especies distribuidas en 40 familias, dominadas principalmente por Galatheidae (16 especies) Los equinodermos presentaron 39 familias y 97 especies, predominando la familia Ophiuridae con 13 especies. Por último, los cnidarios (no formadores de arrecife), presentaron los menores valores en cuanto al número de especies (38) y familias (15); la familia Caryophylliidae se destacó significativamente de las otras porque está representada con al menos una especie en todas las estaciones.
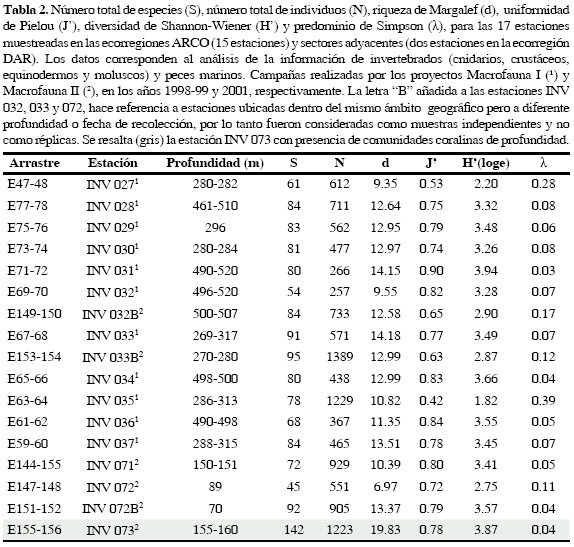
La riqueza observada en la estación INV 073 (Figura 2), fue significativamente mayor, caracterizada por la presencia de corales de profundidad formadores de hábitat, presentando el número de especies más alto (142 en total). Adicionalmente, se destaca que dicha estación presentó más del 50 % de especies en comparación con las demás estaciones, en las cuales el número de especies osciló entre 45 (INV 072) y 95 especies (INV 033B). Igualmente, en INV 073 ninguna especie fue dominante siendo así la estación más heterogénea (Tabla 2).
La abundancia (Figura 3) fue mayor en las estaciones INV 033B, 035 y 073 con 1389, 1229 y 1223 individuos, respectivamente. Los primeros dos valores resultan de la dominancia de las especies Parvamussium pourtalesianum (Mollusca: Pectinidae) y Deltocyathus calcar (Scleractinia: Caryophylliidae), respectivamente. Las estaciones INV 33B (λ = 0.12) e INV 35 (λ = 0.39) mostraron índices de predominio altos comparados con las otras estaciones, siendo la última la que presentó un mayor predominio (Tabla 2). Parvamussium pourtalesianum presenta 367 individuos que representan el 38 % del grupo de los moluscos y el 26 % del total de individuos colectados en la estación INV 033B. El bivalvo P. pourtalesianum es uno de los más representativos componentes de la fauna de fondos blandos del Caribe colombiano y es especialmente abundante en la franja de los 300 y 500 m (Gracia, 2000). De otra parte, el coral solitario D. calcar (Scleractinia: Caryophylliidae), tuvo el valor más alto de abundancia (734 individuos), representando el 96 % de los cnidarios y el 60 % de del total en la estación INV 035; D. calcar pertenece a la familia Caryophylliidae y presenta un amplio ámbito batimétrico entre los 80 y 675 m (Cairns, 1979), y aunque esta especie sólo era conocida en las islas del Rosario (Prahl y Erhardt, 1989), durante las expediciones de MACROFAUNA se recolectó en todo el Caribe colombiano. De las 26 especies de corales escleractínios que se encontraron en estos cruceros, D. calcar presentó el ámbito de distribución más amplio corroborando lo registrado por Lattig (2000) De esta manera, la dominancia de estas dos especies respectivamente en las dos estaciones de mayor abundancia de individuos (INV 033B y 035), evidenció que aún en un tercer lugar, la estación INV 073 presentó una mayor uniformidad en la abundancia relativa de todos los componentes de la fauna.
Los índices de diversidad registrados en este estudio son bajos (H'= 1.82 a 3. 87) si se comparan con datos de diversidad del mar profundo en otras localidades, tales como las clásicas investigaciones en la plataforma y talud de la costa este de Estados Unidos realizadas por Grassle y Maciolek (1992), quienes registran valores de H'= 5.6 a 6, y el de Levin et al. (2001), con diversidad H'= 4.09 a 6.7. Sin embargo, si se compara con estudios de diversidad de arrecifes coralinos someros en el Caribe, se observa que las comunidades profundas de la región estudiada presentan valores dentro del mismo ámbito de variación (p. ej. Porter, 1972, donde H'= 1 a 3.8). Dentro de las estaciones evaluadas, es de resaltar que los mayores valores de diversidad fueron observados para las comunidades coralinas profundas representadas por INV 073, que presentó valores de 3.87 en el índice de Shannon, 19.83 en la riqueza de Margalef y de 0.78 en la uniformidad de Pielou; de esta misma manera, esta estación presenta uno de los índices más bajos de predominio de Simpson (λ = 0. 04), siendo únicamente menor este mismo índice en la estación INV 031 (λ = 0.03) (Tabla 2). Dichos valores de diversidad, riqueza y predominio, sugieren que la estación INV 073 no sólo es la más diversa. Este resultado podría explicarse por la presencia de corales de profundidad formadores de hábitat, que favorecen la presencia de los demás grupos de invertebrados y peces.
De otra parte, aunque la estación INV 031 también presenta altos valores de riqueza de Margalef (3.94) y uniformidad de Pielou (0.90), si se comparan la riqueza específica (80 vs. 142) y el número de individuos (266 vs. 1223) con respecto a la estación INV 073 es significativamente más bajo (Tabla 2). De esta manera, se puede considerar que la estación INV 073 es la de mayor importancia ecológica con base en los índices estimados (Tabla 2).
El análisis de clasificación revela que existe un gradiente definido por profundidad formando tres grupos, con cerca del 30 % de similaridad (Figura 4): las estaciones de 500 m, las de 300 m y las localizadas en aguas más someras por encima de los 200 m de profundidad (plataforma continental). En el NMDS se observó de una manera más clara el mismo patrón de agrupación (Figura 5). El grupo de 500 m está compuesto por seis estaciones y se caracterizó por la presencia de Nezumia aequalis (Macrouridae), Pleoticus robustus (Solenoceridae) y Poromya margarita (Poromyidae), exclusivas de esta profundidad. El grupo de 300 m está conformado por siete estaciones, y se caracteriza porque las especies D. calcar y P. pourtalesianum representan en conjunto más del 97 % de la abundancia total en relación a todas las estaciones y por la presencia de las especies exclusivas Metanephrops binghami (Nephropidae), Neobythites gilli (Ophidiidae) y Plesionika tenuipes (Pandalidae) en seis de las siete estaciones. En el análisis de ordenación este grupo tiende a ubicarse en la parte inferior del gráfico (Figura 5). Sin embargo, la estación INV 033B aparece alejada del grupo, posiblemente por el alto valor de abundancia de la especie P. pourtalesianum Este patrón de agrupación por profundidades concuerda con los obtenidos por Gracia (2000), Benavides-Serrato y Borrero-Pérez (2000), Roa (2000) y Cruz (2001), en taxa particulares como crustáceos, moluscos, equinodermos y peces en los fondos blandos del Caribe colombiano.
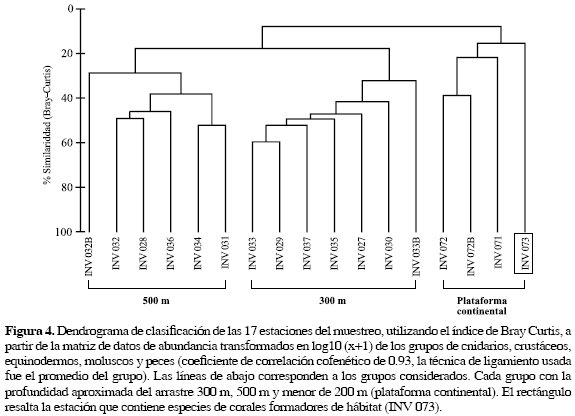
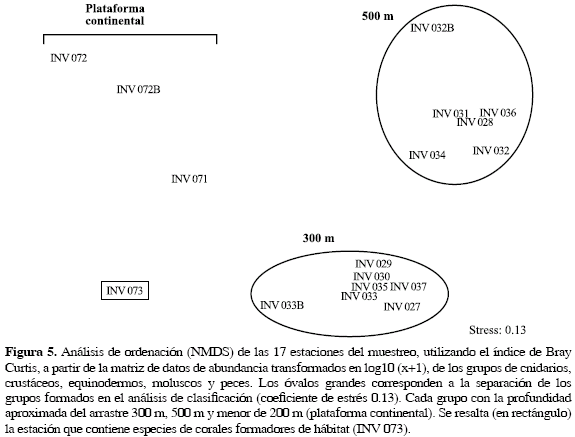
De otra parte, las estaciones localizadas en la plataforma continental no presentaron ningún tipo de agrupación. En el análisis de ordenación tampoco hay evidencia de la formación de un grupo para las estaciones de la plataforma (INV 072, 072B, 071, 073; Figura 5). No obstante, aquellas por encima de los 100 m pueden conformar un subgrupo como el que se sugiere en el dendrograma, conformando por las estaciones INV 072 y 072B, las cuales además se encuentran cercanas espacialmente y en el mismo ámbito de profundidad (Tabla 2). La estación INV 071 no se agrupa con las anteriores posiblemente por su diferencia en la profundidad y no se acerca a INV 073 porque únicamente comparten el 10 % de las especies.
Ahora bien, se sugiere que la estación INV 073 (Figura 5, rectángulo), al estar ubicada en el borde de la plataforma continental, se encuentra en una zona privilegiada de intercambio de ambientes ofrecidos por los sustratos del talud y de la plataforma continental, lo cual incrementa su importancia ecológica. Esta ubicación facilitaría el proceso asentamiento de larvas de corales formadores de hábitat, favoreciendo la alta diversidad de especies asociadas como se ha encontrado en otras latitudes (Krieger, 2001; Tunesi et al., 2001; Freiwald, 2003). Además, cabe resaltar que INV 073 presenta exclusivamente un poco más del 25 % de las especies que no se encontraron en ninguna otra estación, considerándose que ella representa un ambiente particular dentro de los fondos analizados. De esta manera, se puede sugerir que los corales azooxantelados formadores de hábitat constituyen la matriz para el desarrollo de una amplia diversidad de especies de invertebrados y peces en el área analizada.
Criterios biológicos
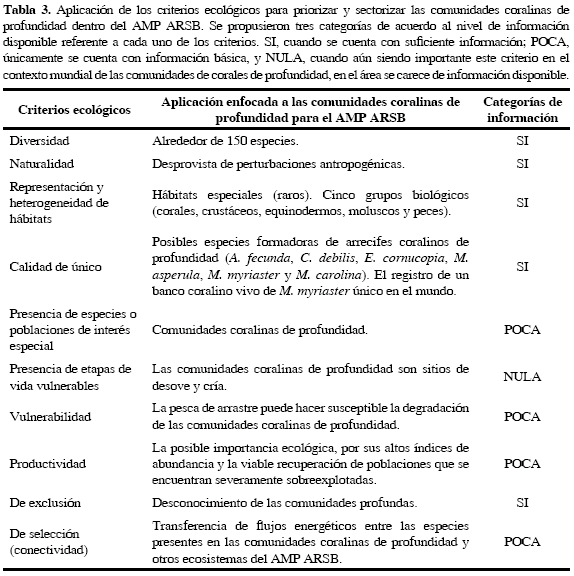
Luego de analizar los diferentes índices de diversidad, abundancia de especies y patrones de similaridad que destacan la importancia de la presencia de corales azooxantelados formadores de hábitat en el área de estudio, se procedió a enmarcar dichos datos ecológicos en el contexto de los conceptos teóricos establecidos para la definición de AMP en Colombia. Aunque no fue posible sustentar con los datos de este estudio todos los criterios escogidos, se discute la importancia de cada uno de ellos en el marco del conocimiento que se tiene del papel que desempeñan las comunidades coralinas de profundidad en otras regiones del mundo y se enfatiza la necesidad de realizar investigaciones encaminadas a suplir los vacíos de información encontrados. Sus características se resumen en la Tabla 3.
Diversidad. El criterio biológico de diversidad es uno de los más importantes al momento de definir las AMP, ya que el esquema de priorización se basa específicamente en la conservación de la biodiversidad incluyendo sitios con una alta riqueza de especies (Roberts et al., 2002). Con base en este criterio, se busca conseguir una mayor representación de los ecosistemas, hábitats, comunidades y especies. En este sentido, las comunidades coralinas de profundidad dentro del AMP ARSB juegan un papel fundamental en el desarrollo de las especies marinas en términos de diversidad. Estas comunidades coralinas están representadas por la estación INV 073, ubicada en el borde de la plataforma continental, la cual presenta el mayor número de especies (142) y altos valores de diversidad (Shannon-Wiener 3.87), riqueza (Margalef 19.83) y una significativa uniformidad (Pielou 0.78). Es así como se justifica que las comunidades coralinas de profundidad sean calificadas con un nivel de importancia mayor entre las de fondos de la plataforma y talud continental, y se sugiere que estos ecosistemas se incluyan como prioritarios a conservar dentro de las normativas de protección del AMP ARSB.
Naturalidad. Cuando los sistemas naturales marinos no presentan degradaciones antropogénicas significativas se favorece el desarrollo biológico de las especies. Podría considerarse que los corales de profundidad y su diversa fauna acompañante en Colombia aún se encuentran en un estado casi prístino, ya que la pesca de arrastre, que es una de las principales causas de los mayores daños directos en las profundidades (Shester y Ayers, 2005), está relativamente poco desarrollada en el país. Sin embargo, recientes estudios demuestran que es posible que el impacto causado por las pesquerías de arrastre en el Caribe colombiano sobre las comunidades de invertebrados marinos haya sido subestimado hasta la fecha (García et al., 2008), y que la "naturalidad" de estos ecosistemas esté en riesgo. Este aspecto se profundizará bajo el criterio de vulnerabilidad.
Representación y heterogeneidad de habitats. La necesidad que todos los hábitats marinos estén representados dentro de una región biogeográfica hace que los corales de profundidad (hábitats raros), sean contemplados con especial importancia, y se les brinde protección adicional y la garantía de la inclusión como un objeto de conservación primordial dentro de la misma (Roberts et al., 2003b; Lourie y Vincent, 2004). Hace menos de una década se sabe de la presencia de comunidades coralinas de profundidad en Colombia (Santodomingo y Reyes, 2003; Reyes et al., 2005), lo cual hace inaplazable la tarea de proteger estos sitios especiales. La incorporación de estas comunidades coralinas profundas, en conjunto con otros ecosistemas como los arrecifes coralinos someros, las praderas de fanerógamas, playas, fondos blandos y los manglares, crea un mosaico único de ecosistemas que incrementa de manera significativa el nivel de representatividad y heterogeneidad en el AMP ARSB. En algunas ocasiones, el uso de este criterio es un mejor indicador de biodiversidad que la riqueza de especies en sí misma, debido a las diferencias intrínsecas de cada hábitat. Por ejemplo, aunque los arrecifes coralinos siempre tienen una riqueza de especies mayor que las playas arenosas, no son más valiosos para la conservación que estas últimas, ya que ambos ecosistemas presentan características ambientales contrastantes y especies únicas, que al presentarse juntos en un mismo sector elevan su valor para la conservación (Roberts et al., 2003b).
Calidad de único. Este criterio contempla varias categorías, una de ellas es la de las especies o poblaciones que tienen características particulares que las hacen de gran interés para la inclusión dentro de un AMP (Roberts et al., 2003a). Para este caso particular, se hace referencia a los corales de profundidad como comunidades únicas para el Caribe colombiano (Reyes et al., 2005), que presentan más de un 25 % de especies exclusivas asociadas. Dentro de las características ambientales que las hacen ser particulares, se puede citar por ejemplo, su desarrollo en aguas con temperaturas menores a 20 °C, en ambientes de fuertes corrientes marinas con alto contenido de materia orgánica para su alimentación, la tolerancia a la sedimentación y a salinidades mayores a 35, entre otras. Estas condiciones hacen únicas a las comunidades coralinas de profundidad, y las diferencian de sus contrapartes someras, los arrecifes coralinos conformados por corales zooxantelados, los cuales requieren de aguas cálidas, transparentes, relativamente tranquilas y pobres en nutrientes para su buen desarrollo (Bellwood et al., 2004). Además, este hallazgo corresponde al único registro conocido de un banco coralino vivo de la especie Madracis myriaster en el mundo, en contraste con comunidades fósiles de Madracis que han sido datadas del Holoceno en los sectores aledaños a la desembocadura del río Níger en Guinea, en Atlántico oriental (Allen y Wells, 1962). Por estas razones, su condición de únicas representa uno de los criterios que otorga un mayor valor ecológico a estas comunidades
Presencia de especies o poblaciones de interés especial. Una de las metas para las AMP es la protección de las poblaciones con un interés especial, las cuales incluyen las especies de importancia comercial, especies clave, amenazadas y/o en peligro. En este caso, los objetos de conservación "bandera" propuestos en este estudio incluyen principalmente los corales formadores de hábitat (Anomocora fecunda, Eguchipsammia cornucopia, Madracis asperula, M. myriaster y Madrepora carolina) que se encuentran contemplados en el apéndice II de CITES, que incluye además todas las especies del orden Scleractinia. Es importante resaltar que, aunque estos corales se encuentran bajo reglamentación de comercio, no sobran los esfuerzos al incluirlos dentro de este criterio, ya que sin un control eficaz, podrían llegar a ponerse en peligro (Reyes y Santodomingo, 2002). De otra parte, es notorio que los ecosistemas profundos no han sido ampliamente estudiados para Colombia (Reyes et al., 2005, Santodomingo et al., 2007), en comparación con el conocimiento que se tiene de ellos en el ámbito mundial. Esto se debe a la mayor disponibilidad de recursos, tecnologías y capacitación disponibles en los países desarrollados para este tipo de investigaciones, los cuales han sacado a la luz la importancia ecológica de estos ecosistemas, en contraste con los estudios aún pioneros que se han desarrollado en Colombia.
Presencia de etapas de vida vulnerables. Diferentes estudios realizados sobre poblaciones de peces comerciales como pargos (Lutjanus spp.), chernas y meros (Serranidae), han demostrado que las comunidades coralinas de profundidad son hábitats fundamentales durante el desove, reclutamiento y cría de estas especies (Sauer et al., 1992; Reed et al., 2005). De este modo, las comunidades coralinas profundas desempeñan un rol vital para muchas especies, debido a que la mayoría de las especies marinas son particularmente susceptibles a la mortalidad en sus primeras etapas de vida y por tanto son las más críticas para el desarrollo poblacional. Aunque es bien conocido este papel en comunidades coralinas profundas de otras regiones (Morgan et al., 2005), la información obtenida hasta la fecha para el Caribe colombiano no es lo suficientemente detallada como para dar soporte a este criterio. Por lo tanto, se incluye en este documento como un vacío de información que debe ser tenido en cuenta por futuras investigaciones.
Vulnerabilidad. Las comunidades coralinas de profundidad en el AMP ARSB son vulnerables principalmente al impacto "subestimado" de las pesquerías de arrastre en la región y al poco conocimiento que se tiene de estas comunidades en el país. Pocos estudios han valorado el porcentaje que representa la fauna acompañante o "by-catch", de invertebrados marinos en las pesquerías de arrastre que actualmente se desarrollan en el sector aledaño al AMP ARSB, aunque una primera aproximación evidencia que el impacto de estas actividades pesqueras podría ser alto (García et al., 2008). Entre los grupos de pesca incidental figuran no sólo invertebrados móviles, como moluscos y crustáceos, sino también organismos sésiles como octocorales y esponjas (García et al., 2008). Algunos estudios puntuales para el Caribe colombiano han registrado valores de la proporción peces/camarón de 10:1 (Osorio, 1986) y 12:1 (Mora, 1989), lo cual evidencia el fuerte impacto que producen las redes de arrastre sobre la biodiversidad (Medina, 2002; Viaña et al., 2004; Zúñiga et al., 2004).
Esta amenaza se hace más latente, en la medida en que los planes de expansión de las pesquerías de arrastre en Colombia contemplan su implementación hasta profundidades de 500 m (Manjarrés et al., 2006). En este punto es importante mencionar, que el hallazgo de comunidades coralinas de profundidad se ha hecho de modo incidental en Colombia (Reyes et al., 2005; Santodomingo et al., 2007), ya que las características de los fondos donde se desarrollan no son detectables con la tecnología básica con que cuentan las pesquerías convencionales. De esta manera, es fundamental destacar la vulnerabilidad de estas comunidades y asegurar su representación en el AMP ARSB, con el fin de evitar graves e irreversibles desastres para la diversidad colombiana. Al igual que se ha establecido en otras regiones del mundo, la pesca de arrastre ha diezmado la alta biodiversidad asociada a corales profundos (Shester y Ayers, 2005), y por lo tanto debe ser restringida en las áreas donde haya evidencias de la presencia de estas comunidades. Aunque es prematuro prever si la actual prospección de hidrocarburos en el Caribe colombiano (Carbó, 2008) resultará en la instalación de plataformas en áreas profundas con presencia de corales azooxantelados, y que esto a su vez causará un efecto negativo para su diversidad asociada, con base en las experiencias negativas de otros países (Davies et al., 2007) es importante acompañar de manera preventiva a la industria petrolera y advertir de los posibles daños.
Productividad. La economía pesquera para cualquier país es importante ya que de estos recursos dependen muchas personas. La protección de poblaciones potencialmente explotables es un prerrequisito para las AMP y para el beneficio de los habitantes que circundan estas zonas. Por tal motivo, el grado de protección para un área que incremente los stocks de importancia comercial debe ser alto, más aún si apoya la recuperación de poblaciones que se encuentran severamente afectadas (Halpern, 2003). En el caso del área aledaña al AMP ARSB donde se pesca industrialmente al camarón Farfantepenaeus notialis (García et al., 2008) y artesanalmente numerosos grupos de peces de arrecifes (pargos, meros, chernas, entre otros; Patiño y Flórez, 1993), con la creación del AMP se propende por incrementar estos beneficios y simultáneamente proteger la biodiversidad. Para dar sustento a este criterio, se requiere la realización de más investigaciones y producir información contundente Por lo pronto, los datos presentados en este documento incluyen indicios de que las comunidades coralinas de profundidad del Caribe colombiano aportarían a estos beneficios productivos, debido principalmente a los altos índices de biodiversidad encontrados, y a la presencia de algunas de las familias con especies de importancia comercial como Serranidae, Lutjanidae, Carangidae y Trichiuridae, así como de crustáceos de las familias Penaeidae y Portunidae, grupos que han sido reconocidos con potencial para su explotación (Carpenter, 2002; Tavares, 2002).
De exclusión. Con la información presentada y discutida anteriormente, la amenaza antropogénica hacia las comunidades coralinas de profundidad localizadas por debajo de los 150 m de profundidad es todavía baja, debido a que en el Caribe colombiano no hay una explotación pesquera de arrastre en profundidades mayores a los 100 m. Por lo tanto, proteger las zonas donde se tenga evidencia de estas comunidades permitirá que se mantengan en condiciones prístinas de desarrollo. Esto contribuirá al mejor entendimiento de su papel ecológico y al diseño de planes de manejo. Así, se garantizaría su protección ante posibles desastres antropogénicos que afectarían directamente la biodiversidad del AMP ARSB. Adicionalmente, este criterio implica que se tengan en cuenta como potenciales de conservación no sólo en el área sino en todo el Caribe colombiano, especialmente porque su protección reduce amenazas futuras anticipadas (Roberts et al., 2003a), como las mencionadas prospecciones pesqueras y de hidrocarburos en el Caribe colombiano que ya fueron discutidas bajo el criterio de vulnerabilidad.
De selección (conectividad). En este contexto, conectividad se define como la transferencia de energía entre diferentes lugares de una reserva (Roberts y Hawkins, 2000), representada por el aporte de biomasa desde las comunidades coralinas de profundidad hacia otros ecosistemas del AMP ARSB, ampliando la biodiversidad poblacional del lugar (e.g. movimientos de juveniles y adultos), en conjunto con los otros ecosistemas del AMP tales como, arrecifes coralinos someros, praderas de pastos marinos y fondos blandos. Este criterio hace referencia a la interconexión que existe entre los diferentes ecosistemas que se presentan en un AMP, ya que algunas especies con rangos batimétricos amplios pueden pasar ciertas etapas de su vida en cada uno de estos ambientes. Algunas de las especies comunes encontradas entre las comunidades coralinas profundas y los ecosistemas someros que ejemplifican esta conectividad son el ofiuro Ophiothrix suensonni, los cangrejos pertenecientes a los géneros Mithrax y Stenorhyncus, y peces como Synagrops bella y Antigonia capros.
Con el registro de especies coralinas de profundidad formadoras de hábitat, que conforman un ensamblaje con una alta biodiversidad asociada de especies, se propone priorizar y sectorizar las áreas que albergan estas comunidades dentro del AMP ARSB, basados en los criterios ecológicos establecidos en este documento. Tal como se ha discutido, de los diez criterios contemplados se dispone de suficiente información para cinco criterios, poca información para cuatro de ellos y ninguna para el criterio de "presencia de etapas de vida vulnerables" (Tabla 3). Es así como se propone continuar con los estudios que permitan llenar los vacíos de información existentes y así equiparar el nivel de conocimiento que se tiene de estas importantes comunidades en Colombia con respecto al alcanzado en otros países, donde los corales de profundidad son prioridades de conservación (Roberts et al., 2009).
Aunque sería ideal poder proteger el 100 % de los ecosistemas coralinos de profundidad esta tarea requiere de muchos esfuerzos (Margules y Pressey, 2000); adicional a esta situación, el conocimiento que se tiene de estas comunidades en Colombia es aún incipiente. Por lo tanto, se recomienda el uso de otras herramientas que permitan generar una modelación para su conservación, por ejemplo el programa MARXAN, el cual genera posibles sitios prioritarios de conservación en los que se incluyen todo tipo de objetos de conservación (atributos ecológicos y presencia de especies). En Colombia, se están produciendo avances en el diseño de AMP en el Caribe continental y creando perspectivas a futuro por medio de estas metodologías de planificación (Alonso, 2006; Alonso et al., 2008a, 2008b). En este contexto se recomendaría darle una prioridad del 100 % a estos sistemas profundos al momento de incluirlos en dichas modelaciones, debido a la importancia que éstos representan para las especies marinas.
Como resultado de la evaluación de las comunidades de corales de profundidad bajo los diferentes criterios biológicos de conservación, se considera que es necesaria la inclusión de éstos en el AMP ARSB, siendo recomendable que se tomen como objetos de conservación antes que sean alterados antropogénicamente. También es importante que se dirijan esfuerzos en la investigación y el mapeo de las zonas profundas en búsqueda de consolidar el conocimiento adquirido en este estudio.
CONCLUSIONES
Se identificaron diez criterios biológicos evaluables para las comunidades coralinas de profundidad del AMP ARSB, de los cuales se cuenta con suficiente información para cinco de ellos: diversidad, naturalidad, representación y heterogeneidad de hábitats, calidad de único y de exclusión. De esta manera, se pudo establecer que las comunidades coralinas de profundidad se encuentran en un estado prístino, albergan especies de importancia ecológica (un banco coralino vivo de Madracis myriaster único en el mundo) y sustentan una amplia diversidad biológica (fauna acompañante).
A pesar de la poca información disponible sobre los criterios biológicos relacionados con la presencia de especies o poblaciones de interés especial, la vulnerabilidad, productividad y de selección (conectividad), estos criterios deben ser tenidos en cuenta en futuros estudios con el fin de evaluar la importancia de las comunidades coralinas de profundidad en un contexto ecológico, evolutivo y productivo más amplio. Aunque no se cuenta con información para evaluar el criterio biológico que hace referencia a la presencia de etapas de vida vulnerables, ya que se requieren de metodologías más detalladas y avanzada tecnología, con base en estudios análogos en otras regiones del mundo, se plantea la necesidad de llenar el vacío de información que se tiene acerca del papel que particularmente podrían desempeñar las comunidades coralinas profundas como sitios de cría y desove de especies marinas en el AMP ARSB.
Finalmente, se pudo establecer que las comunidades coralinas de profundidad del AMP ARSB son estructuras fundamentales para el desarrollo de una alta diversidad de especies marinas en la región. Gracias a las evidencias aquí planteadas debe incentivarse su inclusión dentro de los planes de manejo, no sólo bajo el principio de precaución, sino promoverlas como una comunidad igual de importante a los demás ecosistemas que hacen parte del AMP ARSB.
AGRADECIMIENTOS
Este estudio fue posible gracias al apoyo brindado por el INVEMAR y Colciencias a través del proyecto MARCORAL (código 2115-09-16649) y a nuestros compañeros de equipo A. Gracia, A. Martínez, N. Cruz, P. Flórez, I. Daniel y C. García. A los investigadores que participaron en los proyectos MACROFAUNA I y II (proyectos 210509-10401 y 210509-11248), G. Borrero-Pérez, M. Benavides, A. Roa, L. Saavedra, P. Lattig, N. Ardila, L. S. Mejía, L. M. Mejía, P. Rachello, P. Ariza y A. Polanco. Agradecimientos especiales a G. Navas por sus incontables esfuerzos en la realización de investigaciones marinas de profundidad en Colombia. Igualmente gracias a M. Vides, D. Alonso, C. Segura-Quintero y A. Báez por su asesoría y valiosa ayuda en el análisis de la información. Los comentarios y sugerencias del profesor A. Sanjuán y otros dos evaluadores anónimos enriquecieron enormemente el contenido de este documento. N. Santodomingo agradece el apoyo brindado por el programa Alpan (código E07M402757CO).
BIBLIOGRAFÍA
1 Airamé, S., J. E. Dugan, K. D. Lafferty, H. Leslie, D. A. McArdle y R. R. Warner. 2003. Applying ecological criteria to marine reserve design: a case study from the California Channel Islands. Ecol. Appl., 13 (1): 170-184. [ Links ]
2 Allen, J. R. L. y J. W. Wells. 1962. Holocene coral banks and subsidence in the Niger Delta. J. Geol., 70 (4): 381-397. [ Links ]
3 Alonso, D. 2006. Avances en el diseño de áreas marinas protegidas del Caribe continental y perspectivas a futuro. Taller biodiversidad del margen continental: comunidades coralinas de profundidad. Informe técnico, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta. 39 p. [ Links ]
4 Alonso, D., L. F. Ramírez, C. Segura-Quintero, P. Castillo-Torres, T. Walschburger y N. Arango. 2008a. Hacia la construcción de un Subsistema Nacional de Áreas Marinas Protegidas en Colombia. INVEMAR, UAESPNN y TNC, Santa Marta. 20 p. [ Links ]
5 Alonso, D., C. Segura-Quintero, P. Castillo-Torres y J. Gerhantz-Muro. 2008b. Avances en el diseño de una red de áreas marinas protegidas: estrategia de conservación para el norte del Caribe continental colombiano. Bol. Invest. Mar. Cost., 37 (1): 129-156. [ Links ]
6 Bellwood, D. R., T. P. Hughes, C. Folke y M. Nyström. 2004. Confronting the coral reef crisis. Nature, 429: 827-833. [ Links ]
7 Benavides-Serrato, M. y G. Borrero-Pérez. 2000. Equinodermos de la franja superior del talud continental del Caribe colombiano. Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Santa Marta. 527 p. [ Links ]
8 Borrero-Pérez, G. H. y M. Benavides-Serrato. 2004. New record of Ophiosyzygus disacanthus Clark, 1911 (Echinodermata: Ophiuroidea: Ophiomyxidae) in the Caribbean Sea. Proc. Biol. Soc. Washington, 117 (4): 541-544. [ Links ]
9 Borrero-Pérez, G. H., M. Benavides-Serrato, O. Solano y G. Navas. 2002a. Equinoideos (Echinodermata: Echinoidea) colectados en la franja superior del talud continental del Caribe colombiano. Bol. Inst. Inv. Mar. Cost., 31: 133-166. [ Links ]
10 Borrero-Pérez, G. H., O. D. Solano y M. Benavides-Serrato. 2002b. Lista revisada de los erizos (Echinodermata: Echinoidea) del mar Caribe colombiano. Biota Colombiana, 3: 137-144. [ Links ]
11 Butler, M. 2005. Conserving in Atlantic Canada: a historical perspective. 1199-1209. En: Freiwald, A. y J. M. Roberts (Eds.). Cold-water corals and ecosystems. Springer, Berlin, Heidelberg. 1243 p. [ Links ]
12 Cairns, S. D. 1979. The deep-water Scleractinia of the Caribbean Sea and adjacent waters. Studies on the fauna of Curacao and other Caribbean islands, 57 (108): 1-341. [ Links ]
13 Campos, N. H., G. R. Navas, A. Bermúdez y N. Cruz. 2004. Los crustáceos decápodos de la franja superior del talud continental (300-500 m) del mar Caribe colombiano. Monografría Fauna Colombia 2. Instituto de Ciencias Naturales, Museo de Historia Natural, Univ. Nacional de Colombia, Bogotá. 180 p. [ Links ]
14 Carbó, J. 2008. Colombia: oil and gas drilling equipment. The U.S. Commercial. http://www.ado.alabama.gov/content/media/publications/trade/mission/COL_Oil-Gas_Drilling_Equip_112008.pdf. 08/10/2009. [ Links ]
15 Carpenter, K. E. 2002. The living marine resources of the Western Central Atlantic. FAO species identification guide for fishery purposes and American Society of Ichthyologists and Herpetologists special publication. FAO, Roma, (2-3): 601-2127. [ Links ]
16 Clarke, K. R. y R. M. Warwick. 2001. Change in marine communities: an approach to statistical analysis and interpretation. Segunda edición, PRIMER-E, Plymouth, Reino Unido. 172 p. [ Links ]
17 Cruz, N. 2001. Caracterización de los crustáceos decápodos de la franja superior del talud continental (300 - 500 m prof.) en la parte sur del Caribe colombiano. Tesis Biol., Univ. Nacional de Colombia, Santa Marta. 354 p. [ Links ]
18 Cruz, N., A. Bermúdez, N. H. Campos y G. R. Navas. 2002. Los camarones de la Familia Crangonidae del talud continental entre 200 y 500 m del mar Caribe colombiano. Bol. Inv. Mar. Cost., 31: 183-203. [ Links ]
19 Davies, A. J., J. M. Roberts y J. Hall-Spencer. 2007. Preserving deep-sea natural heritage: emerging issues in offshore conservation and management. Biol. Conserv., 138 (3-4): 299-312. [ Links ]
20 Díaz, J. M. y A. Acero. 2003. Marine biodiversity in Colombia: achievements, status of knowledge, and challenges. Gayana, 67 (2): 261-274. [ Links ]
21 Freiwald, A. 2003. Reef-forming cold-water corals. 365-385. En: Wefer, G., D. Billet, D. Hebbeln, B. B. Jørgensen, M. Schlüter y T. C. E. van Weering (Eds.). Ocean margin systems. Springer-Verlag, Berlin. 505 p. [ Links ]
22 García, C. B., D. Pérez, L. O. Duarte y L. Manjarrés. 2008. Experimental results with a reducing device for juvenile fishes in a tropical shrimp fishery: impact on the invertebrate bycatch. Pan-American J. Aquat. Sci., 3 (3): 275-281. [ Links ]
23 Gracia, M. A. 2000. Taxocenosis de los moluscos epibentónicos de la franja superior del talud continental (300 y 500 m) entre Punta Gloria y Cabo Tiburón. Caribe colombiano. Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Santa Marta. 382 p. [ Links ]
24 Gracia, A., N. E. Ardila y J. M. Díaz. 2002. Cefalópodos (Mollusca: Cephalopoda) del talud superior del Caribe colombiano. Bol. Inst. Inv. Mar. Cost., 31: 219-238. [ Links ]
25 Gracia, A., N. E. Ardila y J. M. Díaz. 2004. Gastropods collected along the continental slope of the Colombian Caribbean during the INVEMAR-macrofauna campains (1998-2001). Iberus, 22: 43-75. [ Links ]
26 Grassle, J. F. y N. J. Maciolek. 1992. Deep-sea species richness: regional and local diversity estimates from quantitative bottom samples. Am. Nat., 139 (2): 313-341. [ Links ]
27 Gray, J. S. 2000. The measurement of marine species diversity, with an application to the benthic fauna of the Norwegian continental shelf. J. Exp. Mar. Biol. Ecol., 250 (1, 2): 23-49. [ Links ]
28 Halpern, B. S. 2003. The impact of marine reserves: do reserves work and does reserve size matter? Ecol. Appl., 13: 117-137. [ Links ]
29 Hatcher, B. G. y R. E. Scheibling. 2001. What determines whether deep water corals build reefs: do shallow reef models apply? 6-18. En: Willison, J. H., J. Hall, S. E. Gass, E. L. R. Kenchington, M. Butler y P. Doherty (Eds.). Proc. First Int. Symp. Deep-Sea Corals. Ecology Action Centre & Nova Scotia Museum, Halifax, Nova Scotia. 231 p. [ Links ]
30 INVEMAR, UAESPNN, CARDIQUE, EPA y Cartagena. 2003. Modelo de desarrollo sostenible para los Archipiélagos del Rosario y San Bernardo. Informe técnico, resolución 456-03, Santa Marta. 256 p. [ Links ]
31 Kelleher, G. y R. Kenchington. 1991. Guidelines for establishing marine protected areas. A marine conservation and development report. IUCN, Gland, Suiza. 90 p. [ Links ]
32 Kelleher, G., C. Bleakley y S. Wells. 1995. A global representative system of marine protected areas. Vol. II. Wider Carribbean, West Africa and South Atlantic. The Great Barrier Reef Marine Park Authority, The World Bank and The World Conservation Union (IUCN), Washington, D.C. 93 p. [ Links ]
33 King, M. C. y K. F. Beazley. 2005. Selecting focal species for marine protected area network planning in the Scotia-Fundy region of Atlantic Canada. Aq. Cons., 15: 367-385. [ Links ]
34 Krieger, K. G. 2001. Coral (Primnoa) impacted by fishing gear in the Gulf of Alaska. 106-116. En: Willison, J. H., J. Hall, S. E. Gass, E. L. R. Kenchington, M. Butler y P. Doherty (Eds.). Proc. First Int. Symp. Deep-Sea Corals. Ecology Action Centre and Nova Scotia Museum, Halifax, Nueva Escocia. 231 p. [ Links ]
35 Lattig, P. 2000. Porifera, Cnidaria y Annelida de la franja superior del talud continental (200-500 m) del Caribe colombiano. Tesis Biol., Pontificia Univ. Javeriana, Santa Marta. 382 p. [ Links ]
36 Lattig, P. y J. Reyes. 2001. Nueve primeros registros de corales azooxanthellados (Anthozoa: Scleractinia) del Caribe colombiano (200-500 m). Bol. Inv. Mar. Cost., 30: 19-38. [ Links ]
37 Levin, L. A., R. J. Etter, M. A. Rex, A. J. Gooday, C. R. Smith, J. Pineda, C. T. Stuart, R. R. Hessler y D. Pawson. 2001. Environmental influences on regional deep-sea species diversity. Ann. Rev. Ecol. Syst., 32: 51-93. [ Links ]
38 Lourie, S. A. y A. C. Vincent. 2004. Using biogeography to help set priorities in marine conservation. Conserv. Biol., 18 (4): 1004-1020. [ Links ]
39 Manjarrés, L., H. Zúñiga, L. O. Duarte, C. B. García, E. Causado, J. Altamar, J. Viaña, F. Escobar, F. Cuello, D. Pérez, D. Ríos, K. Tejada y J. Sánchez. 2006. Evaluación de innovaciones en la tecnología de captura de la pesquería industrial de arrastre camaronero del Caribe colombiano con fines ecológicos y de productividad. Informe técnico, Programa de investigación e innovación tecnológica de la flota industrial camaronera del mar Caribe de Colombia, GIEEP, LIPET, INTROPIC, Univ. Magdalena, INVEMAR (Coord. Nal. Proy. FAO), Dista LTDA., Pesquera Continental S.A. y ASOPESCARIBE, Santa Marta. 57 p. [ Links ]
40 Margules, C. R. y R. L. Pressey. 2000. Systematic conservation planning. Nature, 405: 243-253. [ Links ]
41 Medina, M. 2002. Ensamblaje de peces demersales explotados por la flota industrial camaronera en la plataforma continental de La Guajira (Caribe colombiano). Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Santa Marta. 85 p. [ Links ]
42 Ministerio de Ambiente, Vivienda y Desarrollo Territorial de Colombia (MAVDT). 2005. Resolución 679 de 2005 (mayo 31) del Ministerio de Ambiente, Vivienda y Desarrollo Territorial de Colombia Diario Oficial No. 45.927 de 02 de junio de 2005. [ Links ]
43 Mora, J. 1989. Observaciones sobre la pesca industrial del camarón en la península de La Guajira. Informe técnico, INDERENA, Cartagena. [ Links ]
44 Morgan, L. E., P. Etnoyer, A. J. Scholz, M. Mertens y M. Powell. 2005. Conservation and management implications of deep-sea coral and fishing effort distributions in the Northeast Pacific Ocean. 1171-1187. En: Freiwald, A. y J. M. Roberts (Eds.). Cold-water corals and ecosystems. Springer, Berlin, Heidelberg. 1243 p. [ Links ]
45 Osorio, D. 1986. Estimaciones de las capturas de camarón y fauna acompañante, en el Caribe colombiano durante el 6-12 de junio de 1986. Informe técnico, INDERENA, Cartagena. 41 p. [ Links ]
46 Patiño, F. y F. Flórez. 1993. Estudio ecológico del golfo de Morrosquillo. Univ. Nacional de Colombia y Fondo FEN-COLOMBIA, Bogotá. 295 p. [ Links ]
47 Porter, J. W. 1972. Patterns of species diversity in Caribbean reef corals. Ecology, 53 (4): 745-748. [ Links ]
48 Prahl, H. von y H. Erhardt. 1989. Lista anotada de corales ahermatípicos de Colombia. 539-556. En: CCO (Ed.). Memorias del VI Seminario Nacional de Ciencias y Tecnologías del Mar. Comisión Colombiana de Oceanografía, Bogotá. [ Links ]
49 Reed, J., A. N. Shephard, C. C. Koenig, K. M. Scanlon y R. G. Gilmore. 2005. Mapping, habitat characterization, and fish surveys of the deep-water Oculina coral reef marine protected area: a review of historical and current research. 443-465. En: Freiwald, A. y J. M. Roberts (Eds.). Coldwater corals and ecosystems. Springer, Berlín, Heidelberg. 1243 p. [ Links ]
50 Reyes, J. y N. Santodomingo. 2002. Manual de identificación CITES de invertebrados marinos de Colombia. Serie manuales de identificación CITES de Colombia, No. 8. INVEMAR, Santa Marta. 97 p. [ Links ]
51 Reyes, J., N. Santodomingo, A. Gracia, G. Borrero-Pérez, G. Navas, L. M. Mejía-Ladino, A. Bermúdez y M. Benavides. 2005. Southern Caribbean azooxanthellate coral communities off Colombia. 309-330. En: Freiwald, A. y J. M. Roberts (Eds.). Cold-water corals and ecosystems. Springer, Berlín, Heidelberg. 1243 p. [ Links ]
52 Roa, A. 2000. Caracterización de la comunidad íctica demersal de la región sur del Caribe colombiano (300 y 500 m) y algunas consideraciones zoogeográficas. Tesis Biol., Univ. Nacional de Colombia, Santa Marta. 415 p. [ Links ]
53 Roa-Varón, A., L. Saavedra-Díaz, A. Acero, L. S. Mejía y G. R. Navas. 2003. Nuevos registros de peces óseos para el Caribe colombiano de los órdenes Beryciformes, Zeiformes, Perciformes y Tetraodontiformes. Bol. Inst. Inv. Mar. Cost., 32: 3-24. [ Links ]
54 Roberts, C. M. y J. P. Hawkins. 2000. Fully-protected marine reserves: a guide. WWF Endangered Seas Campaign, Washington y Environment Department, University of York, York, Reino Unido. 131 p. [ Links ]
55 Roberts, C. M., D. J. McClean, J. E. N. Veron, J. P. Hawkins, G. R. Allen, D. E. Mcallister, C. G. Mittermeier, F. W. Schueler, M. Spalding, F. Wells, C. Vynne y T. B. Warner. 2002. Marine biodiversity hotspots and conservation priorities for tropical reefs. Science, 295: 1280-1284. [ Links ]
56 Roberts, C. M., S. Andelman, G. Branch, R. H. Bustamante, J. C. Castilla, J. Dugan, B. S. Halpern, K. D. Lafferty, H. Leslie, J. Lubchenco, D. Mcardle, H. P. Possingham, M. Ruckelshaus y R. R. Warner. 2003a. Ecological criteria for evaluating candidate sites for marine reserves. Ecol. Appl., 13 (1): 199-214. [ Links ]
57 Roberts, C. M., S. Andelman, G. Branch, R. H. Bustamante, J. C. Castilla, J. Dugan, B. S. Halpern, K. D. Lafferty, H. Leslie, J. Lubchenco, D. Mcardle, H. P. Possingham, M. Ruckelshaus y R. R. Warner. 2003b. Application of ecological criteria in selecting marine reserves and developing reserve networks. Ecol. Appl., 13 (1): 215-228. [ Links ]
58 Roberts, J. M., A. J. Wheeler y A. Freiwald. 2006. Reefs of the deep: the biology and geology of coldwater coral ecosystems. Science, 312: 543-547. [ Links ]
59 Roberts, J. M., A. J. Wheeler, A. Freiwald y S. Cairns. 2009. Cold-water corals: the biology and geology of deep-sea coral habitats. Primera edición, Cambridge University Press, Cambridge. 334 p. [ Links ]
60 Roberts, S. y M. Hirshfield. 2004. Deep-sea corals: out of sight, but no longer out of mind. Front. Ecol. Environ., 2 (3): 123-130. [ Links ]
61 Saavedra-Díaz, L., A. Acero y G. Navas. 2000. Lenguados de la familia Paralichthyidae (Pisces: Pleuronectiformes) conocidos del Caribe colombiano. Rev. Acad. Colomb. Cienc. Exact. Fís. Nat., 24: 295-310. [ Links ]
62 Salm, R. V. y J. Clark. 1984. Marine and coastal protected areas: a guide for planners and managers. IUCN, Gland, Suiza. 396 p. [ Links ]
63 Salm, R.V., J. Clark y E. Sirila. 2000. Marine and coastal protected areas: a guide for planners and managers. UICN, Washington D. C. 371 p. [ Links ]
64 Santodomingo, N. y J. Reyes. 2003. Are there southern Caribbean deep-sea coral banks? Three possible scenarios. Abstract 2nd International Symposium on Deep-Sea Corals. Erlanger Geol Abh Sonder, Erlangen. 73 p. [ Links ]
65 Santodomingo, N., J. Reyes, A. Gracia, A. Martínez, G. Ojeda y C. García. 2007. Azooxanthellate Madracis coral communities off San Bernardo and Rosario Islands (Colombian Caribbean). Bull. Mar. Sci., 81 (1): 273-287. [ Links ]
66 Sauer, W. H. H., M. J. Smale y M. R. Lipinski. 1992. The location of spawning grounds, spawning and schooling behaviour of the squid Loligo vulgaris reynaudii (Cephalopoda: Myopsida) off the eastern Cape Coast, South Africa. Mar. Biol., 114: 97-107. [ Links ]
67 Shester, G. y J. Ayers. 2005. A cost effective approach to protecting deep-sea coral and sponge ecosystems with an application to Alaska's Aleutian Islands region. 1151-1169. En: Freiwald, A. y J. M. Roberts (Eds.). Cold-water corals and ecosystems. Springer, Berlín, Heidelberg. 1243 p. [ Links ]
68 Tavares, M. 2002. Shrimps. 251-262. En: Carpenter, K. E. (Ed.). FAO Species identification guide for fishery purposes and American Society of Ichthyologists and Herpetologists special publication No. 5. Vol. 1, FAO, Roma. 600 p. [ Links ]
69 Tunesi, L., G. Diviacco y G. Mo. 2001. Observations by submersible on the biocoenosis of the deep-sea corals off Portofino promontory (Northwestern Mediterranean Sea). 76-87. En: Willison, J. H. M., J. Hall, S. F. Gass, E. L. R. Kenchington, M. Butler y P. Doherty (Eds.). Proc. First Int. Symp. Deep-Sea Corals. Halifax, Nueva Escocia. 231 p. [ Links ]
70 UNEP (United Nations Environment Programme). 2006. WWF's North-East Atlantic programme. http://www.ngo.grida.no/wwfneap/Publication/briefings. 27/02/2006. [ Links ]
71 Vernette, G. 1985. La plateforme continentale Caräibe de Colombie (du debouché du Magdalena au golfe du Morrosquillo). Importance du diapirisme argileux sur la morphologie et la sedimentation. Tesis en Ciencias, Univ. de Bordeaux, Bordeaux, Francia. 318 p. [ Links ]
72 Vernette, G., A. Mauffret, C. Bobier, L. Briceño y J. Gayet. 1992. Mud diapirism, fan sedimentation and strike-slip faulting, Caribbean Colombian margin. Tectonophysics, 202 (2-4): 335-349. [ Links ]
73 Viaña, J. E., J. A. Medina, M. Barros, L. M. Manjarrés, J. Altamar y M. Solano. 2004. Evaluación de la ictiofauna demersal extraída por la pesquería industrial de arrastre en el área norte del Caribe colombiano (Enero/2000-Junio/2001). 115-151. En: Manjarrés, L. (Ed.). Pesquerías demersales del área norte del mar Caribe de Colombia y parámetros biológico-pesqueros y poblacionales del recurso pargo. Univ. de Magdalena, Santa Marta. 250 p. [ Links ]
74 Zúñiga, H., J. Altamar y L. Manjarrés. 2004. Caracterización tecnológica de la flota de arrastre camaronero del mar Caribe de Colombia. Proyecto EP/GLO/201/GEF. Reducción by-catch. FAO. Univ. Magdalena, Santa Marta. 20 p. [ Links ]
FECHA DE RECEPCIÓN: 22/07/2007 FECHA DE ACEPTACIÓN: 25/01/2011
*Contribución No. 1075 del Instituto de Investigaciones Marinas y Costeras-INVEMAR.
