










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.40 no.2 Santa Marta July/Dec. 2011
COBERTURA Y ESTRUCTURA DE LOS BOSQUES DE MANGLE EN LA BAJA GUAJIRA, CARIBE COLOMBIANO*
EXTENSION AND STRUCTURE OF MANGROVE FORESTS IN THE BAJA GUAJIRA, COLOMBIAN CARIBBEAN
Alianis M. Orjuela-Rojas1, Carlos A. Villamil1 y Adolfo Sanjuan-Muñoz2
1 Instituto de Investigaciones Marinas y Costeras-INVEMAR, Cerro Punta Betín, Santa Marta, Colombia, alianis_orjuela@invemar.org.co (A.O.R.), carlos.villamil@invemar.org.co (C.A.V.)
2 Universidad Jorge Tadeo Lozano, Carrera 2 No. 11-68, El Rodadero, Santa Marta, Colombia, adolfo.sanjuan@utadeo.edu.co
RESUMEN
En abril de 2007 se estimó la cobertura y se determinaron los atributos estructurales de los bosques de mangle establecidos en la Baja Guajira. La cobertura del bosque se estimó a partir de imágenes ASTER y de un mosaico de fotografías aéreas. La estructura se determinó sobre transectos de 10 m de ancho y del largo de la franja del bosque de mangle. En cada uno de éstos se identificaron y contaron los árboles de mangle, midiéndoles altura total y diámetro a la altura del pecho (DAP) y se calculó el área basal, la densidad y el índice de Valor de Importancia (IVI). En la Baja Guajira se estimó una cobertura de mangle de 59.9 ha. Laguncularia racemosa fue la especie más frecuente (87%) y dominante (0.54 m2 ha-1), Rhizophora mangle se presentó en el 75% de las estaciones, mientras Avicennia germinans se encontró únicamente al norte del área de estudio. Las mayores alturas y DAP promedios de R. mangle, L. racemosa y A. germinans se presentaron en Rincón Mosquito (12.9±2.3 m y 22.4±11.2 cm), Puerto Brisa (12.2±5.3 m y 20.8±9.8 cm) y La Enea (13.8±4.3 m y 23.8±14.8 cm) respectivamente, sectores caracterizados por tener aportes constantes de agua dulce. La estación de Dibulla constituida por una floresta monoespecífica de Conocarpus erectus presentó el bosque menos desarrollado con una altura promedio de 2.8±0.6 m y DAP promedio de 3.0±0.9 cm. En cuanto a la regeneración natural, L. racemosa presentó la mayor proporción de plántulas en el área de estudio, R. mangle se encontró en las estaciones parcialmente inundadas y A. germinans presentó la mayor densidad de plántulas y propágulos en la estación de La Enea.
PALABRAS CLAVE: Manglares, Estructura, Cobertura, Regeneración Natural, Baja Guajira.
ABSTRACT
In April 2007, cover and structural attributes of mangrove forests in the Baja Guajira were estimated and identified. Forest cover was estimated from ASTER images and aerial photographs. The forest structure was evaluated along transects which were 10 m broad and as long as the width of the mangrove fringe. In each transect mangrove trees were identified and counted. The total height and the diameter at breast height (DBH) of the trees were measured and subsequently the basal area, density and Importance Value Index (IVI) calculated. The estimated mangrove cover of the Baja Guajira was 59.9 ha. The most frequent and dominant species was Laguncularia racemosa (0.54 m2 ha-1). Rhizophora mangle was present in 75% of the stations, while Avicennia germinans was only found in the north of the study area. The highest average heights and diameters for R. mangle, L. racemosa and A. germinans were determined in Rincón Mosquito (12.9±2.3 m and 22.4±11.2 cm), Puerto Brisa (12.2±5.3 m and 20.8±9.8 cm) and La Enea (13.8±4.3 m and 23.8±14.8 cm), sectors with a constant fresh water supply. The less developed forest, a monospecific forest of Conocarpus erectus, was observed in Dibulla station, with height and diameter averages of 2.8±0.6 m and 3.0±0.9 cm. In terms of natural regeneration, L. racemosa presented the highest proportion of seedlings in the study area, R. mangle was found in stations partly flooded and A. germinans had the highest density of seedlings and propagules at La Enea station.
KEY WORDS: Mangrove, Structure, Cover, Natural Regeneration, Baja Guajira.
INTRODUCCIÓN
Los bosques de mangle son formaciones de plantas halófitas facultativas, establecidas en la franja intermareal, que bordean bahías, lagunas costeras, estuarios, deltas y desembocaduras de ríos (Tomlinson, 1986). A pesar de los beneficios ecológicos, económicos y sociales que brinda este ecosistema, es objeto de fuertes impactos debidos principalmente al desarrollo urbano, la tala, el relleno, la contaminación y la explotación forestal sin ordenamiento (Alongi, 2002; Gil-Torres et al., 2009).
Frente a esta problemática, el Ministerio de Ambiente, Vivienda y Desarrollo Territorial (MAVDT) ha generado una serie de normas con la intención de garantizar la conservación de estos ecosistemas en el marco del concepto del Desarrollo Sostenible, permitiendo así equilibrar el progreso de las actividades socioeconómicas sin comprometer el potencial y la protección de los manglares. En este contexto el Ministerio del Medio Ambiente (ahora MAVDT) emitió las resoluciones 1602 de 1995 y 020 de 1996, las cuales reglamentan la implementación obligatoria de estudios de caracterización, diagnóstico, evaluación y zonificación de las áreas de manglar del país (Sánchez-Paéz et al., 1997).
Por tal motivo, en los últimos años en el departamento de La Guajira se han realizado estudios relacionados con aspectos ambientales y ecológicos de los bosques de mangle (Ulloa-Delgado et al., 1998; Sánchez-Páez et al., 2000; Arroyo, 2001; Martínez y Ruíz, 2001; Rosado et al., 2003; Gómez, 2005; Gil-Torres et al., 2009), brindando información de gran importancia para la implementación de medidas preventivas, correctivas o de mitigación a impactos causados a estos ecosistemas. Sin embargo, son insuficientes y poco detalladas las investigaciones efectuadas en la Baja Guajira, a pesar de que este sector exhibe las mejores condiciones ambientales para el desarrollo de este tipo de vegetación, dada la influencia fluvial proveniente de la Sierra Nevada de Santa Marta. El presente estudio tiene como objetivo proporcionar información detallada y actualizada de la cobertura y estado actual de los bosques de mangle de la Baja Guajira.
ÁREA DE ESTUDIO
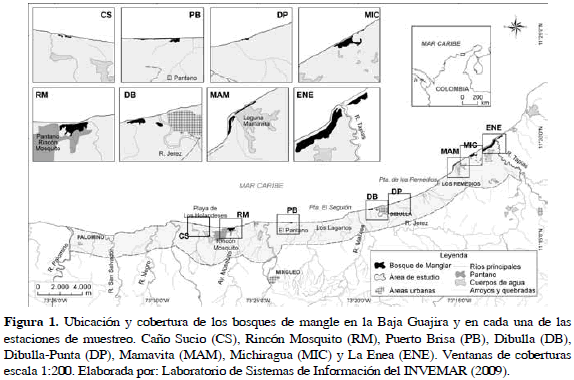
La Baja Guajira está ubicada al sur del departamento de La Guajira, comprende la franja costera desde el río Tapias (este) hasta el río Palomino (oeste). La conforman las estribaciones de la Sierra Nevada de Santa Marta (SNSM), los valles de los ríos Ranchería y Cesar y la Serranía de Perijá, que en conjunto generan cuantiosos drenajes de ríos y quebradas que alimentan los cuerpos lagunares (Ricerca e Cooperazione et al, 2002) y hacen de esta región una zona con diferentes tipos de humedales como las lagunas estuarinas de Trupía y Mamavita, los sistemas riberino-estuarino como Rincón Mosquito, Caño Sucio, Limoncito y El Lagarto; los sistemas de tipo rivereño al que pertenecen los ríos principales de la zona como Palomino, Negro, San Salvador, Ancho, Cañas, Maluisa, Mariamina, Jerez y Tapias y los sistemas palustres con bosques bien desarrollados como los que se encuentran en el río Cañas, en el gran pantano de Dibulla y entre el pantano Michiragua y Laguna Grande (Ricerca e Cooperazione et al., 2002).
Esta región se encuentra en una transición entre las zonas de vida bosque muy seco tropical (bms-T) y bosque seco tropical (bs-T), según la clasificación de Holdridge (Espinal y Montenegro, 1963). De acuerdo con los valores obtenidos entre los años 1994 y 2007 de la estación meteorológica Termoguajira (IDEAM), ubicada en el municipio de Dibulla, Gil-Torres et al. (2009) registran para la Baja Guajira una precipitación media anual de 1401 mm con distribución bimodal y un máximo entre septiembre y octubre, una temperatura media anual de 27 °C, con el período más cálido entre junio y agosto, la humedad relativa media anual entre 70 y 80% y la evaporación media de 1712 mm a-1. Como en el resto del Caribe colombiano, las mareas en La Guajira son casi imperceptibles, pues entre pleamar y bajamar la diferencia en el nivel vertical de las aguas no supera los 35 cm. Sin embargo ciertas perturbaciones atmosféricas ocasionales, como las que produce el paso de tormentas tropicales y huracanes, a una distancia considerable de la costa, además de lluvias torrenciales, pueden ocasionar "mares de leva" que eventualmente anegan los playones y extensiones de tierra en las costas de escaso relieve (Guerra et al., 2003). Es importante resaltar que la mayor parte de ecosistemas de manglar de la Baja Guajira, se establecen detrás de una barra de arena, donde los árboles no están expuestos a las mareas, sino a cambios del nivel de agua y salinidad de acuerdo a la temporalidad.
MATERIALES Y MÉTODOS
Recolecta de datos en campo
A partir del procesamiento e interpretación de un mosaico de imágenes ASTER de La Guajira (año 2002), del mapa de cobertura de la tierra del año 2006 (escala 1:100000) generado en el Laboratorio de Sistemas de Información (LabSI) del INVEMAR y de visitas previas de reconocimiento, se escogieron los sitios de muestreo: Caño Sucio (CS), Rincón Mosquito (RM), Puerto Brisa (PB), Dibulla (DB), Dibulla- Punta (DP), Mamavita (MAM), Michiragua (MIC) y La Enea (ENE) (Tabla 1; Figura 1). En abril de 2007 (época seca), se realizó la medición de los atributos estructurales de los bosques de mangle siguiendo la metodología descrita por Cintrón-Molero y Schaeffer-Novelli (1984). En cada estación se delimitó un cinturón perpendicular al cuerpo de agua principal de 10 m de ancho y 100 m de longitud o de la totalidad de la franja de manglar. El cinturón fue dividido en cuadrantes de 100 m2 (10 x 10 m) donde se identificaron y contaron todos los árboles de mangle, los que se les midió la altura total (m) y el diámetro a la altura del pecho (DAP, cm), con un cimómetro electrónico Häglof y una cinta diamétrica respectivamente.
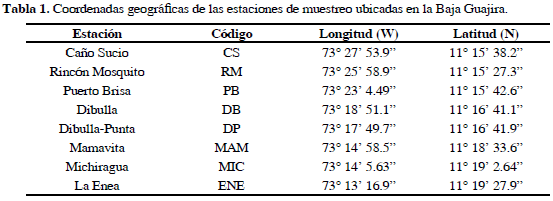
Se realizó un perfil de vegetación de la totalidad de la franja o hasta 100 m de longitud sobre el transecto principal, consignando en una matriz de campo la descripción de cada uno de los individuos (especie, altura y DAP). Adicionalmente, para determinar el reclutamiento del bosque de manglar se delimitó un cinturón de 1 m de ancho por 100 m de largo, paralelo al transecto de estructura, donde fueron identificados y contados todos los propágulos y plántulas presentes. Todos los árboles medidos fueron clasificados en tres categorías diamétricas siguiendo a Cintrón-Molero y Schaeffer-Novelli (1992): brinzales (2.5 ≤ x < 5 cm), latizales (5 ≤ x < 10 cm) y fustales (≥ 10 cm). Por último se realizaron mediciones in situ de salinidad en la fuente de agua más cercana empleando un conductímetro (YSI EC300, Yellow Springs, OH, EE.UU.) y nivel de agua superficial cada 10 m sobre el transecto de estructura.
Determinación de la cobertura de los bosques de mangle
Se interpretó únicamente la cobertura Bosque de Mangle, que comprende florestas conformados exclusivamente por una o más especies de mangle (R. mangle, A. germinans, L. racemosa y C. erectus). Coberturas de bosques de manglar que incluían otro tipo de vegetación arbórea asociada no se tuvieron en cuenta en la interpretación. Para la delimitación de la cobertura Bosque de Mangle de la Baja Guajira, se utilizaron imágenes multiespectrales ASTER (2002) y fotografías aéreas (vuelos C-287-2 de 2005, esc. 1:5000 y C-2687-3 de 2005, esc. 1:5000). El procesamiento de las imágenes se realizó siguiendo la propuesta metodológica de Lozano y Sierra-Correa (2005), la cual es utilizada por el LabSI del INVEMAR en la interpretación de bosques de manglar en diferentes áreas de las zonas costeras del país.
Análisis de datos
Para cada estación se calculó la abundancia, el área basal (m2 ha-1), la densidad (ind 0.1 ha-1), la frecuencia, el índice de Valor de Importancia (IVI) de cada una de las especies, de acuerdo con las fórmulas establecidas por Schaeffer-Novelli y Cintrón-Molero (1986) y diámetro de área basal promedio (Dg), con base en la ecuación propuesta por Lema (2003). A todas las variables obtenidas se le realizó un análisis exploratorio determinando la media aritmética y la desviación estándar (x ± DE). Con el fin de comparar y definir asociaciones biológicas espaciales se efectuó un análisis de clasificación usando el Dg, aplicando el índice de similaridad de Bray-Curtis y empleando como estrategia de agrupamiento el ligamiento promedio no ponderado (Clarke y Warwick, 2001). Los datos fueron transformados con logaritmo (x+1) para que todas las especies contribuyeran más equitativamente a la similaridad (McCune y Grace, 2002).
RESULTADOS
Características estructurales generales
Fueron medidos un total de 646 árboles de mangle, pertenecientes a cuatro especies: Laguncularia racemosa (L.) C. F. Gaertn con 282 individuos (43.6%), Conocarpus erectus L. con 176 individuos (27.2%), Rhizophora mangle L., con 114 individuos (17.6%) y Avicennia germinans (L.) Stearn, con 74 individuos (11.4%) (Tabla 2). El área de estudio estuvo dominada por L. racemosa, que además de ser la especie más abundante fue la más frecuente, encontrándose en siete de las ocho estaciones de muestreo. Presentó la mayor densidad y área basal del sector, y por ende el mayor IVI con 121.5 (Tabla 2).
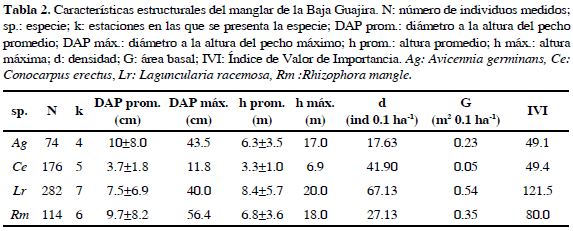
Rhizophora mangle se presentó en el 75% de estaciones (CS, RM, PB, MAM, MIC y ENE) asociada a L. racemosa en cada una de ellas (Figura 2). Aunque R. mangle tiene mayor desarrollo estructural (DAP) que L. racemosa, la densidad y área basal fueron menores, catalogándola como la segunda especie en importancia del sector (Tabla 2).
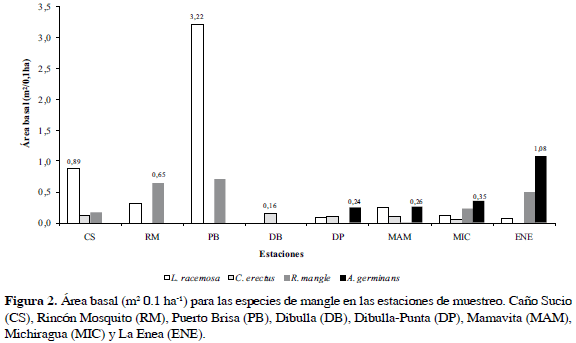
La especie A. germinans se encontró únicamente en los transectos situados desde el río Jerez hasta el río Tapias (DP, MAM, MIC y ENE), a pesar de la baja densidad obtuvo una área basal de 0.23 m2 0.1 ha-1, dominando en la comunidad vegetal de La Enea, por la presencia de individuos bien desarrollados con DAP de hasta 43.5 cm (Tabla 2; Figura 2). Finalmente, C. erectus, aunque con una de las densidades más altas del sector, registró el menor aporte al área basal (4.7%), con individuos que presentaron DAP de 3.7 ±1.8 cm (Tabla 2).
En cuanto a las categorías diamétricas, los brinzales tuvieron una densidad de 75.2 ind 0.1 ha-1 (48.9%), los latizales 52.6 ind 0.1 ha-1 (34.2%) y los fustales 25.9 ind 0.1 ha-1 (16.8%), está última categoría aunque con menor abundancia de árboles aportó el 78.6% del área basal (Figura 3). Por último y de acuerdo con el procesamiento de las imágenes satelitales y las fotografías aéreas, los bosques de mangle muestreados ocuparon una área total de 59.9 ha, registrándose el 69.6% de la cobertura en las estaciones al norte del área de estudio (ENE, MIC, MAM y DP), mientras que los bosques de mangle ubicados en la parte sur de la Baja Guajira (CS, RM, PB y DB) sólo cubren un área de 18.75 ha que corresponde al 31.3%.
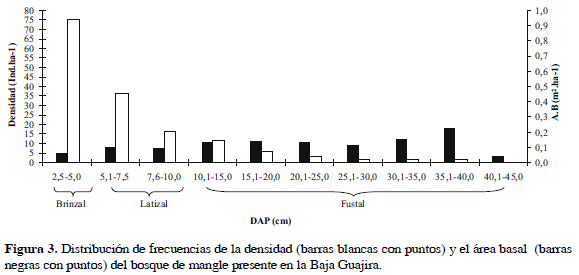
Caracterización de las estaciones de muestreo
La estación de Caño Sucio con 0.56 ha en cobertura de Bosque de Mangle, presentó una franja de mangle mixta y discontinua de aproximadamente 20 m de ancho. En la parte inicial se observaron individuos de L. racemosa hasta de 13.7 cm de DAP y 6.3 m de altura, establecidos sobre un sustrato arenoso. Posteriormente, se presentaron árboles de R. mangle de 7.1±2.5 cm de DAP y 3.9±1.6 m de altura, sobre sustratos limosos y arcillosos, influenciados por el agua dulce proveniente del caño Sucio (salinidad = 0.1), mientras que al interior del rodal, se observaron individuos de C. erectus con 8.0±2.9 cm de DAP y 6.1±0.5 m de altura, establecidos sobre un sustrato consolidado, acompañado por especies de bosque seco tropical (Figura 4a). En términos generales el bosque de Caño Sucio estuvo dominado por L. racemosa, la especie más abundante, con la mayor área basal y de mayor importancia biológica.
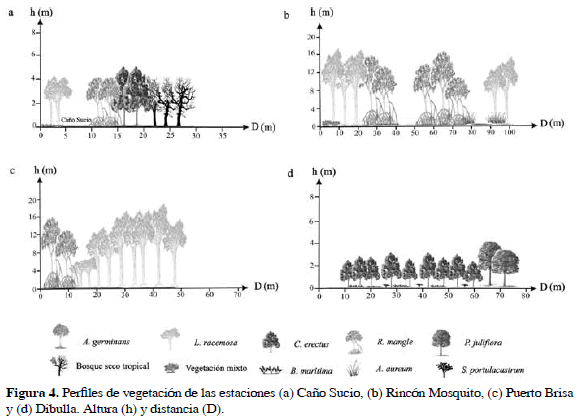
La estación de Rincón Mosquito con una cobertura de Bosque de Mangle de 10.9 ha, presentó una franja de mangle mayor a 100 m de ancho, colonizada inicialmente por individuos de L. racemosa de hasta 11 cm de DAP y 17.8 m de altura, asociados con vegetación herbácea, entre ellas Batis maritima L. (Familia Bataceae). A partir de los 20 m se observó una franja constituida exclusivamente por R. mangle, cuyos individuos más desarrollados (alturas entre 8 y 15 m y DAP hasta de 56.4 cm), se presentaron al interior del bosque entre los 50 a 80 m. Por último, en la parte final del transecto (90 m) se presentaron fustales de L. racemosa con DAP de 13.2±4.0 cm y alturas de 12.9±2.4 m, asociados con Acrostichum aureum L. (Figura 4b). Aunque no fue evidente el predominio de una especie de mangle en particular, L. racemosa por tener mayor densidad que R. mangle presentò mayor valor de importancia (Tabla 3).
La estación de Puerto Brisa con una cobertura de 3.12 ha, presentó un bosque de mangle bordeando un pequeño cuerpo de agua dulce (salinidad = 0.1). La franja inicial del rodal la constituyeron en su mayoría fustales de R. mangle de hasta 36.8 cm de DAP y 18 m de altura, con un sistema radicular desarrollado, con raíces fúlcreas hasta de 5 m de altura. Posterior a ésta se observaron individuos de L. racemosa con DAP de hasta 40 cm y alturas entre 11 y 20 m (Figura 4c). Laguncularia racemosa fue la especie dominante en esta estación, aportando más del 80% del área basai total, mientras que R. mangle por su baja densidad tuvo un IVI de 50, a pesar que sus individuos presentaron buen porte (Tabla 3).
La estación de Dibulla ubicada al costado suroccidental del río Jerez exhibió una cobertura de Bosque de Mangle de 4.19 ha, y una salinidad de 0.1, presentó un bosque monoespecífico de aproximadamente 60 m de ancho, constituido por árboles achaparrados y muy ramificados de C. erectus con alturas y DAP promedio de 2.8±0.6 m y 3.0±0.9 cm respectivamente (Tabla 3). Hacia el interior del bosque se observaron claros cubiertos por especies halófitas como Sesuvium portulacastrum L. (Familia Aizoaceae) y B. maritima. A partir de los 60 m, C. erectus formó pequeñas agrupaciones integradas con especies de bosque seco tropical y matorral espinoso (Figura 4d). A pesar de la gran densidad de árboles de C. erectus, el área basai fue baja dado que la mayoría de individuos fueron poco desarrollados.
La estación de Dibulla-Punta con 0.36 ha en cobertura de Bosque de Mangle, presentó una franja de 40 m de ancho aproximadamente. En los primeros 20 m se presentaron fustales de A. germinans, latizales de L. racemosa y parches de B. maritima. En los 20 m restantes A. germinans fue remplazada por latizales de L. racemosa y C. erectus, esta última se asoció con vegetación xerofítica y bosque seco tropical al final del transecto (Figura 5a). El mar fue el cuerpo de agua de principal influencia sobre esta franja de manglar, sin embargo al costado izquierdo se presentó un pequeño cuerpo de agua con salinidades de hasta 79.2.
Avicennia germinans fue la especie de mayor importancia con IVI de 107.1, dado que presentó los mejores desarrollos estructurales con árboles de 15.2±4.8 cm de DAP y 6.7±2.6 m de altura. Por otra parte, L. racemosa presentó la mayor densidad, razón por la cual obtuvo un IVI de 102.8, catalogándose como la segunda especie en importancia del sector (Tabla 3).
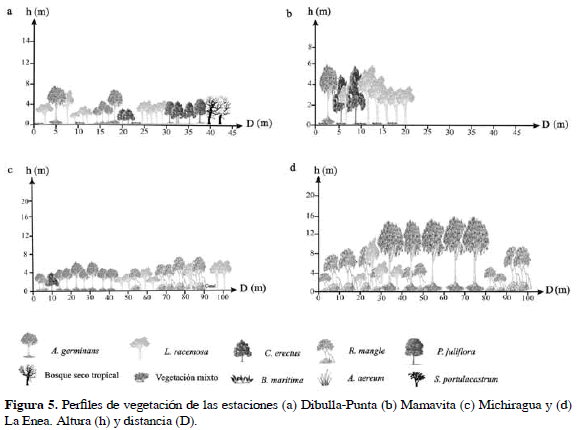
La estación de Mamavita presentó un Bosque de Mangle de 3.92 ha y de 20 m de ancho aproximadamente. Constituido inicialmente por una franja mixta de fustales de A. germinans, L. racemosa y algunos árboles de C. erectus, que se encontraron colonizando sustratos arenosos con salinidad de 35.7. De 10 a 20 m se observó una franja de L. racemosa con 4.0±1.4 cm de diámetro y 3.3±0.9 m de altura. A partir de los 20 m no se observó vegetación de mangle, debido al incremento de la salinidad (93.26±12.43), lo cual favoreció la presencia de B. maritima y S. portulacastrum, especies que toleran ambientes salinos (Figura 5b).
La especie predominante, por su densidad, distribución y dominancia fue L. racemosa, con un IVI de 154. Por otro lado A. germinans, aportó el 41.6% del área total del bosque lo cual la catalogó como la segunda especie en importancia del sector (Tabla 3).
La estación Michiragua, con 7.1 ha en cobertura de Bosque de Mangle, presentó un rodal que se extendió más allá de los 100 m. Compuesto inicialmente por A. germinans, R. mangle y C. erectus, seguido por una franja monoespecifica de A. germinans, con alturas entre 5.5 y 11.8 m. A partir de los 40 m, se registraron juveniles de L. racemosa con alturas entre 2.5 y 8 m mezclados con individuos de R. mangle y árboles muertos de A. germinans. De 60 a 90 m el bosque estuvo conformado exclusivamente por individuos de R. mangle que limitaban con un canal de 5 cm de profundidad y 39 de salinidad, después del cual se observó un parche compuesto por L. racemosa (Figura 5c).
Avicennia germinans, la especie más representativa, registró un IVI de 106.7, siendo el área basal y la densidad las variables que más contribuyeron en el índice. Rhizophora mangle registró un IVI de 100.2, no solo por su frecuencia (78%), sino también por la densidad y el área basal (Tabla 3).
Por último, la estación La Enea con la mayor cobertura de Bosque de Mangle en el área de estudio (29.75 ha), presentó un rodal que superó los 100 m de ancho. La franja inicial del manglar estuvo constituida por individuos de R. mangle que incrementaban la altura hacia el interior del bosque, estos árboles colonizaron un sustrato lodoso con un nivel de agua superficial de 20 cm.
A partir de los 20 m se observaron fustales de R. mangle acompañados de juveniles de L. racemosa, dichas especies fueron remplazadas desde los 40 m por individuos bien desarrollados de A. germinans con alturas entre 10 y 16 m. A partir de los 80 m el nivel de agua superficial aumentó a 40 cm, y se presentó un bosque constituido por juveniles de R. mangle que fueron beneficiados por las salinidades bajas (1.5±0.25) a causa de la inundación. Finalmente entre 90 y 100 m, el nivel del agua superficial llegó a 60 cm, los árboles de R. mangle alcanzaron mayores tallas (entre 7.8 y 8.4 m) formando parches donde las raíces se entremezclaban (Figura 5d). Rhizophora mangle fue la especie con mayor IVI (152.3), lo cual está relacionado con la densidad de individuos y la distribución relativamente homogénea en la estación, mientras que A. germinans, la segunda especie en importancia (IVI = 103.4), presentó la mayor área basal debido al desarrollo de sus individuos que presentaron los mayores registros de DAP y alturas promedios (Tabla 3).
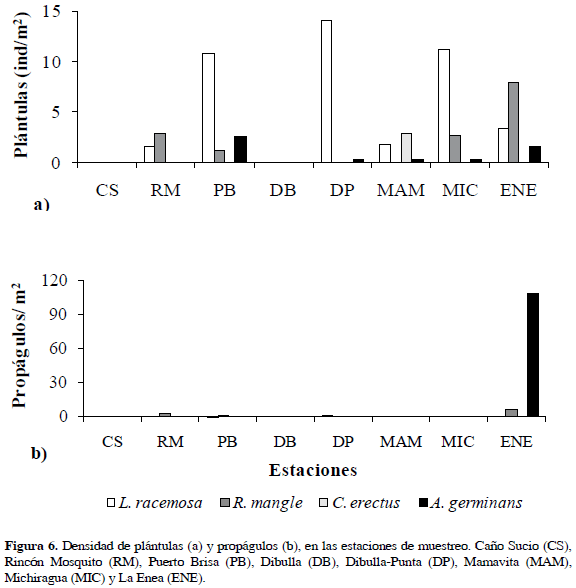
Reclutamiento del bosque
La especie con mayor densidad de plántulas fue L. racemosa, presentándose en la estación de Dibulla-Punta el valor más alto (Figura 6a). Rhizophora mangle, por su parte, sólo presentó establecimiento de plántulas en las estaciones parcialmente inundadas (RM, PB, MIC y ENE). Aunque en términos generales la densidad de propágulos fue baja o nula (Figura 6b), en la estación de La Enea se observó gran cantidad de propágulos y plántulas de A. germinans.
Análisis espacial de la estructura del bosque
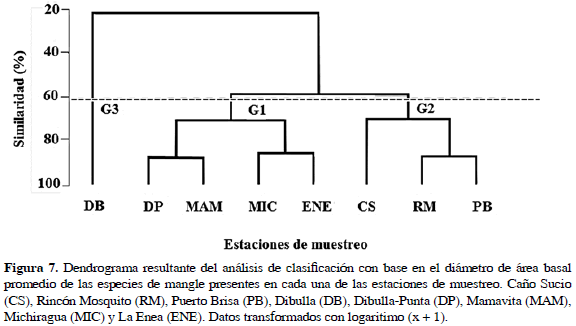
El análisis de clasificación indicó diferencias espaciales en la estructura de los bosques de mangle, mostrando la formación de dos grupos y una estación aislada (Figura 7). El primer grupo (Gl), conformado por las estaciones situadas en la zona norte del área de estudio (Dibulla-Punta, Mamavita, Michiragua y La Enea), se caracterizó por la presencia de bosques mixtos, con A. germinans como especie exclusiva. El segundo grupo (G2) lo constituyen las estaciones ubicadas hacia al sur (CS, RM y PB), caracterizándose por florestas con buen desarrollo, conformadas en su mayoría por R. mangle y L. racemosa. Aislada de los grupos se encontró la estación Dibulla (G3), ya que fue la única estación que presentó una floresta monoespecífica de C. erectus.
DISCUSIÓN
La especie más importante en la Baja Guajira fue L. racemosa, lo cual puede ser atribuido a la capacidad que tiene para crecer bajo una gran variedad de condiciones, desde suelos arenosos hasta depósitos cenagosos o arcillosos (Thom, 1967; Chapman, 1976), franjas internas donde el flujo de las mareas es limitado (Lugo y Snedaker, 1974) y tolerar un amplio ámbito de salinidad de 0 a 40 (Cintrón-Molero y Schaeffer-Novelli, 1983; Rollet, 1984). Según Araujo y Polanía (1985), L. racemosa puede crecer en asociación estrecha con otras especies dominando en condiciones de baja salinidad, dicho patrón fue evidente a lo largo del área de estudio. Sin embargo, en La Enea la especie dominante fue R. mangle, que crece en terrenos inestables asociados a las condiciones de inundación provocada por la influencia del río Tapias, favorecida por el éxito de establecimiento y desarrollo que presenta dicha especie en sustratos poco consolidados (Lugo y Snedaker, 1974).
Considerando que la baja densidad y los valores altos de área basal son indicativos de madurez (Schaeffer-Novelli y Cintrón-Molero, 1986), las estaciones de Puerto Brisa, La Enea y Rincón Mosquito presentaron los bosques más desarrollados del área de estudio dados los valores de DAP y altura promedio registrados de ellas (Tabla 3). Por el contario las formaciones de manglar presentes en las estaciones de Dibulla y Mamavita, con altas densidades y bajas áreas basales, fueron catalogadas como bosques poco desarrollados con ausencia de propágulos y escasa regeneración natural. Siendo Dibulla la estación con menor desarrollo del área de estudio, debido al crecimiento achaparrado y ramificado de C. erectus que difícilmente llega a la categoría latizal, dadas las condiciones desfavorables del suelo (compactación por la exposición directa al sol y escasa vegetación por la tala excesiva) y el déficit hídrico, dificultando el desarrollo de un sistema radicular complejo (Mizrachi et al, 1978).
En general el sector sur (CS, RM, PB y DB) con mayor influencia de aguas continentales (Sánchez-Páez et al, 1997) presenta bajas salinidades y bosques bien desarrollados dominados por L. racemosa, que disminuye en densidad y en desarrollo siguiendo un gradiente hacia el norte que recibe menor aporte de agua continental. Avicennia germinans aparece únicamente en las estaciones del norte lo cual evidencia una relación inversa con L. racemosa y concuerda con lo mencionado por Rabinowitz (1978), Cintrón-Molero y Schaeffer-Novelli (1983) y Chen y Twilley (1998), quienes afirman que el desarrollo de L. racemosa es favorecido en franjas a lo largo de canales o en las desembocaduras fluviales con baja salinidad y altos niveles de inundación, mientras que A. germinans prospera en lugares con bajos niveles de inundación y donde la salinidad puede ser mayor. Se aduce entonces que las diferencias entre los manglares del sector sur y el norte se derivan de la disponibilidad de agua dulce, si bien en ambos sectores los aportes dependen de la lluvia, en los manglares del sur desembocan mayor número de riachuelos que provienen de la SNSM, cuya boca se cierra parte del año formando pequeñas lagunas y zonas inundadas (Garzón-Ferreira y Cano, 1991).
La especie L. racemosa, debido a su plasticidad adaptativa, tiene la capacidad de implantarse y desarrollarse en diversos tipos de ambientes (Tomlinson, 1986; Perdomo, 2001; Rivera-Monroy et al., 2001), razón por la cual se observaron plántulas en todas las estaciones, presentándose la mayor densidad en la estación de Dibulla-Punta, favorecida por la presencia de abundantes neumatóforos que aumentan la retención temporal de propágulos y facilitan el anclaje de los mismos. Las bajas salinidades y la presencia de adultos maduros cercanos son los principales factores de la disponibilidad de propágulos y la densidad de las plántulas (Casas-Monroy, 1999), razón que explica que R. mangle presente reclutamiento en las estaciones inundadas donde se cumplen dichas condiciones.
La estación de La Enea tuvo la mayor densidad de propágulos de A. germinans, situación generada principalmente por la abundancia de árboles adultos en el área y porque está especie tiene mayor producción de propágulos cuando aumenta el nivel y disminuye la salinidad (Cardona y Botero, 1998). En esta estación, el nivel de agua es determinante en la sobrevivencia de las plántulas, ya que con los niveles medidos (entre 20 y 60 cm), las plantas más pequeñas mueren por inmersión debido a que disminuye la concentración de oxígeno en sus raíces (Elster et al., 1999); esta condición afecta más a las plántulas de A. germinans que a las de R. mangle, ya que esta última se presentó en mayor proporción, favorecida posiblemente por la longitud de los propágulos.
La ausencia de propágulos y plántulas de mangle en la estación Caño Sucio podría suponer efectos sobre la regeneración natural del bosque en el área, no obstante, las observaciones en campo sólo se realizaron para la época seca, periodo en el cual la especie dominante del bosque (L. racemosa) no presenta producción de propágulos (Elster et al., 1999). Por otro lado, es posible que las condiciones ambientales tales como tipo de sustrato y nivel de agua, influencien el establecimiento y sobrevivencia de plántulas y propágulos, ya que en la estación prevalecieron sustratos de arenas gruesas (franja inicial con L. racemosa) y sustratos secos muy consolidados hacia el interior del bosque, que dificultan el enraizamiento y la superviviencia de propágulos y plántulas (Elster et al., 1999). En el bosque de la estación de Dibulla, constituido exclusivamente por C. erectus, se evidenció reproducción a partir de las partes vegetativas de la planta (raíces y ramas) las cuales tienen el potencial de generar nuevos individuos cuando crecen en la superficie del suelo, proceso que evita un gasto energético mayor en condiciones ambientales extremas (Mizrachi et al., 1978).
Los bosques de mangle de la Baja Guajira ocupan un área total de 59.9 ha y presentan el mayor desarrollo en el departamento de La Guajira, atribuido principalmente a los aportes de aguas de escorrentía provenientes de la SNSM, que arrastran nutrientes producto de la actividad agrícola, es así como los bosques más desarrollados se establecen en la parte sur de la Baja Guajira donde se encuentra el mayor número de fuentes provenientes de la SNSM. No obstante, la extensión de los bosques (cobertura) no depende únicamente del número de fuentes de agua dulce sino del caudal que transportan, es por esto que las mayores coberturas de mangle de la Baja Guajira se ubican al norte, influenciadas directamente por el río de mayor extensión y caudal del área (río Tapias), mientras que al sur, los bosques de mangle sólo cubren un área de 18.75 ha. Además, a pesar que el sector sur exhibe condiciones favorables para el establecimiento y desarrollo de mangle, es objeto de fuertes presiones de aprovechamiento forestal y cambio de uso del suelo, lo cual va en detrimento de la cobertura vegetal si no se desarrollan programas de vigilancia, control y de educación ambiental.
Si bien, en el presente estudio se estima una cobertura total de mangle para la Baja Guajira de 59.9 ha, mientras que Gil-Torres et al. (2009) registra 177.66 ha, la diferencia es atribuida principalmente a la definición de la clase de cobertura Bosques de Mangle que para el presente trabajo correspondió exclusivamente a formaciones vegetales con una o más especies de mangle (A. germinans, R. mangle, L. racemosa y C. erectus), mientras que Gil-Torres et al. (2009) contempló los Bosques de Mangle como ecosistemas, incluyendo no sólo las especies de mangle sino también especies asociadas y cuerpos de agua. En adición, Gil-Torres et al. (2009) tuvieron en cuenta sectores no considerados en el presente estudio como Boca del río Lagarto-Maluisa con una cobertura de 29.05 ha.
AGRADECIMIENTOS
Este trabajo fue posible gracias al apoyo técnico, logístico y financiero del Instituto de Investigaciones Marinas y Costeras-INVEMAR. Un agradecimiento especial a Carolina Segura (Ingeniera Catastral y Geodesta GEZ-INVEMAR) por su colaboración en la interpretación de las imágenes satelitales, fotografías aéreas y elaboración de los mapas de cobertura.
BIBLIOGRAFIA
1 Alongi, D. M. 2002. Present status and future of world's mangrove forest. Environ. Conserv., 29 (3): 331-349. [ Links ]
2 Araujo, R. y J. Polania. 1985. Manglares: estructura, fitosociología, y geo-morfología en el delta del Canal del Dique (departamentos de Bolívar y Sucre, Colombia). Tesis Biología Marina. Univ. de Bogotá Jorge Tadeo Lozano, Cartagena. 126 p. [ Links ]
3 Arroyo, M. 2001. Caracterización de algunas propiedades físicas y químicas del suelo de manglar en la desembocadura del río Ranchería (Riíto), municipio de Riohacha, Caribe colombiano. Tesis Ing. Ambiental, Univ. de La Guajira, Riohacha. 83 p. [ Links ]
4 Cardona, P. y L. Botero. 1998. Soil characteristics and vegetation structure in a heavily deteriored mangrove forest in the Caribbean coast of Colombia. Biotropica, 30 (1): 24-34. [ Links ]
5 Casas-Monroy, O. 1999. Patrones de regeneración natural del manglar en la región de la Ciénaga Grande de Santa Marta, Caribe colombiano. Tesis Biol. Mar., Univ. de Bogotá Jorge Tadeo Lozano, Santa Marta. 124 p. [ Links ]
6 Chapman, V. J. 1976. Mangrove vegetation. Verlag J. Cramer. Nueva York. 600 p. [ Links ]
7 Chen, R. y R. Twilley. 1998. A gap dynamic model of mangrove forest development along gradients of soil salinity and nutrient resources. J. Ecol., 86: 37-51. [ Links ]
8 Cintrón-Molero, G. y Y. Schaeffer-Novelli. 1983. Introducción a la ecología del manglar. UNESCO/ROSTLAC. Montevideo. 109 p. [ Links ]
9 Cintrón-Molero, G. y Y. Schaeffer-Novelli. 1984. Methods for studying mangrove structure. 91-113. En: Snedaker, S. C. y J. G. Snedaker (Eds.). The mangrove ecosystem: Research methods. Monogr. Oceanogr. Meth., Vol. 8, UNESCO, París. [ Links ]
10 Cintrón-Molero, G. y Y. Schaeffer-Novelli. 1992. Ecology and management of new world mangroves. 233-258. En: Seeliger, U. (Ed.). Coastal plant communities of Latin America. Academic Press Inc., San Diego. 392 p. [ Links ]
11 Clarke, K. y R. Warwick. 2001. Change in marine communities: an approach to statistical analysis and interpretation. Plymouth University, Plymouth. 131 p. [ Links ]
12 Elster, C., L. Perdomo y M. L. Schnetter. 1999. Impact of ecological factors on the regeneration of mangrove in the Ciénaga Grande de Santa Marta, Colombia. Hydrobiología, 413: 35-46. [ Links ]
13 Espinal, L. S. y E. Montenegro. 1963. Formaciones vegetales de Colombia. Instituto Geográfico Agustín Codazzi, Bogotá. 187 p. [ Links ]
14 Garzón-Ferreira, J. y M. Cano. 1991. Tipos, distribución, extensión y estado de conservación de los ecosistemas marinos costeros de Parque Nacional Natural Tayrona. Informe técnico, INVEMAR. Santa Marta, 32 p. [ Links ]
15 Gil-Torres, W., G. Fonseca, J. Restrepo, P. Figueroa, L. Gutiérrez, G. Gómez, P. C., Sierra-Correa, M. Hernández-Ortiz, A. López y C. Segura-Quintero. 2009. Ordenamiento ambiental de los manglares de la Alta, Media y Baja Guajira, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta. 283 p. [ Links ]
16 Gómez, M. 2005. Caracterización del bosque de manglar del Santuario de Fauna y Flora Los Flamencos departamento de La Guajira, Caribe colombiano. Tesis de Biol. Mar., Univ. de Bogotá Jorge Tadeo Lozano, Santa Marta. 373 p. [ Links ]
17 Guerra, W., L. Díaz, N. González y F. Forero. 2003. La Guajira colombiana. IM editores, Bogotá. 153 p. [ Links ]
18 Lema, A. 2003. Elementos estadísticos de dasometría y medición forestal. Silvano Ltda, Medellín. 372 p. [ Links ]
19 Lozano, P. y P. Sierra-Correa. 2005. Propuesta metodológica para la delimitación de bosques de manglar a partir de datos satelitales utilizando SPRING v 4.1, en la reserva de biósfera Ciénaga Grande de Santa Marta, Caribe colombiano. Anais XII Simpósio Brasileiro de Sensoramiento Remoto, 4285-4283. [ Links ]
20 Lugo, A. y S. Snedaker. 1974. The ecology of mangroves. Ann. Rev. Ecol. Syst., 5: 39-64. [ Links ]
21 Martínez, L. y M. Ruíz. 2001. Caracterización de algunas propiedades físicas y químicas del suelo manglárico en el Valle de los Cangrejos (Brazo Calancala). Tesis Ing. Ambiental, Univ. de La Guajira, Riohacha, Colombia. 68 p. [ Links ]
22 McCune, B. y J. Grace. 2002. Analysis of ecological communities. MjM Software Design. Oregon, Estados Unidos. 300 p. [ Links ]
23 Mizrachi, D., R. Pannier y F. Pannier. 1978. Estudio de algunas características de las estrategias de propagación e implantación de Conocarpus erecta. 286-294. En: UNESCO (Ed.). Estudio científico e impacto humano en el ecosistema de manglar. UNESCO. Montevideo. [ Links ]
24 Perdomo, L. 2001. Evaluación de procesos de regeneración natural del manglar después de la reapertura de los caños de agua dulce (Ciénaga Grande de Santa Marta, Colombia). Tesis de Maestría en Ciencias-Biología Marina, Univ. Nacional de Colombia, Santa Marta. 106 p. [ Links ]
25 Rabinowitz, D. 1978. Dispersal properties of mangrove propagules. Biotropica, 10 (1): 47-57. [ Links ]
26 Ricerca e Cooperazione, Unión Europea y Corporación Autónoma Regional de La Guajira. 2002. Plan de uso y manejo de los humedales y lagunas costeras de La Guajira: Informe final. Proyecto Prolagunas: Protección y recuperación de humedales costeros del Caribe colombiano, CD-ROM. Riohacha. [ Links ]
27 Rivera-Monrroy, V., E. Mancera, R. Twilley, O. Casas-Monrroy, E. Castañeda, J. Restrepo, F. Daza-Monrroy, L. Perdomo, P. Reyes, E. Campos, M. Villamil y F. Pinto. 2001. Estructura y función de un ecosistema de manglar a lo largo de una trayectoria de restauración: El caso de la región de la Ciénaga Grande de Santa Marta. Informe final, Univ. of Louisiana-Lafayette-INVEMAR, Santa Marta. 224 p. [ Links ]
28 Rollet, B. 1984. Ecología de los manglares con especial referencia a la base biológica para la ordenación sostenida, forestal y pesca. Memorias del Seminario Nacional sobre Ordenación Integrada de Manglares. FAO, Roma. 33 p. [ Links ]
29 Rosado, J., E. Márquez, L. Cortes, J. Moscote y A. Deluque. 2003. Caracterización vegetal y ecológica del Valle de Los Cangrejos, Riohacha. Tesis de Ing. Ambiental, Univ. de La Guajira, Riohacha. 83 p. [ Links ]
30 Sánchez-Páez, H., R. Alvarez-León, F. Pinto-Nolla, A. Sánchez-Alférez, J. Pinto-Rengifo, H. García-Hansen y M. Acosta-Peñalosa. 1997. Diagnóstico y zonificación preliminar de los manglares del Caribe de Colombia. Ministerio del Medio Ambiente, Bogotá. 511 p. [ Links ]
31 Sánchez-Páez, H., G. Ulloa-Delgado, R. Alvarez-León, W. Gil-Torres, A. Sánchez-Alférez, O. Guevara-Mancera, L. Patiño-Callejas y F. Páez-Parra. 2000. Hacia la recuperación de los manglares del Caribe de Colombia. Ministerio del Medio Ambiente, Bogotá. 294 p. [ Links ]
32 Schaeffer-Novelli, Y. y G. Cintrón-Molero. 1986. Guía para estudo de areas de manguezal: estrutura, funjao e flora. Caribbean Ecological Research, Sao Paulo. 150 p. [ Links ]
33 Thom, G. 1967. Mangrove ecology and deltaic geomorphology, Tabasco, México. J. Ecol., 55: 301-343. [ Links ]
34 Tomlinson, P. 1986. The botany of mangroves. Cambridge University Press, Nueva York. 419 p. [ Links ]
35 Ulloa-Delgado, G., H. Sánchez-Páez, W. Gil-Torres, J. C. Pinto-Rengifo, H. Rodríguez-Cruz y R. Alvarez-León. 1998. Conservación y uso sostenible de los manglares del Caribe colombiano. Ministerio del Medio Ambiente, Bogotá. 224 p. [ Links ]
FECHA DE RECEPCIÓN: 29/04/2010 FECHA DE ACEPTACIÓN: 23/04/2011
*Contribución No. 1086 del Instituto de Investigaciones Marinas y Costeras - INVEMAR.
