











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCTION
The Eremophilus mutisii is a silurid endemic to the Colombian Cundinamarca-Boyacá highlands which is classified as a threatened species, mainly due to aspects related to the deterioration of its habitat (Mojica et al. 2012; Álvarez & Rodríguez 2007). The interest in recovering this species in systems in which it has disappeared and, to some extent, the potential for its cultivation and commercialization, are circumstances that motivate the development of different research initiatives, among which those that address different aspects of reproductive management stand out (Landines P. et al. 2017).
There is consensus on the environmental importance of the species and its potential for aquaculture (Rodríguez Forero, 2000; Rosado-Puccini, 2007), the latter based on criteria of recognition and acceptability, as well as size and quality of the meat; it is a feasible candidate to expand the current fish farming offer for cold water systems in the high Andean region of Colombia, whose only representative is the rainbow trout (Oncorhynchus mykiss).
Regardless of the approach, both fundamentally converge on the need to develop the technical elements that make it possible to have breeding programs that allow, in practice, a constant supply of fry of the E. mutisii; although there are advances in this line, indeed relevant aspects related to the response to induction and management in incubation and larviculture are not yet consolidated (Rosado-Puccini, 2007). Only preliminary trials have been conducted on the maintenance of specimens in captivity, with no published references, and the work on induced reproduction, in most cases, is still being carried out with adult specimens from natural habitats.
On the other hand, the determination of the general composition aspects of gametes, incorporating fatty acid profiles, and the relationship with reproductive indexes is a valid approach to defining quality criteria for eggs and fry (Kjørsvik et al. 1990; Bobe 2015). In particular, fatty acids are part of the analyses on structure, and composition, because they influence maturation, reproduction, egg quality, hatching, early survival, and larval growth (Brooks et al. 1997). Knowledge of the general egg composition is a mechanism that allows inferences about the relationship between environmental supply and female nutrition, in addition to early fry performance outcomes. While it is the characteristics of the diet that ultimately modulate the compositional parameters of an egg, as recorded for several species (Tocher, 2010), the constituents that will ultimately be present are related to the nutritional resources to which females have had access to during the ovarian maturation process (Wiegand 1996), although, depending on the system, there are variations in the diet due to changes in the quality and quantity of the food supply (Tolussi et al. 2018; Lopes de Souza et al. 2020).
The gonads and muscle are tissues frequently used to determine normal ranges in aspects of body composition for a given species. During the reproductive season lipids and proteins are mobilized from the muscle to the gonads (Tolussi et al. 2018), so the nutrition of the breeder will modulate, among others, the supply of essential fatty acids in the eggs that, in the end, are a determining part of reproductive success (Sharma et al. 2024). Therefore, compositional aspects thus become an analytical reference that allows to deduce or project the nutritional quality of the food supply to which they had, or should have, access to specimens destined for breeding (Izquierdo et al. 2001; Glencross, 2009). Possible deficiencies or differences can be detected, whether they are associated with a particular diet, or with the conditions in which the fish developed or were kept, considering that there is a seasonal availability of different types of food in the environment (Petenuci et al. 2021).
The approach to the fatty acid composition of eggs in specimens from the environment constitutes an input to determine how these should be considered in the formulation of diets designed for captive breeders (Rainuzzo, 2020), being a particularly useful and practical approach in cases where there are no references for a given species, as in the case of the E. mutisii.
Accordingly, this study describes the fatty acid composition of eggs and muscle in female E. mutisii and analyzes the possible variation of the profiles between different capture sites that are currently sources of broodstock.
MATERIALS AND METHODS
Experimental fish. The specimens were obtained from three sites, all in the Bogotá river basin. Of these, there were two lentic sites and one lotic: the Tominé reservoir, located near Sesquilé and Guatavita; the Fúquene lagoon, located between the municipalities of Susa and Fúquene; and finally in the axis of the Bogotá River in its upper course, in the municipality of Suesca. The capture period coincided with the rainy season in the central region of the country, and the sampling was concentrated in two months.
In all cases, fishing was carried out using hooks, obtaining a total of 42 specimens, distributed as follows: 12 females and 8 males in Tominé, 9 females in Suesca, and 11 females and 2 males in Fúquene. It should be noted that the collection of wild specimens for non-commercial purposes is authorized for the Universidad Nacional de Colombia by Resolution 000697 of the National Environmental Licensing Authority of Colombia (ANLA).
On the other hand, for the destination of the waste from the specimens worked on, the collection, transfer, and disposal protocols established for this purpose at the University were followed.
Six specimens were selected from each locality based on the criteria of fully developed ovaries, occupying a large part of the coelomic cavity, and with eggs with a diameter greater than 1000 µm, one of the indicators used to identify individuals suitable for reproduction induction procedures (Moncaleano Gómez et al. 2018). In the laboratory, the females were euthanized by overexposure to the anesthetic MS 222 (400 ppm), the total length in mm (approximation to 1 mm) and weight (approximation to 0.1 g) were determined; during dissection, the complete ovaries and liver were removed and weighed (approximation to 0.1 g).
For fatty acid analyses, a 5 g sample was taken from the middle portion of the right ovary and a 10 g sample of muscle from the dorsal muscle of the animal. The samples were stored at -70˚C until processing (Valbuena-Villarreal et al. 2013; Rosado-Puccini et al. 2014).
Fatty acid profile. The samples were freeze-dried prior to lipid extraction, which was performed according to the methodology of Folch et al. (1957) and the chromatographic determination of fatty acids was carried out according to that described by Betancourt et al. (2005). For this purpose, each sample of eggs and muscle was homogenized in a chloroform and methanol solution, filtered and the solvent was centrifuged at 2000 rpm for 15 minutes, taking the organic phase to dryness under vacuum at room temperature. Methylesterification was carried out using MethPrep II reagent (Alltech Associates Inc., Deerfield, IL, USA) and the determination was performed in a Shimadzu® GC-14A gas chromatograph, using helium as carrier gas. Fatty acid methyl esters were identified by comparing their retention times with those of a standard fatty acid mixture (Supelco 37 component FAME Mix, Supelco Inc., Bellefonte, PA, USA).
The profiling of eggs and muscle from each female was performed for 19 and 18 fatty acids respectively; totals corresponding to the monounsaturated (MUFA), polyunsaturated (PUFA) and saturated (SFA) groups were calculated. Those of the n-3 and n-6 series are recorded, in addition to the n-3/n-6 ratios.
Data analysis. Using the records of weight, length, and weights of gonads and liver, the condition factor (K) and the gonadosomatic (GSI) and hepatosomatic (HSI) indexes were calculated for each specimen; the data are presented as mean ± standard deviation (SD) and, using analysis of variance (ANAVA) (p<0.05), it was determined if there were differences between capture sites.
Likewise, the values of each fatty acid are expressed as the mean ± standard deviation (SD). An ANOVA (p<0.05) was performed, and Tukey's test was conducted to compare means. The software SPSS V25 statistical was used for analysis.
RESULTS AND DISCUSSION
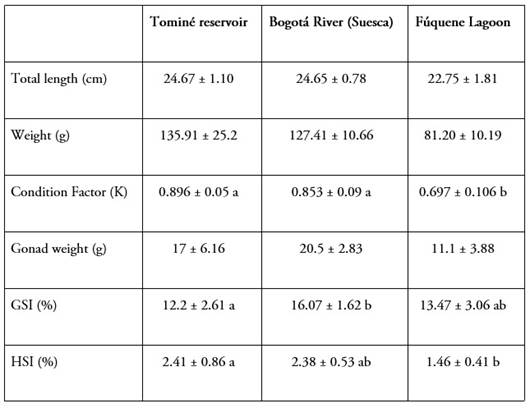
The average total length of the 18 females studied was 24 ± 1.53 cm, ranging from 20 to 26.3 cm; the weight was 114.8 ± 29.4 g, ranging from 71.7 to 176.6 g; the group of fish from the Fúquene lagoon had the lowest total length and weight records. The mean data for the different variables for the selected females are presented in Table 1.
The size, in length and weight, of the females that were selected to obtain the samples is within the intervals that correspond to fish that have reached first maturity, which, in the species, as recorded by Pinilla et al. (2006), it is presented in sizes of 14 to 20 cm in total length and between 40 and 50 g in weight.
For the Tominé reservoir and the Bogotá River, the sizes of the females were similar (Table 1); this translates into a Condition Factor that did not present significant differences between the two sites and, in addition, the mean K for these sites is close to those reported by Moreno et al. (2019) for specimens from the same area of the Bogotá River. The same does not occur with the specimens from the Fúquene lagoon where, regardless of the smaller size presented by the females chosen, the K value was significantly lower (Table 1). However, since the condition of the fish is related to seasonal variations, it is feasible that, for this system, the values recorded are more a consequence of the time of capture and the particularities of the available food supply than to any other circumstance (Lopes de Souza et al. 2020).
Rodríguez Forero (2000), Álvarez & Rodríguez (2007) and Andrade-López (2006) describe the species as omnivorous, generalist in terms of feeding strategy, which suggests a broad trophic spectrum and that individuals within the same population are capable of using different resources at the same time. This characterization of habits and the variable supply of different food items in a system throughout an annual cycle explains the variations that occur in K, both among the mean values of the population in each season and among individuals in that system.
Beyond the mean sizes of individuals, in the possible relationship of individual condition with reproductive indicators, the variation in the GSI of the fish among the three sites showed that the lowest value was presented for the Tominé reservoir (Table 1). Even so, the range of 12.2% at this site, up to 16.7% determined for the Bogotá River, are within the range and are in accordance with those evidenced with maturing or mature specimens recorded by Landines P. et al. (2017) for fish from the Suesca area, also in rainy periods. This interval also contains the value of 13.6% that Moreno et al. (2019) report for maturing females in the same area.
Table 1 Morphometric records and body and reproductive indexes in E. mutisii females from each of the three localities
Data for each site are expressed as mean ± SD (n=6). Different letters between rows of K, gonadosomatic index GSI and hepatosomatic index HSI indicate significant differences (p<0.05)
Regarding the HSI, there is also a lower, significant value in females from Fúquene, with 1.46%; in Tominé (2.41%) and Suesca (2.38%), the records were higher and without differences between them. Regardless of these differences, in all cases the mean values were higher than those recorded by Moreno et al. (2019) in the range of 0.4% to 1.9% for specimens from Suesca, an evaluation that included mature females. Specifically, the general analysis of condition relationships with reproductive status indicators in the set of females chosen suggests a similar state of maturation among localities; this circumstance validates the results, taking into account that the provision of fatty acids to the egg has vitellogenin as a vehicle (March, 1993), incorporated during vitellogenesis in ovarian development.
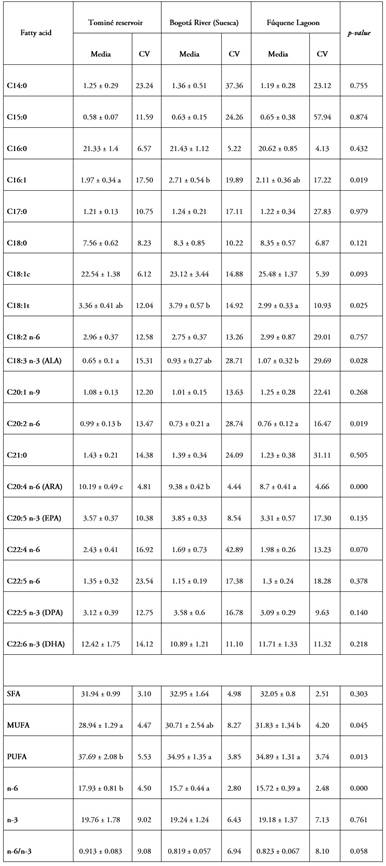
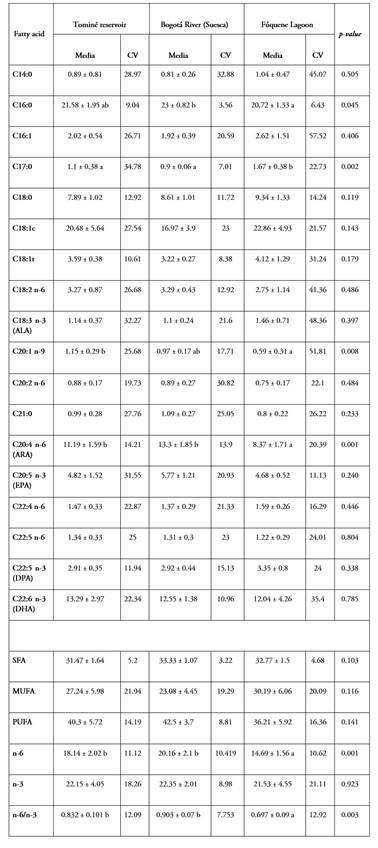
In a global view of the fatty acid profiles, the emerging picture is one of stability in the contents across localities. In fact, of the total fatty acids determined in the eggs, only four showed significant differences between sites; except for arachidonic acid (C20:4 n-6, with an average of 9.42%), the content of the other three acids was less than 5% and with high variability between females. A similar situation is also present in muscle, since out of 18 acids, four have differences among the zones, two of them with contents lower than 5%, with palmitic (21.77% on average) and arachidonic (with an average of 10.95%), better represented in content and with differences among the sites (Tables 2 and 3). The importance of arachidonic acid in reproduction and egg and seed quality in fish is well established and widely documented (Wu et al. 2021).
Table 2 Mean content (%) and coefficient of variation (%) of each fatty acid analyzed for egg samples in female E. mutisii from the three locations.
Data for each fatty acid is expressed as mean ± SD (n=6). Different letters between rows indicate significant differences (p<0.05). Monounsaturated - MUFA, polyunsaturated - PUFA and saturated - SFA
Table 3 Mean content (%) and coefficient of variation (%) of each fatty acid analyzed for muscle samples in female E. mutisii from the three locations.
Data for each fatty acid is expressed as mean ± SD (n=6). Different letters between rows indicate significant differences (p<0.05). Monounsaturated - MUFA, polyunsaturated - PUFA and saturated - SFA
Comparison of the content of each fatty acid present at the three sites, performed between egg and muscle averages, showed differences in myristic (C14; 1.267% and 0.91%; p=0.013), eneicosyl (C21; 1.391% and 0.969%; p=0.006), eicosapentaenoic (C20: 5n-3; 3.574% and 5.091%; p=0.016) and in the n3 set (19.393% and 22.01%; p=0.001).
It is also consistent that the highest contents among the SFA are with palmitic (C16:0), with oleic (C18:1c) among the MUFA and with docosahexaenoic (DHA, C22:6n-3) among the PUFA. There are no differences in the mean contents of these acids among localities and the variation among females in each zone is not high (CV<15%). The sum of these three acids represents between 55.4% (Bogotá River, Suesca) and 57.8% (Fúquene lagoon) of the total present in the ovaries. This absolute value, based on the sum of the contents of the three fatty acids, is similar to that reported for rainbow trout (O. mykiss) by Rosado Puccini et al. (2012), in which they reach up to 61%, and for red tilapia (Oreochromis sp.) by Valbuena-Villarreal et al. (2013), with 69% for the same group of acids. It is interesting to note that, in both trout and tilapia, species that are at the extremes of a feeding continuum, there is similarity in the absolute content of these three fatty acids and that this condition is conserved in the E. mutisii, which has an omnivorous diet with a carnivorous tendency (Álvarez & Rodríguez 2007; Rojas et al. 2018).
It is also noteworthy that, for all three species, the amounts of palmitic (C16:0) and oleic (C18:1) acids lie within narrow ranges, from 21 to 26% and 18 to 22%, respectively. This high relative weight in the content distribution is explained by their role and importance in sexual maturity (Sharma et al. 2024), in addition to being mobilized and incorporated during vitellogenesis, since these acids are used as an energy source during the embryonic process and until exogenous feeding begins. In this regard, Tocher (2003) mentions that the use of oleic acid is manifested in a significant and progressive decrease in its concentration as embryonic development advances. A similar utilization is recorded for palmitic acid in rainbow trout embryos, indicating its important role as an energy source in early development (Yeganeh, 2014). It is noteworthy that this fatty acid is the saturated fatty acid with the highest content in both muscle (between 20.72 and 21.43%) and eggs (between 20.62 and 23.08%), with similar means.
Regardless of the dietary and nutritional regime, minimum levels appear to be maintained for certain acids, even in the case that the nutrition scheme is based on diets that vary in different composition and balance (Tocher, 2010). Its importance is reaffirmed when the distribution of contents is usually similar, even among species of completely different habitats, habits and locations, which has been documented in several cases (Glencross, 2009). To illustrate the variety of species that maintain a similar pattern in the content of these three acids, examples include the Asian freshwater silurids such as Ompok pabda and Wallago attu (Mukhopadhyay & Ghosh, 2007), the estuarine fish Centropomus undecimalis (Yanes-Roca et al. 2009) and the Atlantic salmon Salmo salar (Pickova et al. 1999).
In this regard, Sargent et al. (2003) and Tocher (2003) state that the composition of fatty acids in eggs is usually conserved and less influenced by diet than other tissues, which reflects the importance of maintaining a specific composition in the gametes; provided that a diet is not particularly deficient in any component and minimum contents are maintained, the mobilization of fatty acids towards the gametes allows stable levels of concentration to be maintained at the end, which has been reported in different species (Sargent et al. 2003; Rosado Puccini et al. 2012; Valbuena-Villarreal et al. 2013; Rosado-Puccini et al. 2014). The correspondence observed in fatty acid concentrations between eggs and muscle for the E. mutisii seems to confirm these selective retention mechanisms (Wiegand 1996; Sargent et al. 2003), especially when the samples evaluated come from females captured in completely different systems.
The set of saturates was found to be equivalent among localities, both for eggs (32.3%) and muscle (32.5%). In the monounsaturated fatty acids (MUFA) in eggs, differences between localities were present, but with a variation of 2.9% between the two extremes (28.9% in the Tominé reservoir and 31.8% in Fúquene); this is a narrow range, lower than the 8.7% reported for the same set in red tilapia () and 8.9% in rainbow trout (Rosado Puccini et al. 2012). Although in total monounsaturates in muscle were not different among zones, with an average of 26.8%, the dispersion among females reached a coefficient of variation of about 20%. A similar result is shown by the polyunsaturated group, which is best represented in both egg and muscle content. With differences between localities and a variation of 2.8% in eggs, the maximum occurs in the Tominé reservoir (37.7%) and the minimum in Fúquene (34.9%); on the other hand, in muscle there are no differences between localities, but with an average of 39.7%, the variation among females ranges from 8.8 to 16.3%.
The comparative approach of fatty acid profiles between eggs from captive and wild specimens can be an indicator of the quality of diets provided to broodstock when fry production programs are maintained in farmed species; for the particular case of E. mutisii, considering the circumstances that currently define the management and the progress of research on the species, the profiles found can offer an adequate baseline for future work on broodstock nutrition (Sharma et al. 2024).
The relative stability observed in muscle and egg contents, as well as between localities, suggests that these profiles are a reference point for formulating and evaluating nutritional schemes, at least regarding fatty acids are concerned. It is clear that the next steps for the control of reproductive management and obtaining fry in the E. mutisii must necessarily consider the reduction of dependence on wild catches in the environment and achieve the maintenance of broodstock in captivity conditions; thus, these results allow us to advance, in complement with evaluations that contemplate other nutrients, in the development of possible diets for the species.

