











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Los bosques periurbanos, frecuentemente, son amenazados por diversas causas, entre las más importantes, los procesos de migración humana, el crecimiento demográfico, el aumento de oferta inmobiliaria, la construcción de vías de acceso, el turismo, entre otros. Estas modificaciones resultan prácticamente irreversibles, por lo que son una amenaza sobre las escasas áreas verdes urbanas, suprimiendo por completo funciones ecológicas, como la regulación climática, la purificación del aire y agua y, en el caso de bosque inundables, los procesos hidrológicos de regulación y almacenamiento del recurso hídrico (Seto et al. 2011; Moschella, 2023; Cavallero et al. 2024). Teniendo en cuenta lo anterior, se considera prioritario realizar esfuerzos para conservar, recuperar y restaurar estos ecosistemas estratégicos para la supervivencia de las grandes ciudades (Moschella, 2023).
Garwood (1989) afirma que los dos mecanismos principales que contribuyen a la regeneración de los ecosistemas son el banco de semillas y las semillas recientemente dispersadas; sin embargo, considera que es más importante la contribución del banco de semillas. Los bancos de semillas pueden ser transitorios, es decir, aquellos donde las semillas permanecen viables por un periodo aproximado de un año o persistentes, en donde las semillas permanecen viables durante años.
La recuperación de la vegetación inicia con la expresión del banco de semillas, proceso que se encuentra relacionado con factores bióticos, abióticos y procesos ecológicos; entre los factores bióticos más sobresalientes y prácticamente condicionantes es la existencia de propágulos en el medio, como agentes dinamizadores naturales de la sucesión (Hill et al. 2001; Bedoya-Patiño et al. 2010; Muñoz, 2017).
La regeneración natural después de un disturbio requiere de la presencia de semillas o propágulos en el medio; en este proceso se presentan fases, como la producción y dispersión de semillas, la germinación y establecimiento de las plántulas, Se considera que los estados más vulnerables durante este ciclo son las semillas y las plántulas, debido a que las condiciones bióticas y abióticas en el ecosistema pueden generar alta mortalidad de estas (Norden, 2014).
La dispersión de las semillas es una estrategia de las plantas para colonizar nuevos lugares donde las condiciones sean adecuadas para las especies; sin embargo, esto no siempre ocurre, muchas veces, las semillas no encuentran las condiciones necesarias para la germinación y el establecimiento (Norden, 2014). Los estudios sobre lluvia de semillas brindan información sobre cómo el bosque responderá a ciertos cambios ambientales, utilizando su propio potencial regenerativo (Calegari et al. 2013).
En áreas degradadas, los bancos de semillas se encuentran empobrecidos o ausentes, según el grado de deterioro y cuando existen están dominados por especies pioneras herbáceas, dando como resultado una lenta recuperación y una baja diversidad del ecosistema (Vargas et al. 2009; Muñoz et al. 2017, Foresto et al. 2021). Cuando esto ocurre, es necesario iniciar procesos de restauración ecológica asistida, donde se incorporen al medio especies nativas, que dinamicen el proceso de recuperación.
En este sentido, en áreas de interés donde se requiera iniciar acciones de manejo, es muy importante conocer cómo ocurre la regeneración natural de las especies y sus procesos ecológicos asociados, con el fin de contar con herramientas que permitan diseñar estrategias de manejo sostenible y de restauración ecológica más eficientes (Harms & Paine, 2003; Martins, 2016; Muñoz et al. 2017; Poorter et al. 2023).
Teniendo en cuenta lo anterior, el Jardín Botánico de Bogotá inició acciones de restauración ecológica en el Bosque las Mercedes, desde el 2011. Esta área es un importante relicto de bosque inundable, que hace parte de la estructura ecológica principal de Bogotá; por lo tanto, esta investigación buscó analizar la composición, la abundancia, la densidad, la riqueza y la diversidad de la lluvia y el banco de semillas, entre coberturas vegetales con diferentes años de intervención y, de esta manera, proporcionar información sobre el avance en el proceso de restauración ecológica en el Bosque Las Mercedes (Figura 1), ya que estudios han demostrado que el conocimiento del banco y la lluvia de semillas de un área permite enterarse del estado de sucesión, el grado de perturbación y las posibles fuentes de propágulos (Padilla, 2008; Solorza-Bejarano, 2012; Fernández et al. 2016; Solorza-Bejarano, 2017).
MATERIALES Y MÉTODOS
Área de estudio. El Bosque Las Mercedes se encuentra ubicado en la localidad de Suba (Figura 1a), entre las coordenadas N 4º46’, W 74º05’ a 2.554 m s.n.m.; se considera un ecosistema de gran importancia para la ciudad, debido a que es reconocido como el último relicto de Bosque andino montano bajo (bh-MB), de planicie lacustre inundable y valle aluvial del río Bogotá. A pesar de su importancia ecológica, el área se ha enfrentado a una continua degradación antrópica, debido a las actividades productivas realizadas, presentando disturbios, como la fragmentación, la pérdida de biodiversidad y la invasión por especies arvenses trepadoras; por lo anterior, fue establecida como un Área Piloto de Investigación en Restauración Ecológica APIRE para el Jardín Botánico de Bogotá (JBB) (Pérez, 2016).
Las acciones de restauración ecológica han sido implementadas por el JBB desde el 2012 y han consistido en la erradicación de especies vegetales invasoras, oportunistas y trepadoras agresivas; el enriquecimiento del sotobosque con la adición de especies nativas; la plantación de especies pioneras arbóreas y arbustivas en áreas de pastizal y la formación de corredores ecológicos en zonas invadidas por pastizales de Cenchrus clandestinus (Hochst. ex Chiov.) Morrone (Pérez, 2016; Pérez, 2020).
Dentro del área, denominada Bosque Las Mercedes, el JBB definió una zonificación, de acuerdo con el estado de su vegetación, encontrándose la cobertura de bosque, que se refiere a la zona mejor conservada dentro del ecosistema (Tabla 1); sin embargo, posee diferentes tensionantes que han sido abordados. En esta cobertura se han identificado tres subdivisiones: Bosque denso (BD), Bosque abierto (BA) y Bosque Inundable (BI), en donde se ha realizado el control de especies, como Rubus bogotensis Kunth, Muehlenbeckia tamnifolia (Kunth) Meisn., Salpichroa tristis Walp., Cucurbita ficifolia Bouché, entre otras y la adición de especies vegetales nativas, propias del ecosistema de bosque inundable (Pérez, 2016; Pérez, 2020).
La cobertura de Pastizales Misceláneos (PMI) se encuentra alrededor del bosque, presenta especies exóticas e invasoras, en su mayoría herbáceas, pertenecientes a la familia Poacea, como C. clandestinus y Holcus lanatus L., especies nativas que han sido plantadas en los procesos de restauración ecológica y otras que han arribado, provenientes del bosque o de otras áreas anexas (Tabla 1). Esta cobertura fue subdividida teniendo en cuenta el periodo de intervención: PMI01 (en esta área se realizó la intervención en enero del 2021), PMI3A (intervención en febrero del 2020) y PMI3B (intervención desde junio del 2017) (Figura1b).
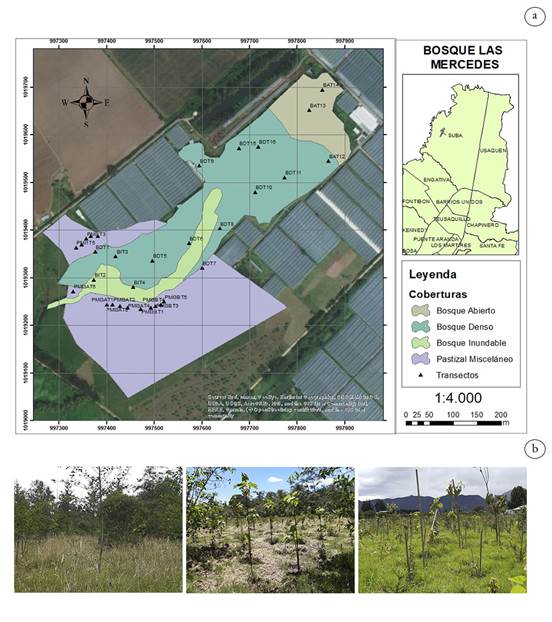
Figura 1 a) Mapa ubicación área de estudio Bosque Las Mercedes. b) Fotografías Pastizales de izquierda a derecha: PMI3B- Pastizal Misceláneo (intervención desde 2017); PMI3A - Pastizal Misceláneo (intervención desde marzo de 2020) y PMI1- Pastizal Misceláneo (intervención desde enero de 2021).
La investigación se desarrolló evaluando la lluvia de semillas y el banco de semillas; asimismo, se realizó la caracterización de la vegetación en pie, con el objetivo de identificar su correspondencia, similitudes o diferencias.
Caracterización vegetal: se llevó a cabo por medio de 31 transectos de 25 x 4 m (16 en bosque y 15 en pastizal), con el fin de conocer la vegetación en pie, tanto adultos como lo individuos en proceso de desarrollo; se registraron todos los individuos de porte arbóreo y arbustivo presentes, con un DAP ≥ a 2,5 cm (Campo Kurmen, 2010); para las herbáceas, se registró el porcentaje de cobertura y para las plántulas reclutadas de especies de hábito arbustivo y arbóreo fueron contabilizadas y registradas. Asimismo, sobre estos transectos, se tomaron las muestras para el banco de semillas y se ubicaron las trampas para la evaluación de la lluvia de semillas.
Banco de semillas germinable - BSG: la evaluación se realizó con la recolección de 310 submuestras de suelo de 5 cm de altura x 5 cm de diámetro, en mayo del 2021; en cada transecto, se tomaron 10 submuestras, para conformar dos muestras por transecto, para un total de 62 muestras compuestas. Estas se secaron parcialmente a temperatura ambiente; se eliminaron, de modo manual, los restos de raíces, hojas y piedras y se dispusieron uniformemente, en bandejas de germinación plásticas, ubicadas dentro de un microinvernadero, localizado al sur de Bogotá, donde se presentó una temperatura interior promedio de 25 °C y humedad del 90 %; con frecuencia, se suministró riego a capacidad de campo. El análisis de la densidad y composición del banco de semillas se realizó por el método de germinación directa, durante un periodo de cinco meses, donde cada 15 días se registró el número de plántulas por morfoespecie y en la identificación se trató, en lo posible, de llegar hasta especie (Moscoso Marín & Diez Gómez, 2005).
Lluvia de semillas: se evaluó por medio de trampas, en forma de cono invertido, con un diámetro de 40 cm; la boca de la trampa fue ubicada a un metro de altura. El muestreo de la lluvia de semillas se realizó mensualmente, durante junio y julio del 2021; las semillas de cada trampa se recolectaron en bolsas plásticas, se llevaron al laboratorio de semillas de la subdirección científica del JBB para ser secadas a temperatura ambiente, procesadas y observadas bajo el estereoscopio. La identificación se realizó con la ayuda de ejemplares de herbarios, carpotecas y comparación con semillas recolectadas en la zona; los frutos recolectados en las trampas se abrieron para contabilizar sus semillas.
Dentro de las tres coberturas localizadas en el bosque se instaló una trampa ubicada en la mitad del transecto (a 12,5 m), para un total de 16 trampas; en los pastizales misceláneos se evaluó la lluvia de semillas a diferentes distancias del borde del bosque, por lo tanto, en cada transecto se ubicaron tres trampas de semillas: la primera, a los 6 m de distancia del borde; la segunda, a los 12 m y la tercera, a los 24 m, con el objetivo de conocer el aporte de la lluvia de semillas provenientes desde el bosque, a diferentes distancias en los pastizales circundantes (Tercero-Bucardo et al. 2010).
Análisis de los datos: la densidad de las semillas se calculó por metro cuadrado; la diversidad se evaluó a partir de los índices de Shannon y Simpson y la riqueza, a partir del índice de Margalef. En el análisis de diversidad Beta para la comparación entre los bancos, la lluvia de semillas y la vegetación en pie dentro de las coberturas estudiadas, se empleó el método ordenación de escalamiento multidimensional no métrico con distancia de Bray - Curtis, el cual, no solo tiene en cuenta las especies, sino que, además, incluye las abundancias de forma ponderada, es decir, esta medida no considera las ausencias compartidas como similares (Rojas-Jiménez, 2022). Finalmente, para la comparación entre los resultados obtenidos entre las diferentes coberturas y mecanismos se realizó el análisis de componentes principales con el programa estadístico Past (Hammer et al. 2001), donde se comparó las especies y sus abundancias en cada mecanismo y cobertura.
RESULTADOS Y DISCUSIÓN
Composición de la vegetación en pie. Para el total del área de estudio se encontraron 5.908 individuos de 115 especies, pertenecientes a 100 géneros y 51 familias botánicas (Tabla 3). La familia con mayor abundancia de individuos fue la Solanaceae, seguido de Asteraceae y Poaceae. Las especies con mayor abundancia fueron: Solanum oblongifolium Humb. & Bonpl. ex Dunal, C. clandestinus, Smallanthus pyramidalis (Triana) H. Rob., Vallea stipularis ex L. fil. y R. bogotensis.
En la cobertura de bosque se obtuvo 4.393 individuos de 96 especies, pertenecientes a 87 géneros y 49 familias botánicas. La cantidad de individuos obtenidos en esta cobertura corresponde al 79 % del total de los individuos registrados; las familias más representativas de la cobertura fueron: Solanaceae, Asteraceae, Rosaceae y Elaeocarpaceae. La mayor parte de las especies registradas corresponde a arbustos 40,5 % y árboles 27,4 %; las trepadoras representaron el 16,4 % y las herbáceas y gramíneas 15,7 %. En cuanto a las especies, se observó que las mayores abundancias las presentaron S. oblongifolium, R. bogotensis, S. pyramidalis, V. stipularis y Bocconia frutescens L.
En la cobertura de pastizal misceláneo se encontraron 1.515 individuos de 76 especies, distribuidas en 67 géneros y 37 familias botánicas; las familias con mayor representatividad fueron Poaceae, Asteraceae, Fabaceae y Araliaceae. La mayor abundancia la presentaron las especies herbáceas, con un 31,4 % y gramíneas, con 30,8 %; asimismo, los árboles representaron 23,2 y los arbustos 9,9 % de la muestra. Por último, solo el 4,7 %, lo presentaron las trepadoras.
Las especies herbáceas más frecuentes fueron C. clandestinus, Hydrocotyle andina Cuatrec, Trifolium pratense L., Taraxacum officinale Weber ex F. H. Wigg. y Holcus lanatus L., entre otras y en cuanto a las especies más abundantes del componente arbóreo y arbustivo, se encontraron S. pyramidalis, V. stipularis, Myrcianthes leucoxyla (Ortega) Mc Vaugh y Abatia parviflora Ruiz & Pav.
Para la cobertura de Bosque se observó que R. bogotensis fue la segunda especie más abundante; se trata de una especie colonizadora agresiva (Pérez, 2020; Muñoz, 2022), que ha sido identificada como un tensionante en el Bosque Las Mercedes, al competir por luz con otras especies, tanto arbóreas como arbustivas (Pérez, 2016; Pérez, 2020; Muñoz, 2022). En este sentido, los resultados obtenidos en la presente investigación difieren con lo encontrado por Pérez (2020), quien observó que, a raíz del control de R. bogotensis, se había logrado disminuir la dominancia de la especie en las coberturas de Bosque. Por otra parte, para esta misma cobertura, la alta abundancia de S. oblongifolium podría ser un factor tensionante para la regeneración de otras especies nativas, ya que se encuentra compitiendo por espacio y luz en el sotobosque.
En el caso de las especies arbóreas y arbustivas encontradas en los pastizales misceláneos corresponden a especies que han sido incorporadas en los procesos de restauración ecológica desarrollados (Pérez, 2016) y no por procesos de regeneración natural, es decir, la dominancia de las gramíneas invasoras no ha permitido el establecimiento de nuevos individuos de las especies nativas plantadas o provenientes del bosque.
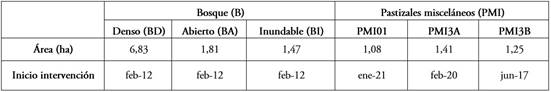
De acuerdo con Mora-Donjuán et al. (2017), los índices de diversidad para la vegetación en pie mostraron que para Margalef la riqueza fue alta para todas las coberturas, al encontrarse valores cercanos a 5 o superiores, especialmente, para BD, PMI3B y PMI1 y el índice de Shannon (H) fue mayor en el sector de BA; sin embargo, este valor se considera como diversidad media, debido a que se reporta un valor entre 2 - 3 (Tabla 2). Los valores obtenidos para este índice muestran que algunas pocas especies presentan abundancias mayores al promedio en todas las coberturas. Para el índice de Simpson (1-D), todas las áreas presentaron una alta diversidad, con valores cercanos a 1 (Martínez, 2023). En cuanto a la densidad de individuos por hectárea fue mucho mayor en la cobertura de BD, seguido por BI y las menores densidades se presentaron en las dos zonas más recientemente intervenidas.
Tabla 2 Riqueza e índices de diversidad para las zonas estudiadas en el Bosque Las Mercedes.
BD- Bosque Denso; BA- Bosque Abierto; BI- Bosque Inundable; PMI3B- Pastizal Misceláneo (intervención desde 2017); PMI3A - Pastizal Misceláneo (intervención desde marzo de 2020) y PMI1- Pastizal Misceláneo (intervención desde enero de 2021).
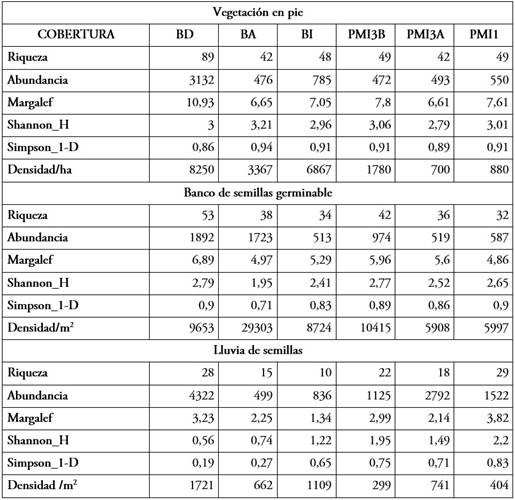
En el análisis de diversidad beta para la comparación entre las coberturas y los sectores para la vegetación en pie, el índice de Bray - Curtis mostró que el bosque y los pastizales misceláneos son diferentes (Figura 2a), incluso, dentro de la cobertura de bosque, se observó que BA presenta un comportamiento diferente; se encontraron algunas semejanzas entre BD y BI. También fue visible que los transectos BDT7 y BIT2 poseen una alta semejanza con la vegetación de los pastizales; dentro de estos últimos, se apreciaron diferencias significativas entre PMI1 comparado con PMI3A y PMI3B. En general, las coberturas tienen media similitud en cuanto a la composición y abundancia de especies (R=0,70), indicando que las zonas son diferentes y de acuerdo con el análisis estadístico no paramétrico de una sola vía ANOSIM, se obtuvo diferencias significativas entre las coberturas analizadas (P= 0,0001).
Composición del banco de semillas. El seguimiento al banco de semillas germinable se realizó durante 5 meses y se obtuvo un total de 6.208 plántulas de 78 morfoespecies, de las cuales, se logró identificar 66 especies pertenecientes a 28 familias botánicas (Tabla 3). En la cobertura de bosque se obtuvo 4.128 plántulas y en pastizal misceláneo 2.080; las primeras especies en germinar pertenecieron al estrato herbáceo de las familias botánicas Solanaceae, Rubiaceae, Asteraceae y Poaceae.
Las semillas germinadas en la cobertura de bosque pertenecen a 65 especies de 26 familias botánicas; las más abundantes fueron Solanaceae, Rubiaceae y Asteraceae. En estas tres familias se encuentra el 84 % de las semillas germinadas; la mayor parte de las semillas germinadas pertenece al estrato herbáceo, con un 81,7 %, seguido del estrato arbóreo, con 6,7 % y arbustivo, con 5,9 %; las especies trepadoras y gramíneas, con un 4,2 % y 1,5 %, respectivamente.
Las especies más frecuentes fueron: Oldenlandia corymbosa L., Solanum nigrescens M. Martens & Galeotti, Solanum americanum Mill., Erigeron bonariensis L., Physalis peruviana L., R. bogotensis, V. crassiramea, S. pyramidalis y S. oblongifolium. Esto coincide con lo reportado por Gioria & Pyšek (2016) y Abreu et al. (2021), quienes observaron que los bancos de semillas estaban compuestos, en su mayoría, por especies herbáceas de sucesión temprana.
En la cobertura de pastizal misceláneo se observaron 58 morfoespecies, correspondientes a 25 familias botánicas; las familias más representativas en esta cobertura fueron Rubiaceae, Asteraceae, Poaceae y Solanaceae. En estas 4 familias se encuentra representado el 78 % de las semillas germinadas; la mayor abundancia de semillas germinadas corresponde al estrato herbáceo, con el 47,3 %; las gramíneas, con 20,22 %; las trepadoras, con 17 %; los árboles, con un 10,4 % y, finalmente los arbustos, con un 5,1 %.
Las especies más abundantes en orden fueron: Galium murale (L.) All., O. corymbosa, H. lanatus, S. nigrescens, V. crassiramea, T. pratense, C. clandestinus y Baccharis latifolia (Ruiz & Pav.) Pers. Las dos primeras especies de la familia Rubiaceae han sido consideradas invasoras en otros países (Causton et al. 2020; Pauchard et al. 2020) y H. lanatus es considerada invasora para Colombia (Díaz-Espinosa et al. 2012; Mora-Goyes & Barrera-Cataño, 2015).
En cuanto a las especies invasoras o colonizadoras agresivas, se encontró que, en el banco de semillas, una especie muy abundante fue R. bogotensis; también se observó Muehlenbeckia tamnifolia (Kunth) Meisn., en menor proporción. Estas dos especies se encontraron, tanto en bosque como en pastizales, asociadas, principalmente, a BD, BI, PMI3A y PMI3B. Por otra parte, una especie muy abundante en todas las coberturas, en particular, en BD y BA, fue el S. oblongifolium y en pastizales se observó únicamente de forma abundante en PMI3A.
En este sentido, Gioria & Pyšek (2016) y Velosa et al. (2018) observaron que las especies invasoras tienen un efecto negativo sobre la densidad y la riqueza de especies nativas en el banco de semillas, ya que sugieren que la biomasa producida por estas impide la llegada de semillas de especies nativas de comunidades vecinas al suelo. En efecto, las mayores densidades de semillas se presentaron en BA y en PMI3B (Tabla 2); sin embargo, esto se debe a la abundancia de las especies de la familia Rubiaceae. Asimismo, la menor densidad de semillas se presentó en PMI1 y PMI3A, que corresponde a las zonas más recientemente intervenidas con acciones de restauración ecológica, coincidiendo con Vargas et al. (2009), Muñoz et al. (2017) y Foresto et al. (2021), quienes afirman que en áreas, fuertemente degradadas, los bancos de semillas se encuentran empobrecidos o ausentes.
Las coberturas PMI01 y BA presentaron los menores valores para los índices de Shannon y Simpson (Tabla 2); sin embargo, corresponde a una diversidad intermedia (Mora-Donjuán et al. 2017; Martínez, 2023). Asimismo, para todas las coberturas, estos índices presentaron valores intermedios; en el caso de BA, obtuvo la mayor densidad de semillas germinadas, pero estas correspondían a pocas especies, como O. corymbosa, por esta razón, presentó el menor valor para el índice de Simpson. Por otra parte, las coberturas que mostraron una mayor riqueza y diversidad son las coberturas de PMI3B, BD y BI, donde el índice de Margalef y el índice de Simpson mostraron mayor valor; asimismo, presentaron una alta densidad de semillas. En este sentido, Ma et al. (2019) encontraron que los bancos de semillas persistentes de alta diversidad y densidad son un indicador importante de la alta capacidad de recuperación del ecosistema después de un disturbio.
En el análisis de diversidad Beta se observó que dentro del bosque se presentan diferencias marcadas entre las coberturas BD y BA; en la cobertura pastizal misceláneo se observaron diferencias entre las zonas PMI1y PMI3A, en contraste con PMI3B, ya que este último comparte características con BD (Figura 2b). En general, las coberturas tienen de baja a media similitud en cuanto a la composición y la abundancia de especies (R=0,5753), indicando que las zonas son diferentes. De acuerdo con el análisis ANOSIM, se obtuvo diferencias significativas entre las coberturas analizadas (P=0,0001).
Figura 2 Ordenación multivariada con índice de similitud de Bray - Curtis. a) vegetación en pie, b) banco de semillas y c) lluvia de semillas.
Composición de la lluvia de semillas. Durante junio y julio del 2021, se recolectaron en total 11.096 semillas de 48 morfoespecies, de las cuales, se logró la identificación de 28 especies de 19 familias botánicas (Tabla 3). Las familias con mayor representatividad fueron Betulaceae, Asteraceae, Poaceae y Solanaceae; la especie más abundante fue Alnus acuminata Kunth, con 5.810 semillas, que corresponden al 52 % de semillas recolectadas, seguida de V. crassiramea, con 1.604, S. pyramidalis, con 839 y H. lanatus, con 716 semillas.
Tabla 3 Especies vegetales presentes en cada fase de regeneración natural y cobertura en el Bosque Las Mercedes.
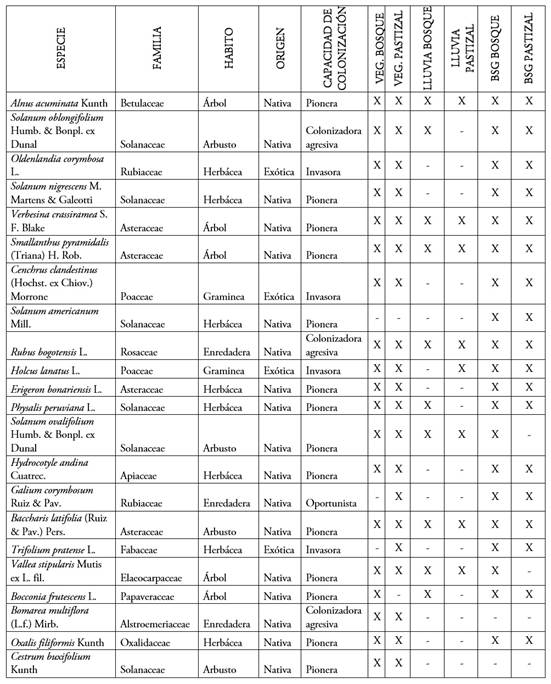
Las especies se encuentran ordenadas, de acuerdo con su abundancia total de mayor a menor (X para especie presente en cada etapa de la regeneración y cobertura y - especie ausente en etapa de la regeneración y cobertura).
En la cobertura bosque se colectaron 5.657 semillas, distribuidas en 36 morfoespecies, de las cuales, se identificaron 22 especies pertenecientes a 16 familias botánicas; las más abundantes fueron Betulaceae, Asteraceae, Solanaceae y Rosaceae. En estas familias se encuentra el 97 % de las semillas recolectadas. El estrato arbóreo fue el más abundante, con un 94,6 %; seguido estrato arbustivo, con un 3,7 %; las trepadoras, con 1,3 %; las herbáceas, con un 0,4 % y las gramíneas no estuvieron presentes en la lluvia de semillas del bosque. En cuanto a las especies, la más abundantes fueron A. acuminata, seguida de V. crassiramea, S. pyramidalis, S. oblongifolium y R. bogotensis.
En la cobertura de Pastizal Misceláneo se presentaron 5.439 semillas, se distribuyeron en 39 morfoespecies, de las cuales, se identificaron 23 especies de 16 familias botánicas; la más abundante fue la familia Asteraceae, seguido de la familia Betulaceae, Poaceae y Solanaceae; en estas familias se encuentra el 83 % de las semillas recolectadas. En esta oportunidad, los estratos dominantes fueron el arbóreo y arbustivo, con el 59,1 % y 20,7 %, respectivamente; las gramíneas ocuparon el tercer lugar, con 13,2 %; las trepadoras, con 5,8 % y las herbáceas, con 1,2 %. Las especies más abundantes para esta cobertura fueron V. crassiramea, A. acuminata, H. lanatus y S. pyramidalis.
Los resultados de densidad promedio de semillas por m2 demostraron que el mayor valor fue observado en la cobertura de BD, con 1.721 semilla/m2 (Tabla 2); la mayoría correspondiente a A. acuminata y S. pyramidalis, seguida de BI, con 1.109 semillas/m2, con las especies A. acuminata, V. crassiramea y S. oblongifolium y en tercer lugar, se encuentra PMI3A, donde la especie más abundante fue V. crassiramea. Las coberturas con menor densidad de semillas/m2 fueron en PMI3B y PMI1.
El índice de Margalef mostró una riqueza baja en la cobertura BI, los demás sectores presentaron media riqueza. El índice de Simpson mostró alta y media diversidad para los transectos de PMI1 y PMI3B y baja diversidad para BD y esto se debe a la alta abundancia de semillas de A. acuminata; según Shannon la diversidad fue baja para todos los sectores (Tabla 2).
En el análisis de la diversidad Beta se observó que dentro de la cobertura de bosque no se encuentran diferencias marcadas (Figura 2c). En cuanto al pastizal misceláneo se observó que PMI3A y PMI3B son similares a las coberturas de BD; por otra parte, la cobertura PMI1 presenta un comportamiento diferente a la cobertura de bosque. Se observó que, aproximadamente, el 80 % de la muestra de semillas se recolectó en la trampa ubicada a 6 m del borde del bosque; el 15 %, en las trampas ubicadas al 12 m y el 5 % restante, en las trampas ubicadas a 24 m; esto demuestra que, a mayor distancia del bosque, menor es el aporte de semillas proveniente de este sobre el pastizal, en concordancia con lo reportado por Alcalá et al. (2014). En general, las coberturas presentaron baja similitud en cuanto a la composición y abundancia de especies (R=0,373). De acuerdo con el análisis estadístico ANOSIM, se obtuvo diferencias significativas entre las coberturas analizadas (P= 0,0001).
Comparación entre mecanismos y vegetación. En la comparación entre las especies y sus abundancias de las variables del banco, lluvia de semillas y la vegetación en pie, se observó que el componente 1 representó el 29,5 % de los datos; el componente 2, el 27,4 % y el componente 3, el 17,5 %, entre los tres se logró capturar el 74,4 % de los datos (Figura 3). Las variables banco de semillas y lluvia de semillas mostraron una mayor representación; los vectores explican cada una de las variables, que están compuestas por el mecanismo de regeneración evaluado y la cobertura.
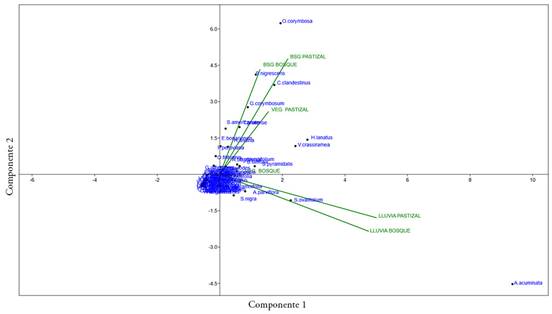
Figura 3 Componentes principales las especies vegetales y sus abundancias en la vegetación en pie (VEG), banco (BSG) y lluvia de semillas en el Bosque Las Mercedes y áreas anexas.
El análisis mostró que existe una correlación muy alta y positiva para las especies y su abundancia entre los bancos de semillas del bosque y de los pastizales misceláneos aledaños, es decir, hay una alta semejanza entre estos. También, se observa una relación alta y positiva de los bancos de semillas con la vegetación de los pastizales; estas semejanzas están dadas por la presencia y la abundancia de especies herbáceas, como O. corymbosa, S. nigrescens, S. americanum, C. clandestinus, H. lanatus, Galium corymbosum Ruiz & Pav, entre otras y de especies arbóreas y arbustivas, como V. crassiramea, S. pyramidalis, S. oblongifolium y B. latifolia y la trepadora R. bogotensis, principalmente.
Por otra parte, se observó una correlación alta y positiva entre la lluvia de semillas para las dos coberturas analizadas y estuvo dada, principalmente, por la presencia y la abundancia de la especie A. acuminata. La vegetación del bosque presenta una correlación media y positiva, tanto con los bancos de semillas como con la vegetación en pastizales y la lluvia de semillas; con este último, la relación es un poco más estrecha y se debe a que comparten especies arbóreas y arbustivas, como A. acuminata, S. ovalifolium, A. parviflora y S. nigra, entre otras.
Por último, se observa que no se presentan correlaciones, es decir, que actúan como variables independientes, la lluvia de semillas del bosque y pastizal, con el banco de semillas en la cobertura de bosque, el banco de semillas de pastizal y la vegetación de pastizal, es decir, en el agrupamiento dado por las especies y sus abundancias, es visible la división entre la lluvia de semillas en las dos coberturas, de los bancos de semillas y vegetación en pie (Figura 3).
Como conclusión, en los pastizales misceláneos, las especies exóticas e invasoras, como son C. clandestinus y H. lanatus, presentan las mayores abundancias y coberturas en la vegetación en pie y el banco de semillas. En el caso de la lluvia de semillas, la especie que se encontró en forma abundante fue H. lanatus, especialmente, en la cobertura PMI3B. En este sentido, Shono et al. (2007) y Muñoz et al. (2017) encontraron que, en áreas fuertemente degradadas, donde los fenómenos de compactación del suelo, escases de lluvia de semillas, baja fertilidad del suelo y colonización agresiva de pastos exóticos, provocan que la regeneración natural no se dé o se presente de forma muy lenta; esto se puede apreciar en la composición de vegetación en pie y el banco de semillas, donde estas especies herbáceas invasoras continúan dominando, a pesar del aporte de semillas de especies nativas pioneras de hábito arbóreo y arbustivo desde el bosque y las especies plantadas en los procesos de restauración ecológica.
Al comparar el comportamiento del banco y la lluvia de semillas se observó que existen diferencias significativas entre las coberturas; sin embargo, también es visible algunos comportamientos similares entre el sector BD y PMI3B, que corresponde al sector donde se han desarrollado actividades de restauración ecológica desde el 2017; de acuerdo con los gráficos de ordenación multivariada, se confirma una mayor afinidad en cuanto a las especies y sus abundancias en el banco de semillas de estas dos coberturas. Asimismo, el gráfico elaborado para la vegetación en pie también mostró que PMI3B presenta semejanza con BD, más específicamente, con el transecto BDT7 y en la lluvia de semillas con PMI3A y PMI3B.
En los pastizales misceláneos, al desarrollar proyectos de restauración ecológica, se han plantado algunas especies dinamizadoras del proceso de sucesión vegetal, de rápido crecimiento y abundante producción de frutos (Pérez, 2016); por este motivo, se encuentran algunas semejanzas entre estas coberturas con bosque. En el caso de PMI3B, algunas de las especies plantadas se encuentran en producción de frutos y semillas, como A. acuminata, S. pyramidalis y A. parviflora, entre otras, por lo tanto, se observan en la lluvia y el banco de semillas.
La riqueza y la diversidad de especies fue alta para la vegetación en pie y el banco de semillas, tanto para el bosque como para los pastizales misceláneos y media a baja, para la lluvia de semillas; esto se presenta porque solo algunas especies pueden estar en dispersión, en el momento de la toma de datos.
Es importante exponer que la investigación también permitió determinar que la cobertura de bosque no corresponde a un ecosistema maduro, ya que, como se observó, presentan diferencias en cuanto a composición de especies y abundancias entre los diferentes sectores estudiados, especialmente, entre bosque denso y bosque abierto; además, la alta presencia de especies invasoras, colonizadoras agresivas u oportunistas, en el banco, la lluvia de semillas y la vegetación en pie, están causando modificaciones en esta cobertura, donde por competencia por la luz, están desplazando a otras especies nativas, importantes para este ecosistema.
Por otra parte, se observó que especies invasoras, colonizadoras agresivas u oportunistas frecuentes en el bosque, como R. bogotensis, O. corymbosa, S. nigra y M. tamnifolia, se encuentran formando parte del banco y la lluvia de semillas en los pastizales misceláneos, especialmente, en los sectores donde se ha observado un mayor avance en el proceso de restauración ecológica, como son PMI3B y PMI3A.
De acuerdo con los resultados obtenidos, se considera que esta información resulta valiosa para el seguimiento a los procesos de restauración ecológica en el Bosque Las Mercedes, ya que se logró establecer que el análisis de las fases de la regeneración pueden ser consideradas como indicador del proceso de restauración ecológica en las coberturas vegetales, teniendo en cuenta que las áreas que fueron objeto de acciones de restauración ecológica asistida, desde hace más tiempo, ya muestran algunas semejanzas con la trayectoria sucesional más avanzada de bosque denso, mientras la cobertura con menor tiempo de intervención (PMI1), aún presenta comportamientos de zonas fuertemente antropizadas.
Se recomienda que en las áreas de cobertura de bosque y áreas anexas se continúen desarrollando acciones de restauración ecológica, específicamente, en lo referente al control de especies invasoras, colonizadoras agresivas u oportunistas (Tabla 3) y de regeneración natural asistida, donde se busque favorecer el desarrollo de las especies nativas que se expresan, a través del banco de semillas.

