










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.31 no.1 Bogotá Jan./June 2009
Revision of the genus Sporobolus (Poaceae: Chloridoideae: Sporobolinae) for northwest South America: Peru, Ecuador, Colombia, and Venezuela
DIEGO GIRALDO-CAÑAS
PAUL M. PETERSON
Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá D. C., Colombia. dagiraldoc@unal.edu.co
Department of Botany, National Museum of Natural History, Smithsonian Institution, Washington, DC 20013-7012, U.S.A. peterson@si.edu
RESUMEN
Se presenta un estudio taxonómico de las especies de Sporobolus para el noroeste de Sudamérica. Se reconocen once especies para el área de estudio. Se incluyen las claves para determinar las especies así como las descripciones morfológicas y sus ilustraciones. Se discuten para cada una de las especies sus relaciones morfológicas, su distribución geográfica y sus preferencias ecológicas; adicionalmente, se relacionan sus usos y sus nombres comunes. Asimismo, se propone la lectotipificación para Sporobolus lasiophyllus Pilg. Por otra parte, se registran tres especies por primera vez para Colombia [Sporobolus pilifer (Trin.) Kunth, Sporobolus tenuissimus (Mart. ex Schrank) Kuntze y Sporobolus virginicus (L.) Kunth]. Se excluyen de este tratamiento a Sporobolus brasiliensis (Raddi) Hack. (=Eragrostis airoides Nees) y Sporobolus domingensis (Trin.) Kunth. Adicionalmente, se propone la adopción y la unificación de algunos términos morfológicos en agrostología, tales como caña, panoja, espiguilla, lema, antecio y cariopsis, en lugar de culmo, panícula, espícula, lemma, flósculo y cariopse-cariópside, respectivamente.
Palabras clave. Sporobolus, Sporobolinae, Chloridoideae, Poaceae, Gramíneas neotropicales, Flora de Sudamérica.
ABSTRACT
A taxonomic treatment of Sporobolus for northwest South America is given. Eleven species are recognized in the study area. Keys for determining the species, morphological descriptions, and illustrations are provided. Morphological relationships, geographical distributions, ecological preferences, common uses, and vernacular names are given. Sporobolus lasiophyllus Pilg. is lectotypified. Sporobolus pilifer (Trin.) Kunth, Sporobolus tenuissimus (Mart. ex Schrank) Kuntze, and Sporobolus virginicus (L.) Kunth are reported in Colombia for the first time. Sporobolus brasiliensis (Raddi) Hack. (=Eragrostis airoides Nees) and Sporobolus domingensis (Trin.) Kunth are excluded from this treatment.
Key words. Sporobolus, Sporobolinae, Chloridoideae, Poaceae, Neotropical grasses, Flora of South America.
INTRODUCCIÓN
La subfamilia Chloridoideae comprende 146-166 géneros y 1360-1500 especies (Van den Borre & Watson 1997, Hilu & Alice 2001, Liu et al. 2005a, 2005b, Peterson et al. 2007), principalmente representados en las regiones tropicales y subtropicales de ambos hemisferios, siendo frecuentes en ambientes abiertos de suelos salinos y de condiciones xéricas y subxerofíticas, así como en áreas típicamente ruderales. A esta subfamilia pertenece el género Sporobolus R. Br., el cual se distribuye en los trópicos y los subtrópicos con algunos representantes en las regiones templado-cálidas y comprende aproximadamente 160 especies (Nicora & Rúgolo de Agrasar 1987, Baaijens & Veldkamp 1991, Tovar 1993, Pohl et al. 1994, Simon & Jacobs 1999, Watson & Dallwitz 1999, Ortiz-Díaz & Culham 2000, Lægaard & Peterson 2001, Peterson et al. 1995, 1997, 2003, 2004), de las cuales 76 especies (de éstas 72 son nativas) crecen en el Hemisferio Occidental (Peterson et al. 2003, 2007). Las especies de Sporobolus, por lo general, crecen en suelos secos, salinos o alcalinos, arenosos o arcillosos, tanto de sabanas, páramos y praderas como de playas y ambientes alterados, entre el nivel del mar y los 3900 m de altitud.
Sporobolus es, junto con Eragrostis Wolf, uno de los géneros más heterogéneos de la subfamilia Chloridoideae (Van den Borre & Watson 1997). Este género se caracteriza por sus pequeñas espiguillas unifloras, perfectas, glabras, sin aristas, las cuales están comprimidas lateralmente o en ocasiones son redondeadas, en las que la raquilla está articulada por encima de las glumas; sus glumas son membranáceas a escariosas, traslúcidas y brillantes, generalmente uninervias o enervias, a veces con nervaduras laterales débiles, siendo frecuentemente la gluma inferior más corta que la gluma superior y que el antecio; la lema y la pálea son subiguales y de igual consistencia a la de las glumas; la lema es 1 (3)-nervia, mientras que la pálea es 2-nervia; por su parte, sus dos lodículas son cuneadas y uninervias y el fruto está libre en el antecio y corresponde a un utrículo sin surco ventral y con hilo punctiforme.
El presente aporte pretende contribuir al conocimiento de la rica flora neotropical y en especial, al de su diversa aunque escasamente conocida flora agrostológica. Cabe destacar que Sporobolus no cuenta con una monografía continental (véase de Castro Boechat & Longhi-Wagner 1995) y además, este género es, después de Festuca, uno de los géneros más "problemáticos" de las Poaceae (Ortiz-Díaz & Culham 2000). Aquí presentamos la revisión taxonómica de las especies de Sporobolus para el noroeste de Sudamérica, área en la que el género está representado por once especies.
MATERIALES Y MÉTODOS
Estudios exomorfológicos. Se incluyeron en el análisis los caracteres vegetativos y reproductivos, fundamento importante para la comparación de los taxones. Las técnicas utilizadas corresponden a las clásicas en taxonomía vegetal. La terminología de las estructuras morfológicas planas y tridimensionales está basada en los términos propuestos por Font Quer (1993), excepto en lo que tiene que ver con las palabras "caña" [= culmo sensu Chase & Luces de Febres (1972), Sousa & Zárate (1988), Dávila Aranda & Manrique Forceck (1990) y Pinto-Escobar (2002)], "panoja" [= paniculodio sensu Rua (1999), panícula sensu Chase & Luces de Febres (1972), Sousa & Zárate (1988), Dávila Aranda & Manrique Forceck (1990), Gould & Shaw (1992) y Font Quer (1993)], "espiguilla" [= florescencia sensu Rua (1999), espícula sensu Font Quer (1993) y Pinto-Escobar (2002)], "lema" [= lemma sensu Parodi (1964), Chase & Luces de Febres (1972) y Nicora & Rúgolo de Agrasar (1987)], "antecio" [= flósculo sensu Chase & Luces de Febres (1972), Sousa & Zárate (1988) y Dávila Aranda & Manrique Forceck (1990)], y "cariopsis" [= cariopse sensu Nicora & Rúgolo de Agrasar (1987); = cariópside sensu Gould & Shaw (1992) y Pinto-Escobar (2002)], siguiéndose en este caso - salvo en las excepciones antes detalladas - a Parodi (1964), McClure & Soderstrom (1972), Zuloaga & Soderstrom (1985), Nicora & Rúgolo de Agrasar (1987), Gould & Shaw (1992), Vegetti & Pensiero (1993) y Mora-Osejo (2004), ya que éstas son las formas castellanas adoptadas en la agrostología moderna. Cabe destacar que la dimensión de la panoja está medida desde el punto de origen de la primera ramificación florífera del pedúnculo hasta el final del eje principal.
Estudio de material de herbario. Se estudiaron colecciones de diferentes herbarios, como por ejemplo CAUP, COAH, COL, HUA, LE, LINN, MEDEL, MO, PSO, S, SI y US, abreviados de acuerdo con Holmgren et al. (1990). Asimismo, se realizaron varias salidas de campo con el fin de enriquecer las colecciones de gramíneas y observar las plantas en su hábitat. Las obras originales de las descripciones de las especies aquí consideradas se tomaron de la bibliografía disponible, del "International Plant Name Index Query IPNI" (www.ipni.org), de la base de datos "W3-TROPICOS" del Jardín Botánico de Missouri (EE.UU, www.mobot.org) y del Catalógo de las gramíneas del Nuevo Mundo (Peterson et al. 2001). Véase Peterson et al. (2001) para una lista actualizada de los sinónimos de las especies aquí tratadas. Cabe destacar que las palabras holotipo, isotipo, lectotipo, isolectotipo y otras similares, están escritas de acuerdo con Kiesling (2002: xi), autor que adopta y aconseja la forma sin tilde.
RESULTADOS Y DISCUSIÓN
Caracteres morfológicos
Hábito y sinflorescencia. La sinflorescencia se define como el sistema de vástagos floríferos producidos a partir de la yema apical del eje embrional o de una yema de innovación en un período de crecimiento (Rua 1999). Las plantas son extremadamente variables y diversas en su hábito; éstas pueden ser anuales o perennes, rizomatosas, raramente estoloníferas y conspicuamente cespitosas, desde gráciles hasta robustas.
La estructura de las plantas de Sporobolus se ajusta al modelo generalizado de las Poaceae, en el que cada planta está constituida por una serie de vástagos equivalentes de orden de ramificación sucesiva. Estos vástagos muestran la secuencia de diferenciación típica de las sinflorescencias y pueden ser tratados como ejes de innovación. En cada vástago se puede diferenciar una porción distal que lleva exclusivamente paracladios florales, denominada antotagma, de una proximal provista de nomofilos, el trofotagma. En la región del trofotagma se pueden distinguir, a su vez, una zona proximal, cuyos entrenudos permanecen cortos, de otra distal en la que se alargan notablemente. La zona proximal constituiría una zona de innovación (denotada tipológicamente por las letras IZ), mientras que la distal representaría una zona de inhibición (HZ). Por su parte, el antotagma constituye la inflorescencia y hace parte de la zona de enriquecimiento o zona paracladial (BZ).
Cañas. Las cañas son herbáceas y simples y raramente pueden ramificarse, situación evidenciable principalmente en los nudos inferiores. Éstas pueden ser decumbentes a más frecuentemente erectas. Como dato curioso, merece destacarse que en ocasiones las cañas muestran alternancia de macroblastos y braquiblastos, naciendo, a partir de los nudos de estos últimos, hojas de vainas muy apretadas, con algunas raíces desarrolladas en la base del respectivo nudo. Esta situación sólo la observamos en S. lasiophyllus y únicamente en dos ejemplares procedentes de Perú [Peterson et al. 21870, 21879 (US)].
Hojas. Las hojas pueden ser gráciles a robustas y el follaje es principalmente basal y menos frecuentemente caulinar; las hojas son simples y no presentan aurícula. Cabe destacar que en S. virginicus el follaje es conspicuamente dístico. Por su parte, las vainas pueden ser redondeadas o carinadas; las lígulas corresponden a una diminuta membrana ciliada, en la que los tricomas son extremadamente cortos y rígidos. Las láminas son aplanadas o plegadas. Las características de la anatomía foliar del género y de varias de las especies aquí documentadas, pueden encontrarse en Goossens (1938), Schwabe (1949), Ellis (1977) y Baaijens & Veldkamp (1991).
Inflorescencia. Las inflorescencias pueden ser generalmente exertas o estar parcialmente incluidas en las vainas superiores; éstas varían desde panojas amplias (piramidales a oblongas), densas o hasta contraídas (espiciformes), caso último en el que las ramificaciones (paracladios) se muestran adpresas al eje principal y éstas pueden ser alternas o estar dispuestas en verticilos o pseudoverticilos. Los paracladios pueden variar enormemente en cuanto a su número por panoja y además, pueden aparecer ramificados o no, o sea, puede haber paracladios de órdenes consecutivos. Las ramificaciones son floríferas desde la base o sólo hacia su porción distal, y en ocasiones éstas pueden ser glandulares como en S. pilifer y S. pyramidatus.
En el género, así como en la familia, la inflorescencia es politélica (Liu et al. 2005a, Reinheimer et al. 2005). No obstante, esta afirmación se podría prestar a confusión toda vez que el eje principal y las ramificaciones (paracladios) terminan en una espiguilla uniflora, lo que llevaría a pensar en un caso de monotelia (Reinheimer et al. 2005). Sin embargo, las flores de las gramíneas son siempre laterales, ya que la presencia de la pálea define la condición lateral de las mismas (Cialdella & Vega 1996, Soreng & Davis 1998), toda vez que las páleas son homólogas a los profilos y éstos sólo se desarrollan en ramificaciones axilares (Nicora & Rúgolo de Agrasar 1987, Soreng & Davis 1998) y así, la flor no es verdaderamente terminal, por lo tanto, las inflorescencias son politélicas. Al igual que en otros géneros de Poaceae, la inflorescencia además de ser politélica no es truncada, pues hay florescencia principal (o sea, el eje principal remata en una espiguilla) casi sin excepciones, así como co-florescencia (los paracladios terminan en una espiguilla). Véanse Liu et al. (2005a) y Reinheimer et al. (2005) para ampliar el análisis de la configuración tipológica y morfológica de la inflorescencia en el género Sporobolus y en las Chloridoideae.
Espiguillas. Las espiguillas son unifloras (un único antecio por espiguilla), perfectas, glabras y sin aristas, las cuales están comprimidas lateralmente o en ocasiones son redondeadas; éstas pueden ser subsésiles o pediceladas, en las que la raquilla está articulada por encima de las glumas; cada espiguilla tiene dos glumas desarrolladas, las cuales son iguales, subiguales o marcadamente desiguales en longitud, membranáceas a escariosas, traslúcidas y brillantes, generalmente uninervias o enervias (muy raramente en algunas poblaciones colombianas de S. purpurascens la gluma superior es 2 ó 3-nervia), a veces con nervaduras laterales débiles, siendo frecuentemente la gluma inferior más corta que la gluma superior y que el antecio. Cabe destacar, además de la sencillez de las espiguillas en cuanto al número y la forma de las brácteas (glumas, lema y pálea) (Fig. 1A), el pobre ornamento micromorfólogico y anatómico de las mismas (Fig. 1B), el cual se limita básicamente a la presencia de células epidérmicas largas, cuerpos de sílice y papilas simples). Esta situación del pobre ornamento ya había sido destacada para Sporobolus por Valdés-Reyna & Hatch (1991).

Las espiguillas son casmógamas, aunque muy raramente puede haber espiguillas cleistógamas en una misma planta; cuando hay espiguillas cleistógamas, éstas pueden ser axilares (incluidas en las vainas foliares) o expuestas; así, el primer caso corresponde al tipo "Ia" de cleistogamia, mientras que las espiguillas cleistógamas expuestas (ubicadas en inflorescencias exertas) corresponden al tipo "IV" propuestos por Campbell et al. (1983). A las espiguillas cleistógamas incluidas en las vainas, Connor (1979, 1981) las llama "espiguillas clandestinas", las cuales son una forma especializada y son más avanzadas que las espiguillas cleistógamas de inflorescencias exertas (Connor 1981). En la familia Poaceae se conocen 14 géneros con espiguillas clandestinas (Connor 1981, Giraldo-Cañas 2008).
Cabe destacar que la cleistogamia ha sido registrada en 70 géneros de gramíneas (Connor 1979), y ésta es usualmente facultativa (Connor 1979, 1981, 1987), puesto que espiguillas casmógamas son registradas generalmente en la misma especie (Connor 1979). La presencia o ausencia de flores cleistógamas está directamente relacionada, tal como ocurre con los caracteres vegetativos, con la adaptación de las plantas a condiciones ambientales desfavorables (Connor 1981, Morrone & Zuloaga 1991) y según Astegiano (1986), la cleistogamia puede responder a causas de diverso orden: morfológico, fisiológico, genético o ambiental. Campbell et al. (1983) señalan que alrededor del 60% de las especies de gramíneas con cleistogamia son colonizadoras de ambientes modificados, lo que concuerda con los ambientes en los que crece S. indicus. Estos autores (Campbell et al. 1983) resaltan que la cleistogamia intensifica las ventajas potenciales de la autogamia.
En Sporobolus indicus a pesar de que puede haber espiguillas casmógamas y cleistógamas en una misma planta, no se encuentran diferencias morfológicas entre las mismas - espiguillas isomorfas - (glumas, lemas, antecios) ni entre los diferentes órganos florales (lodículas, estambres, polen, estilos y estigmas), tal como lo documentó Astegiano (1986). Por su parte, en otras especies del género pueden encontrarse diferencias evidentes en el tamaño de las anteras entre espiguillas casmógamas y cleistógamas, siendo más pequeñas en las espiguillas cleistógamas.
Antecio. El antecio se define como la casilla floral formada por la lema y la pálea (éstas dos constituyen las glumelas), dentro de la cual se halla una única flor (Parodi 1964, McClure & Soderstrom 1972, Nicora & Rúgolo de Agrasar 1987), cuya función consiste en proteger la flor y fundamentalmente la formación del fruto [véanse Roth (1977) y Cialdella & Vega (1996)]. En Sporobolus el antecio posee una flor perfecta, y en éste la lema y la pálea son subiguales y de igual consistencia a la de las glumas, siendo la lema 1-nervia (raramente 3-nervia), mientras que la pálea es 2-nervia; por su parte, cada antecio posee dos lodículas (glumélulas), las cuales están libres y son glabras, cuneadas y uninervias. Cabe destacar que en Sporobolus los nervios de la pálea están muy apartados, siendo el espacio internerval de textura delgada, el cual se rompe fácilmente al madurar el fruto. Esta situación de rompimiento es más fuerte en unas especies, en las que el ápice de la pálea se rompe en dos mitades, de tal manera que la pálea se muestra como si fuera dos páleas uninervadas. Cada flor posee tres estambres, aunque raramente este número puede reducirse a dos.
Fruto. El fruto está libre en el antecio y corresponde a un utrículo sin surco ventral y con hilo punctiforme (Colbry 1957). Éste puede variar de subgloboso, obovoide-elíptico a oblongo. En vista de que el fruto presenta el pericarpio libre, éste fue nombrado por Brandenburg (2003) como una cariopsis modificada. Cabe destacar que el color del fruto es siempre café y de forma subglobosa, obovoide-elíptica a oblonga, siendo plano en la cara ventral y redondeado en la dorsal (véanse Colbry 1957, Liu et al. 2005b). Por su parte, el embrión corresponde a cerca de la mitad de la longitud del utrículo. En el género Sporobolus el fruto presenta el pericarpio mucilaginoso y cuando está en contacto con el agua deja salir la semilla fácilmente (Nicora & Rúgolo de Agrasar 1987).
Géneros afines a Sporobolus
Diferentes análisis fenéticos así como filogéneticos (Clayton & Renvoize 1986, Peterson et al. 1995, 1997, 2007, Van den Borre & Watson 1997, 2000, Hilu & Alice 2001), muestran a Sporobolus como género más cercano a Calamovilfa (A. Gray) Hack. ex Scribn. & Southw., Crypsis Aiton, Fingerhuthia Nees, Spartina Schreb. y Urochondra C. E. Hubb. Estos seis géneros constituyen el "grupo Spartina" de Van den Borre & Watson (2000). Ahora bien, Calamovilfa, Crypsis, Sporobolus y Urochondra están ubicados en la subtribu Sporobolinae de Clayton & Renvoize (1986), junto con otros cuatro géneros (Hubbardochloa Auquier, Lycurus Kunth, Muhlenbergia Schreb. y Pereilema J. Presl). No obstante, la reciente circunscripción de la subtribu Sporobolinae (Columbus et al. 2007, Peterson et al. 2007) sólo considera a los géneros Calamovilfa, Crypsis, Spartina y Sporobolus. Así, la subtribu Sporobolinae está ubicada en la tribu Zoysieae (Columbus et al. 2007, Peterson et al. 2007). Véanse Peterson et al. (1995, 1997, 2004, 2007) para ampliar la información y la discusión taxonómica y filogenética en torno de estos géneros.
Origen del género Sporobolus
Según Roodt & Spies (2003), Sporobolus se originó probablemente de un linaje aneuploide de Eragrostis. Al respecto de la cercana relación entre estos dos géneros, Roodt & Spies (2003) destacan que Sporobolus puede representar un linaje especializado del género Eragrostis, con la diferenciación de tres números cromosómicos básicos (x = 6, 9, 10), en contraposición de un único número cromosómico básico paleopoliploide de Eragrostis (x = 10). Las especies de Sporobolus con un x = 6 representan especies antiguas del género, algunas probablemente extintas, mientras que las especies con un x = 9 son derivadas de paleopoliploides basadas en x = 10. Por lo tanto, los miembros de Sporobolus más recientes se han derivado después de iniciales eventos de poliploidización (Roodt & Spies 2003). Con relación a la cercanía entre Sporobolus y Eragrostis, Veldkamp (2002: 160) consideró que esta situación puede inferirse por la presencia del hongo tizón del género Bipolaris, el cual infecta únicamente las espiguillas de Sporobolus indicus (L.) R. Br. y de algunas especies de Eragrostis.
Cabe destacar que la principal diferencia morfológica entre los géneros Eragrostis y Sporobolus consiste en el mayor número de antecios por espiguilla en el primero. Al respecto, es necesario destacar la existencia de Eragrostis airoides Nees, una enigmática especie sudamericana (presente en Argentina, Bolivia, Brasil, Colombia, Paraguay, Uruguay, Venezuela y el Caribe), frecuentemente tratada como Sporobolus brasiliensis (Raddi) Hack., en virtud de su número cromosómico básico de x = 9, sus espiguillas paucifloras (1-3 antecios) y la morfología de su cariopsis, pero la posesión de espiguillas con no sólo un antecio sino frecuentemente con dos o tres y el modo de desarticulación de la espiguilla, son caracteres que respaldan la inclusión de S. brasiliensis en el género Eragrostis.
Distribución de las especies de Sporobolus en el noroeste de Sudamérica
Las once especies estudiadas presentan particularidades en cuanto a su distribución geográfica, ecológica y altitudinal, las cuales se detallan en la Tabla 1. Cabe destacar que de las especies registradas, diez se encuentran en Colombia, nueve en Perú, ocho en Venezuela y siete en Ecuador. Las especies, en su mayoría, crecen en hábitats abiertos de tierras bajas, y frecuentemente en hábitats ruderales e incluso urbanos.
Tratamiento taxonómico
Sporobolus R. Br., Prodr. 169. 1810. TIPO: Sporobolus indicus (L.) R. Br., Lectotipo designado por L. K. G. Pfeiffer, Nom. Bot. 2: 1274 (1874).
Agrosticula Raddi, Fl. Bras. Enum. Pl. 33. 1823. TIPO: Agrosticula muralis Raddi [= Sporobolus tenuissimus (Mart. ex Schrank) Kuntze].
Bennetia Raf., Bull. Bot. (Geneve) 1: 220. 1830. TIPO: Agrostis juncea Michx., hom. illeg. Heleochloa juncea P. Beauv. [= Sporobolus junceus (P. Beauv.) Kunth].
Triachyrum Hochst. ex A. Braun, Flora 24: 712. 1841. TIPO: Triachyrum adoense Hochst. ex A. Braun (= Sporobolus discosporus Nees).
Spermachiton Llanos, Fragm. Pl. Filip. 25. 1851. TIPO: Spermachiton involutum Llanos [= Sporobolus diandrus (Retz.) P. Beauv.].
Cryptostachys Steud.,Syn. Pl. Glumac. 1: 181. 1854. TIPO: Cryptostachys vaginata Steud., nom. nud. [= Sporobolus vaginiflorus (Torr. ex A. Gray) Alph. Wood].
Diachyrium Griseb., Abh. Konigl. Ges. Wiss. Gottingen 19: 257. 1874. TIPO: Diachyrium arundinaceum Griseb. [= Sporobolus rigens (Trin.) Desv.].
Bauchea E. Fourn., Mexic. Pl. 2: 87. 1886. TIPO: Bauchea karwinskyi E. Fourn. (= Sporobolus wrightii Munro ex Scribn.).
Plantas herbáceas, gráciles a robustas, anuales o perennes (en este caso pueden formar macollas muy densas y altas), cespitosas, rizomatosas, raramente estoloníferas. Cañas ramificadas (principalmente en los nudos inferiores) a más comúnmente simples, decumbentes a más frecuentemente erectas. Hojas gráciles a robustas, no auriculadas, basales (principalmente) y caulinares; vainas redondeadas o carinadas; lígulas una membrana muy reducida, ciliada, con tricomas cortos, rígidos; láminas aplanadas a convolutas, raramente cilíndricas en la parte superior y acanaladas hacia la base. Inflorescencias en panojas abiertas o contraídas a espiciformes, subincluidas en las vainas foliares superiores a más comúnmente exertas, con ramificaciones alternas o verticiladas, ramificaciones rematando en espiguilla, floríferas desde la base o sólo hacia su porción distal, en ocasiones las ramificaciones pueden ser glandulares. Espiguillas unifloras, sin aristas, glabras, subsésiles a pediceladas, comprimidas lateralmente o redondeadas; desarticulación por encima de las glumas o permaneciendo intactas hasta la expulsión del fruto, casmógamas, muy raramente cleistógamas; glumas 2, iguales o desiguales, múticas, glabras, hialinas, generalmente brillantes, a menudo de color verde oliva o grisáceas con matices metálicos, membranáceas a escariosas, uni-nervias o enervias, en ocasiones con nervaduras laterales apenas perceptibles; gluma inferior más corta que el antecio; gluma superior más corta o tan larga como el antecio, uni-nervia (muy raramente en algunas poblaciones colombianas de S. purpurascens la gluma superior es 2 ó 3-nervia); antecio perfecto; lema 1 (3)-nervia, membranácea, glabra, glumiforme, entera, aguda o acuminada, mútica; pálea 2-nervia, casi tan larga como la lema o poco mayor que ella, glabra, de textura y consistencia similar a la de la lema, entera, frecuentemente separándose (haciéndose bífida) entre los nervios cuando los frutos maduran; lodículas 2, uni-nervias, cuneadas, libres, glabras; estambres 3, raras veces 2; ovario glabro; estilos 2, libres; estigmas plumosos. Fruto un utrículo, pericarpio libre del antecio, café, sin surco ventral, plano en la cara ventral y redondeado en la dorsal, subgloboso, obovoide-elíptico a oblongo; embrión ca. ½ de la longitud del utrículo; hilo punctiforme.
Etimología. Del griego spora (semilla) y ballein (arrojar), en referencia a que sus semillas son expulsadas, por efecto de la humedad que disuelve el pericarpio (Nicora & Rúgolo de Agrasar 1987).
Número cromosómico básico. x = 5, 6, 9, 10, siendo más común un x = 9 ó 10 (Baaijens & Veldkamp 1991, Roodt & Spies 2003). En la Flora de China, Zhenlan & Phillips (2006) citan un x = 9, 12.
Tipo fotosintético. Todas las especies poseen el sistema de fotosíntesis Kranz (C4), con los subtipos bioquímicos NAD-ME y PCK (Ellis 1977, Baaijens & Veldkamp 1991, Simon & Jacobs 1999).
Observaciones. Sporobolus es confundido frecuentemente con Agrostis L. (Pooideae), ya que ambos géneros poseen espiguillas unifloras, pequeñas, con desarticulación por encima de las glumas e inflorescencias semejantes (panojas abiertas a contraídas). Es por esta razón que varias de las especies de Sporobolus fueron inicialmente descritas bajo el género Agrostis. Al respecto, Brown (1810) fue el que propuso el género Sporobolus para separar un conjunto de especies ubicadas en Agrostis, diferenciándolo por tener vainas pilosas, lígulas reducidas y semillas libres. No obstante, éstos pueden separarse fácilmente pues en Agrostis las glumas son más largas que el antecio (en Sporobolus la gluma inferior es más corta que el antecio y la superior más corta o tan larga como éste), la lema es 3-5-nervia (uni-nervia en Sporobolus), el fruto es una cariopsis con surco ventral (mientras que en Sporobolus es un utrículo sin surco ventral) y la lígula es una membrana (correspondiendo a una hilera de tricomas cortos en Sporobolus). Por otra parte, en Agrostis la pálea está generalmente suprimida o representada por un minúsculo rudimento (siempre presente en Sporobolus) y además, frecuentemente se encuentran especies con lema aristada o denticulada, mientras que en Sporobolus ésta es siempre mútica.
En cuanto a la historia taxonómica de Sporobolus, podemos destacar que a partir de la creación del género por Brown (1810), sólo Stapf (1898) retomó su estudio y fue el primero en dividirlo en dos secciones: Sporobolus sect. Chaetorhacia Stapf y Sporobolus sect. Eusporobolus Stapf. Posteriormente, Pilger (1956) dividió la última sección, elevándola a la categoría de subgénero [Sporobolus subgen. Sporobolus (Stapf) Pilg.], en seis grupos, los que basó en el hábito y en las características de las panojas y las glumas. Basándose en la morfología del fruto, Bor (1960) dividió el género en cinco grupos no naturales. Más recientemente, Baaijens & Veldkamp (1991) dividieron a Sporobolus subgen. Sporobolus en cinco secciones, las que basaron en la morfología integral con especial referencia al sistema de ramificación de la inflorescencia. Por otra parte, en su estudio de las especies de Sporobolus del nordeste de México, Peterson et al. (2004) reconocieron tres "grupos" taxonómicos, los que delimitaron con base en caracteres morfológicos y anatómicos. No obstante, aún no hay una división satisfactoria del género Sporobolus (véase Ortiz-Díaz & Culham 2000).
Clave para separar las especies de Sporobolus presentes en el noroeste de Sudamérica
1. Plantas anuales.
2. Inflorescencia abierta, oblonga a piramidal.
3. Gluma superior de 0,2-0,5 mm de largo; espiguillas de 0,7-1,1 mm de largo.
10. Sporobolus tenuissimus
3'. Gluma superior de 1,2-1,8 mm de largo; espiguillas de 1,0-1,8 mm de largo.
9. Sporobolus pyramidatus
2'. Inflorescencia comprimida-espiciforme. 7. Sporobolus pilifer
1'. Plantas perennes.
4. Vainas de las hojas viejas adheridas a las plantas, las cuales se disponen en forma de numerosos rizos o rotas a manera de numerosas fibras; espiguillas pajizas. 6. Sporobolus mirabilis
4'. Vainas de las hojas viejas caducas o siempre enteras cuando permanecen adheridas a las plantas.
5. Inflorescencia abierta, piramidal.
6. Gluma inferior 0,3-0,8 mm de largo. 9. Sporobolus pyramidatus
6'. Gluma inferior 1,5-2,5 mm de largo.
7. Espiguillas pajizas. 2. Sporobolus cubensis
7'. Espiguillas oscuras con matices metálicos.
8. Vainas densa, corta y suavemente pilosas. 5. Sporobolus lasiophyllus
8'. Vainas glabras y/o ciliadas. 1. Sporobolus bogotensis
5'. Inflorescencia comprimida-espiciforme o con las ramificaciones levemente divergentes.
9. Plantas fuertemente rizomatosas. 11. Sporobolus virginicus
9'. Plantas sin rizomas.
10. Gluma inferior 1,6-2,5 mm de largo. 8. Sporobolus purpurascens
10'. Gluma inferior 0,2-1 mm de largo.
11. Gluma superior entera, aguda a obtusa; espiguillas de 1,9-2,7 mm de largo; inflorescencia comprimida-espiciforme. 3. Sporobolus indicus
11'. Gluma superior erosa a denticulada, truncada; espiguillas de 1,3-2,0 mm de largo, inflorescencia con las ramificaciones que divergen del eje principal, en ocasiones levemente plegadas al eje principal. 4. Sporobolus jacquemontii
Descripción de las especies
1. Sporobolus bogotensis Swallen & García-Barr., Caldasia 2 (8): 301. 1943. TIPO: Colombia. Cundinamarca: Macizo de Bogotá, Sierra de Siete Picos, 2900-3100 m, 28 jun 1939, J. Cuatrecasas 5680 (holotipo US-1795919!, isotipo COL!). Fig. 2 A-B.
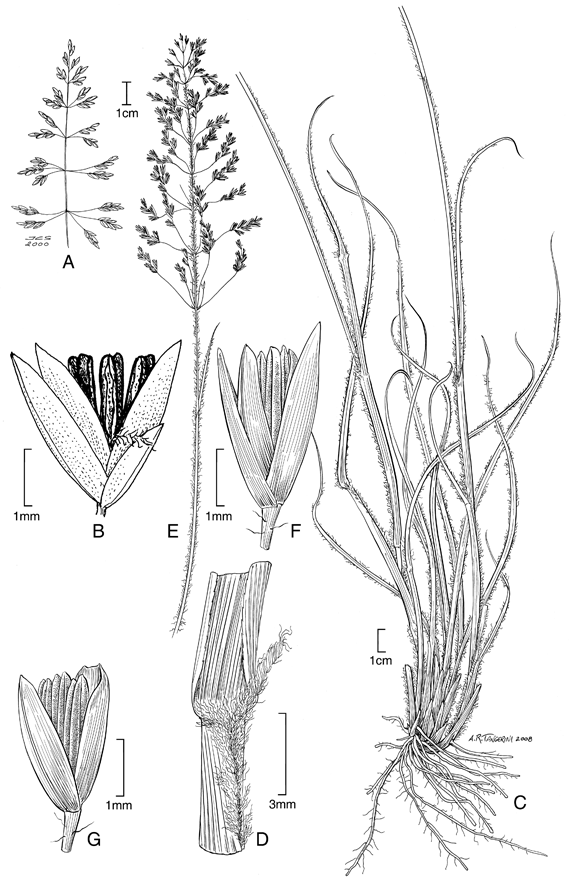
Plantas perennes, cespitosas, generalmente gráciles. Cañas erectas, simples, hasta de 0,8 m de altura. Hojas principalmente basales; vainas redondeadas, glabras aunque con las márgenes ciliadas, imbricadas; lígulas densamente ciliadas, 0,2-0,3 mm de largo; láminas lineares, involutas, abaxialmente glabras o glabrescentes y estriadas, adaxialmente cortamente híspidas, 5-35 cm de largo, 0,2-0,4 cm lat. Inflorescencias en panojas piramidales, 7-20 cm de largo, con ramificaciones verticiladas o subverticiladas, éstas últimas sólo floríferas desde la mitad hasta su porción distal. Espiguillas 2,6-3,5 mm de largo, verdes oscuras a café oscuras, brillantes con matices metálicos, cortamente pediceladas, pedicelos glabros, 0,5-1 mm de largo; gluma inferior 1,5-2(-2.3) mm de largo, lanceolada a aguda, uninervia o enervia; gluma superior de igual longitud que la espiguilla, ovada, aguda, uninervia, lisa, brillante; lema glumiforme; pálea ovada, aguda o obtusa, un poco más corta que la lema; estambres 3, anteras 1,5-2 mm de largo. Fruto 1,5-2 mm de largo, elíptico, lateralmente comprimido.
Distribución geográfica y ecológica. Sporobolus bogotensis es endémica de áreas altoandinas de Colombia, Ecuador y Perú (Lægaard & Peterson 2001, Peterson et al. 2001), y se distribuye en áreas de fuertes pendientes con arbustales y/o matorrales, entre los 2000 y 3300 m de altitud.
Material examinado
COLOMBIA. Boyacá: Mun. Villa de Leyva, saliendo por el camino que conduce a Iguaque, en las afueras del casco urbano de Villa de Leyva, 2200 m, 18 dic 2004, D. Giraldo-Cañas et al. 3830 (COL). Mun. Villa de Leyva, ca. 2000 m, año 1862, M. Lindig 1084 (COL). Mun. Villa de Leyva, cerca de Sáchica, ca. 2000 m, ago 1964, C. Saravia 4081 (COL). Cundinamarca: Ubaté, 100 km N of Bogotá, 2600 m, 18 may 1952, M. Køie 4519 (US). Santander: Mule path to Bolívar, 10 km NW of Vélez, 22 abr 1944, N. Fassett 25097 (US).
ECUADOR. Azuay: South of Oña, slope above Tablón de Oña, 2740 m, 21 feb 1945, F. Fosberg 23197 (US). Loja: Entre Rancho Ovejero y Saraguro, 3020 m, 3 ago 1959, H. Barclay & P. Juajibioy 8543 (US). S of Loja, E of Universidad Nacional Ills in Argelia, 2350 m, 21 feb 1987, I. Grignon 84271 (US). Mountain range W of Loja, behind Barrio de La Reina del Cisne, along old road to Catamayo, 2300-2400 m, 22 feb 1987, I. Grignon 84279 (US).
PERÚ. Cajamarca: Prov. Chota, 7 km N of Chota on road towards Conchán, 2570 m, 19 mar 2000, P. Peterson & N. Refulio Rodríguez 14970 (US).
2. Sporobolus cubensis Hitchc., Contr. U.S. Natl. Herb. 12 (6): 237. 1909. TIPO: Cuba. La Habana: Isle of Pines, 6 mar 1904, A. H. Curtiss 392 (holotipo US!). Fig. 3 A-B.
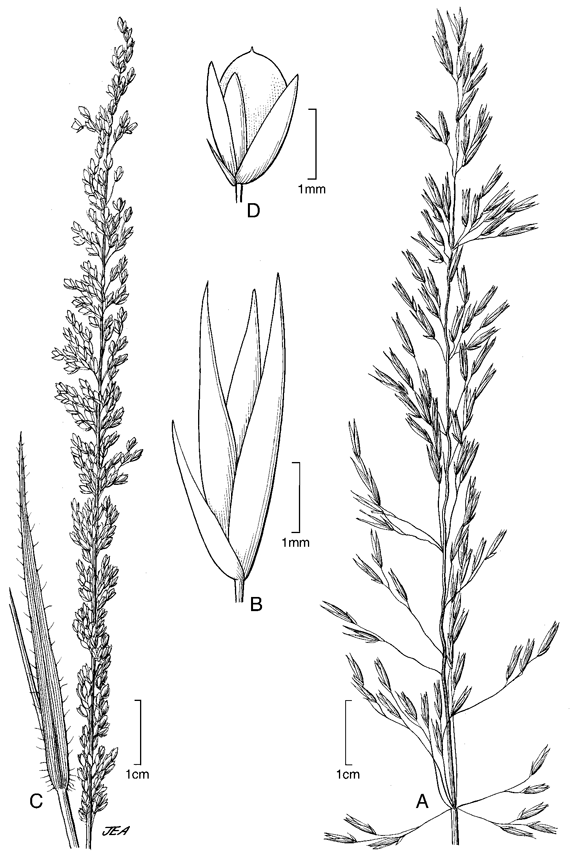
Plantas perennes, fuertemente cespitosas, con fascículos vigorosos fuertemente arraigados al suelo. Cañas erectas, simples, hasta de 0,70 m de altura. Hojas principlamente basales; vainas redondeadas, glabras a glabrescentes, márgenes densamente ciliadas; lígulas 0,1-0,3 mm de largo, ciliadas; láminas aplanadas a convolutas, acuminadas, glabras a glabrescentes, márgenes ciliadas principalmente en sus porciones inferiores, 15-57 cm de largo, 0,3-0,4 cm lat. Inflorescencias en panojas abiertas, piramidales, 8-18 cm de largo, con las ramificaciones verticiladas y floríferas en sus porciones distales. Espiguillas 3,2-4,1 mm de largo, pajizas; gluma inferior 1,6-2,1 mm de largo, aguda, uninervia, el nervio apenas perceptible; gluma superior 2,7-3,1 mm de largo, aguda, uninervia; lema 2,8-3,9 mm de largo, más angosta que la gluma superior, inconspicuamente nervada; pálea tan larga como la lema; estambres 3, anteras 2,6-2,8 mm de largo. Fruto 1,8-2,1 mm de largo, elíptico, redondeado en el ápice.
Distribución geográfica y ecológica. Esta especie se distribuye desde México y el Caribe hasta Bolivia y Brasil. Sporobolus cubensis se encuentra normalmente en sabanas (principalmente en sabanas no inundables) así como en los límites entre esteros y morichales, y menos frecuentemente en matas de monte de tierras bajas, entre el nivel del mar y los 900 (1300) m de altitud. Regularmente se le encuentra en sabanas sujetas a quemas periódicas.
Nombre vulgar. Pasto de sabana (H. García-Barriga 4981).
Observaciones. Sporobolus cubensis es una especie poco variable en su morfología y se caracteriza, principalmente, por sus vainas de márgenes densamente ciliadas y por sus inflorescencias piramidales con ramificaciones verticiladas y grandes espiguillas. Los ejemplares de herbario referidos a esta especie, generalmente presentan señales de fuego en sus bases, lo que según de Castro Boechat & Longhi-Wagner (1995) estaría indicando que esta especie es una de las primeras en rebrotar y florecer después de las quemas en los sitios en que crece.
Material examinado
COLOMBIA. Caquetá: Mun. San Vicente del Caguán, sabanas del Yarí, hato Canadá, vereda El Recreo, 300 m, 27 ene 1990, J. Betancur & M. Velásquez 1550 (COL, HUA), 1551 (COAH, COL, HUA). Guaviare: Mun. San José del Guaviare, sitio Los Pozos Naturales, en sabanas de suelos arenosos, ca. 262 m, 27 mar 1997, R. López & O. Rodríguez 2419 (COAH, COL). Meta: Altillanura ondulada con morichales, estero de un morichal, al sur del río Casibare, 8 may 1963, Blydenstein & Saravia 872 (COL). En sabanas, sin localidad, 300 m, feb 1937, H. García-Barriga 4981 (COL, US). Carimagua, ca. 300 m, 2 abr 1972, J. Hilton 8 (COL). Mun. Puerto Gaitán, entre Carimagua y El Porvenir, jun 1985, B. Rivera 2 (COL). Carimagua, grassland, 200 m, 18 sep 1992, S. Renvoize 5405 (COL, US). Reserva Natural La Macarena, northern bank of the Río Güéjar, near the mouth of Río Zanza, savannas east of El Tablazo, Cuchilla del Neme, 21 ene 1968, J. Thomas et al. 1490 (COL). Llanos de San Martín, 200 m, ene 1856, J. Triana 125 (COL, US). Vichada: Altillanura plana, cerca del hato Carigene, al este de Orocué, 14 nov 1963, Blydenstein 1798 (COL). Gaviotas, Cumaribo, 24 mar 1973, I. Cabrera 2819 (COAH). Mun. Puerto Carreño, 3 km al oriente de El Tapón, ca. 200 m, 11 abr 1995, M. Córdoba et al. 1452 (COL). Mun. Puerto Carreño, vía a Puente Nuevo, 13 abr 1995, M. Córdoba et al. 1508 (COAH). Llanos along the Río Vichada, 3 km SE of Masaguaro, ca. 27 km NE of San José de Ocuné, 100 m, 20 ene 1944, F. Hermann 11012 (COL, US). Carretera entre Puerto Carreño y Puerto Gaitán, fundo El Delirio, bajo Rompe-Cajas, ca. 150 m, 21 mar 1971, P. Pinto & C. Sastre 1466 (COL).
VENEZUELA. Anzoátegui: Vicinity of San Tomé, 264 m, 22 mar 1940, A. Chase 12484 (US). Sabanas anegadas, San Tomé, 26 ene 1944, H. Pittier 15172 (US). Aragua: High savannas of Guayabito, 15 mar 1926, H. Pittier 12133 (US). Bolívar: SW of La Vergarena, S of Ciudad Bolívar, 3 dic 1953, J. de Alba 53-25 (US). La Gran Sabana, abr 1939, Cristophel s.n. (US). La Gran Sabana, km 287 along road El Dorado-Santa Elena, 24 km N of Santa Elena, 830 m, 5 dic 1973, G. Davidse et al. 4913 (COL). Near El Paujil, Río Cabas, 800-900 m, 3 nov 1985, R. Liesner 19440 (MO, US). La Gran Sabana, between Kun and Uaduara-parú, in valley of Río Kuknán, 1065-1220 m, 1 oct 1944, J. Steyermark 59046 (US). Mérida: Between Mucuchachí and Canaguá, 6 may 1944, J. Steyermark 56325 (US). Monagas: Jusepin along road to Manresa, 220-250 m, 30 mar 1967, R. Pursell et al. 8603 (US). Portuguesa: Llanos de Araure, 9 abr 1925, H. Pittier 11747 (US). Sucre: Valley of Cocollar, 820 m, 28 abr 1945, J. Steyermark 62389 (US). Brasil-Venezuela. Roraima: rodovia BR-174 à 7 km de Santa Helena, 1000 m, 19 jun 1974, J. Murça Pires et al. 14568 (US).
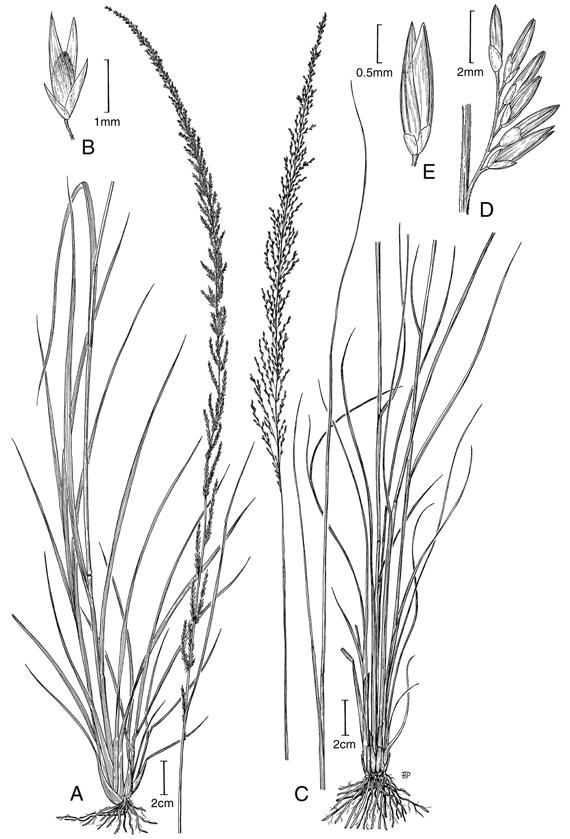
3. Sporobolus indicus (L.) R. Br., Prodr. 170. 1810. Agrostis indica L., Sp. Pl. 63. 1753. TIPO: Estados Unidos de América. Virginia: Clayton 460B [lectotipo LINN-84.36!, designado por Hubbard, Agron. Lus. 28. 1967; isolectotipo SI! (fragm.)]. Fig. 4 A-B.
Vilfa berteroana Trin., Mem. Acad. Imp. Sci. Saint-Petersbourg, Ser. 6, Sci. Math., Seconde Pt. Sci. Nat. 6,4 (1-2): 100. 1840. Sporobolus berteroanus (Trin.) Hitchc. & Chase, Contr. U.S. Natl. Herb. 18 (7): 370. 1917. TIPO: Santo Domingo, Bertero s.n. (holotipo LE-TRIN-1682.01!, isotipos B, MO-2095245!).
Plantas perennes, cespitosas, frecuentemente densamente cespitosas, fuertemente arraigadas al suelo. Cañas erectas, simples, hasta de 1 m de altura, aunque pueden en algunas ocasiones alcanzar portes de 1,3 m de altura. Hojas principalmente basales; vainas redondeadas a levemente carinadas, estriadas, glabras, ciliadas; lígulas diminutas, ciliadas, 0,1-0,3 mm de largo; láminas lineares, aplanadas, acuminadas, terminando en una punta muy fina y flexuosa, 5-35 (-50) cm de largo, 0,3-0,5 cm lat., cara abaxial glabra, cara adaxial ligeramente escabrosa. Inflorescencias en panojas densas, contraídas a espiciformes, lanceoladas a lineares, erectas, (6-) 10-35 cm de largo, con ramificaciones cortas y adpresas al eje principal, las ramificaciones floríferas desde su base. Espiguillas subsésiles a cortamente pediceladas, adpresas, grisáceas, 1,9-2,7 mm de largo; gluma inferior 0,4-1 mm de largo, obtusa a truncada, levemente erosa, enervia; gluma superior 0,7-1,6 mm de largo, ovada, aguda a obtusa o redondeada en el ápice, frecuentemente erosa, generalmente ½ a 3/4 la longitud de la espiguilla, uninervia o enervia; lema 2-2,6 mm de largo, glumiforme, ovada, aguda, generalmente uninervia, aunque en ocasiones puede presentar tres nervios débiles; pálea tan larga como la lema o levemente más corta, de igual consistencia a la de la lema; estambres 3, anteras 0,5-1 mm de largo. Fruto 0,7-1,3 mm de largo, obovado, lateralmente comprimido e incluso cuadrangular, redondeado a truncado en el ápice.
Distribución geográfica y ecológica. Sporobolus indicus presenta una amplia distribución latitudinal y altitudinal, conociéndose en el continente americano desde el sur de los Estados Unidos de América y el Caribe hasta Argentina y Chile. Ésta crece en una variada gama de ambientes abiertos como suelos arenosos o rocosos, orillas de caminos, bancos de arena y especialmente en áreas ruderales y alteradas así como urbanas, entre el nivel del mar y los 2700 m de altitud, aunque raramente puede alcanzar altitudes de 3600 (3900) m. Cabe destacar que Rúgolo de Agrasar & Puglia (2004) consideran a esta especie nativa de América templada-cálida y naturalizada en zonas cálidas de todo el mundo, Clayton (1965) la considera restringida al Nuevo Mundo, Baaijens & Veldkamp (1991) la consideran cosmopolita, mientras que Peterson et al. (2003) la reconocen como pantropical.
Nombres vulgares. Colombia: Espartillo (H. García-Barriga 6411), grama (S. L. Amaya 45), guayacán (E. Pérez-Arbeláez 268), rabo de burro (L. C. Wiedemam 5, F. Crespo 55), raboeburro (F. Crespo 55), rabo de mula (A. Rangel-Galindo 93); Ecuador: Purún, sinchiquigua (M. Acosta-Solís 9375); Perú: Mula tsampa (P. Hutchinson & J. Wright 4285); Venezuela: Según Luces (1942), a S. indicus se le conoce en Venezuela con los nombres de paja de gallina, tucupén y tucupera.
Usos. En Boyacá se emplea para la elaboración de artesanías diversas (F. Crespo 55). Por otra parte, se emplea para aliviar dolencias del corazón (S. L. Amaya 45), como alimento de cabras, vacas, ovejas y caballos (A. Rangel-Galindo 93). Sporobolus indicus se considera una forrajera de mediano valor, pero tiene la ventaja de que es muy resistente al pisoteo (Rúgolo de Agrasar & Puglia 2004). No obstante, según Pohl et al. (1994: 273) las plantas pueden ser tóxicas para el ganado, cuando éstas están infectadas (las inflorescencias) por el hongo tizón [Bipolaris ravenelii (Curt.) Subram. & Jain]. Por otra parte, Rúgolo de Agrasar & Puglia (2004) le confieren interés ornamental, aunque adicionan que esta especie merece ser revalorizada en los ambientes donde crece espontáneamente. Luces (1942) comenta que S. indicus es una de las gramíneas más apetecidas por el ganado.
Número cromosómico. 2n = 18, 24, 36 (Peterson et al. 2003, 2004).
Observaciones. A raíz de su amplia distribución geográfica y altitudinal, S. indicus es una especie altamente polimorfa; así, las plantas de tierras intermedias (800-1700 m) son más robustas y sus panojas son más largas. Adicionalmente, las anteras exhiben una mayor longitud a medida que se asciende en el gradiente altitudinal. No obstante, el tamaño de las espiguillas y los frutos es constante, independientemente de la altura sobre el nivel del mar.
Por otra parte, cabe mencionar que en los ejemplares de herbario correspondientes a S. indicus, frecuentemente se determinan como S. poiretii (Roem. & Schult.) Hitchc., pero este binomio es un sinónimo de S. junceus (P. Beauv.) Kunth (Peterson et al. 2001), una especie exclusiva de México y los Estados Unidos de América. Veánse los comentarios dados para S. jacquemontii. En el ejemplar J. Soukup 5474 (US) había mezcla de dos panojas de S. indicus y dos panojas de Eragrostis sp.
Material examinado
COLOMBIA. Antioquia: Rionegro (Río Negro), 2100 m, 22 jun 1930, W. Archer 285 (US). Medellín, Santa Elena, 1500-2000 m, 1 ene 1931, W. Archer 1223 (US). Mun. Jardín, sitio La Truchería, carretera Jardín-Riosucio, km 5, 1900-2000 m, 5 ene 2003, D. Giraldo-Cañas et al. 3431 (COL). Mun. Envigado, loma de El Escobero, ca. 2 km antes de la intersección con la carretera Las Palmas, ca. 1850 m, 26 dic 2003, D. Giraldo-Cañas et al. 3620 (COL). Mun. Jardín, cerro Amarillo, 7 km de Jardín, 2100 m, R. Pohl & M. Palacio 15584 (HUA, MO). Mun. Urrao, La Nevera, río Las Juntas, 2450 m, R. Pohl & J. Betancur 15475 (HUA, MO). Mun. Frontino, km 15 of road Nutibara-Murrí, 1840 m, J. Zarucchi et al. 5770 (HUA, MO). Bogotá D. C.: Campus de la Universidad Nacional de Colombia, en las jardineras internas del Edificio Sindú, 2600 m, 28 abr 2009, D. Giraldo-Cañas 4207 (COL). Sabana de Bogotá, 2600 m, sin fecha, E. Pérez-Arbeláez 268 (COL). Boyacá: Mun. Paipa, vereda Caños, 2517 m, 23 sep 1984, S. L. Amaya 45 (COL). Mun. Iza, en inmediaciones de Los Termales, 2500 m, 22 jun 2003, D. Giraldo-Cañas et al. 3431 (COL). Mun. Villa de Leyva, camino entre la hacienda "Torcoroma de Arriba" y el cañón de Las Clusias, ca. 2500 m, 18 jul 2003, D. Giraldo-Cañas et al. 3544 (COL). Mun. Villa de Leyva, saliendo por el camino que conduce a Iguaque, en las afueras del casco urbano de Villa de Leyva, 2200 m, 18 dic 2004, D. Giraldo-Cañas et al. 3827 (COL). Mun. Villa de Leyva, valle de Saquencipá, 2200 m, mar 1941, A. Rangel-Galindo 93 (COL). Mun. Moniquirá, vereda El Ajizal, 1700 m, 25 ene 2008, F. Crespo 55 (COL). Caldas: Alrededores de Manizales, carretera a Fresno, 2280 m, 6 jul 1959, P. Pinto 446 (COL). Caquetá: Mun. Florencia, carretera Florencia-Neiva (Huila), puente Cerro Negro, 1950 m, 1-5 jul 2005, D. Giraldo-Cañas et al. 3929 (CAUP, COL). Cauca: Mun. Popayán, predios internos y jardineras abandonadas de la Facultad de Educación de la Universidad del Cauca, 1750 m, 7-11 nov 2004, D. Giraldo-Cañas et al. 3812 (COL, HUA). Cundinamarca: Sabana de Bogotá, 2600 m, 29 dic 1939, J. Cuatrecasas 489 (US). Mun. Albán, cordillera Oriental andina, vereda La María, Granjas del Padre Luna, carretera Bogotá-Honda, 1800-2000 m, 21 abr 2001, D. Giraldo-Cañas et al. 3222 (COL). Provincia de Río Negro, Mun. Pacho, carretera Pacho-La Capilla, alrededores del estadio municipal de fútbol, ca. 1600 m, 13 ene 2004, D. Giraldo-Cañas et al. 3734 (COL). Mun. Susa, cerro Morroplancho, carretera Susa-Ubaté, a unos 2 km del casco urbano de Susa, ca. 2700 m, 13 mar 2004, D. Giraldo-Cañas et al. 3738 (COL). Huila: Mun. Suaza, carretera Suaza-Florencia (Caquetá), en borde de carretera, ca. 1900 m, 1-5 jul 2005, D. Giraldo-Cañas et al. 3927 (COL). La Guajira: Alrededores de Uribia, 40 m, 28 feb 1962, C. Saravia 133, 284 (COL). Magdalena: Alrededores de San Andrés, 1300 m, 20 ene 1959, R. Romero Castañeda 6987 (COL). Nariño: Mun. Chachagüí, barrancos de la entrada principal al aeropuerto "Antonio Nariño", ca. 1700 m, 24 abr 2009, D. Giraldo-Cañas 4206 (COL). Mun. Genoy, carretera Genoy-Pasto, laderas del volcán Galeras, en áreas de canteras húmedas abandonadas (explotación de arenas y rocas) al pie de la carretera, ca. 2700 m, 22 abr 2009, D. Giraldo-Cañas 4197 (COL). Mun. La Unión, cerro La Jacoba, 2070 m, 31 jul 1977, P. Pinto et al. 1736 (COL). Putumayo: Valle de Sibundoy, Sibundoy, 2200 m, 7 may 1963, M. Bristol 931 (COL, US). Quindío: Mun. Filandia, camino principal de la reserva Bremen, 1500 m, 20-25 may 2005, D. Giraldo-Cañas 3883 (COL). Santander: Mun. Charalá, hacienda La Palma, 1300 m, 26 ene 1942, L. C. Wiedemam 5 (COL). Valle del Cauca: Entre El Cerrito y Palmira, El Trejo, 1050 m, 28 dic 1938, H. García-Barriga 6411 (COL).
ECUADOR. Chimborazo: Cañón of Río Chanchán, near Huigra, 7-14 may 1945, W. Camp E-3098 (US). 4.5 km W of Río Bamba on road to Guamote, 2900 m, 24 may 1990, P. Peterson & E. Judziewicz 9305 (US). 3.5 km SW of Alausí on road to Huigra, 2330 m, 29 may 1990, P. Peterson & E. Judziewicz 9322 (US). Cotopaxi: 6.2 km N of Latacunga on the Pan American Highway, 2900 m, 14 abr 1990, P. Peterson et al. 8749 (US). 6.8 km W of Pilalo on road to El Tingo, 2000 m, 15 abr 1990, P. Peterson et al. 8753 (US). 20 km N of Ambato and 21.5 km S of Latacunga, just off the Pan American highway, 2745 m, 16 abr 1990, P. Peterson et al. 8788 (US). Imbabura: Yaguarcocha, 2200 m, 15 sep 1950, M. Acosta-Solís 19038 (US). Loja: 9 km NE of Celica on road to Guachanama, 2500 m, 3 jun 1990, P. Peterson & E. Judziewicz 9509 (US). Pastaza: At Río Pastaza, at km 5 on road Shell-Palora, 1000 m, 3 dic 1984, S. Lægaard 53438 (COL). Pichincha: Calderón, N of Quito, 2450 m, 6 mar 1984, S. Lægaard 51698 (COL). Km 43,4 Quito-Tabacundo at junction of road to Cochasqui, 2500 m, 18 mar 1988, S. Lægaard & S. Renvoize 70806 (COL). 13 km N of Calderón on the Pan American Highway, 2300 m, 10 may 1990, P. Peterson et al. 9065 (US). Tungurahua: Entre Cevallos y Quero, 2700-2900 m, 30 dic 1944, M. Acosta-Solís 9834 (US). West of Banos where the bridge crosses the Río Chambo, 2050 m, 16 abr 1990, P. Peterson et al. 8779 (US). Alrededores de Ambato, 2600 m, 14 dic 1944, M. Acosta-Solís 9375 (US).
PERÚ. Amazonas: Prov. Chachapoyas, 1 km SW of Chachapoyas, 2300 m, 22 may 1962, J. Wurdack 464 (US). Ancash: Prov. Carhuas, Cordillera Blanca, valley of Río Marcará, Vicos, 2960 m, 10 mar 1964, P. Hutchinson & J. Wright 4285 (US). Prov. Recuay, Cordillera Blanca, 16 km NE of Catac on road towards Chavin de Huantar, 3630 m, 21 mar 1997, P. Peterson & N. Refulio Rodríguez 13829 (US). Ayacucho: Parinacochas, along arroyo near small río above (north) of Laguna Parinacochas, 3300 m, 1 mar 2002, P. Peterson et al. 16327 (US). Prov. Huamanga, S of Ayacucho ca. 23 km on Hwy 3 towards Abancay, 3575 m, 15 mar 2007, P. Peterson et al. 20500 (US). Ayacucho, sin datos, 2407 m, 7 feb 1968, J. Soukup 5474 (US, en este ejemplar había mezcla de dos panojas de S. indicus y dos panojas de Eragrostis sp.). Cajamarca: Prov. Cajamarca, 12 km N of Cajamarca on road towards Bambamarca, 3020 m, 14 mar 2000, P. Peterson & N. Refulio Rodríguez 14862 (US). Prov. Cutervo, 13 km W of Cutervo on road towards Socoto, 2160 m, 20 mar 2000, P. Peterson & N. Refulio Rodríguez 15013 (US). Prov. San Ignacio, 5 km NW of San José de Lourdes on road towards La Sidra, 1370 m, 28 mar 2000, P. Peterson & N. Refulio Rodríguez 15076 (US). 2 km E of La Cruce de Tabaconas on road towards Tamborapa, 1700 m, 31 mar 2000, P. Peterson & N. Refulio Rodríguez 15119 (US). Cuzco: Machu-Pichu, camino entre la ciudad inca de Machu-Pichu y el Puente del Inca, ca. 2000 m, 18 sep 2004, D. Giraldo-Cañas 3763 (COL). Prov. Calca, Pisac, 3000 m, mar 1944, F. Marín 105 (US). Prov. Cuzco, entre Socorro y Kimko, 3560 m, 15 mar 1960, C. Vargas 13147 (US). Huánuco: Prov. Huánuco, Chinchao, entre Huánuco y Tingo María, 200-2100 m, 9 jul 1958, R. Ferreyra 13108 (US). Prov. Pachitea, ENE of Huanuco ca. 21 air km, 13 km ESE of Puerto Rancho jct. Hwy 3, near S side of bridge over Río Huallaga on road to Panao, 2498 m, 6 mar 2007, P. Peterson et al. 20344 (US). La Libertad: Prov. Santiago de Chuco, 22 km E of Huamachuco on road towards Sarin above Río Chusgón, 2500 m, 29 mar 1997, P. Peterson & N. Refulio Rodríguez 13976 (US). Puno: Prov. El Collao, 10 km NW of Pomata on road towards Ilave, 3900 m, 4 mar 1999, P. Peterson et al. 14624 (US). San Martín: Divisoria, 59 km from Tingo María on highway to Pucallpa, 1540 m, 17 nov 1949-15 ene 1950, H. Allard 21287 (US).
VENEZUELA. Distrito Federal: Caracas, 1500 m, A. Fendler 1682 (US). Lara: Near Sanare, parque Yacambu, 1400 m, 16 feb 1967, R. Smith V-1361 (US). Mérida: Entre Mucurubá y Mucuchíes, 20 sep 1942, T. Lasser 364 (US), 385 (US). Miranda: Alrededores del Centro de Ecología, Altos de Pipe, 1670 m, 22 abr 1994, L. O. Marulanda 1594 (HUA). Tabay, 1800-2000 m, 17 sep 1930, W. Gehriger 452 (US). Monagas: Hillside ca. 14 km SW of Caripe, 930 m, 23 jun 1967, R. Pursellet al. 9382 (US). Trujillo: Sabanas de Altamira, serranía La Cristalina, 1950 m, 28 ago 1941, F. Tamayo 1836 (US).
4. Sporobolus jacquemontii Kunth, Revis. Gramin. 2: 427, t. 127. 1831. Vilfa jacquemontii (Kunth) Trin., Mém. Acad. Imp. Sci. Saint.Pétersbourg, Sér. 6, Sci. Math., Seconde Pt. Sci. Nat. 6,4 (1-2): 92. 1840. Sporobolus pyramidalis P. Beauv. var. jacquemontii (Kunth) Jovet & Guédès, Taxon 22: 163. 1973. TIPO: Crescit in insula St. Domingo, 1827, Jacquemont s.n. (holotipo P, isotipo LE-TRIN-1712.01!). Fig. 4 C-E.
Plantas perennes, cespitosas, fuertemente arraigadas al suelo. Cañas erectas, simples, hasta de 1,2 m de altura. Hojas principalmente basales; vainas redondeadas, glabras, ligeramente ciliadas, estriadas; lígulas 0,1-0,3 mm de largo, ciliadas; láminas aplanadas a convolutas, lineares y largamente atenuadas, terminando en una punta muy fina y flexuosa, abaxialmente glabras, adaxialmente piloso-híspidas, márgenes escabrosas, 10-40 (-70) cm de largo, 0,2-0,5 cm lat. Inflorescencias en panojas oblongas o lineares, erectas, exertas, 12-30 (-35) cm de largo, con ramificaciones levemente divergentes del eje principal, alternas y floríferas desde su base. Espiguillas 1,3-2 mm de largo, grisáceas, lanceoladas, subsésiles a cortamente pediceladas; gluma inferior 0,2-0,8 mm de largo, hialina, enervia, oblonga, erosa o redondeada a obtusa; gluma superior 0,5-1 mm de largo, 1/4 a 1/3 del largo de la espiguilla, uninervia, largamente oblonga, redondeada o truncada en el ápice, a menudo erosa a denticulada; lema ovado-elíptica, uninervia, raramente 3-nervia, caso en el que los nervios son apenas perceptibles, aguda, del largo de la espiguilla o ligeramente más corta que ésta; pálea igual o ligeramente más corta que la lema y similar a ella; estambres 3, anteras 0,8-1,1 mm de largo.Fruto obovoide-elíptico a oblongo, lateralmente comprimido, truncado en el ápice, 0,8-1,1 mm de largo
Distribución geográfica y ecológica. Sporobolus jacquemontii se distribuye desde el sur de los Estados Unidos de América y el Caribe hasta Bolivia y Brasil, así como en África y Australia. Según Clayton (1965) y Peterson et al. (2004), S. jacquemontii es nativa del continente americano. Ésta crece en una variada gama de ambientes abiertos, aunque es básicamente una especie ruderal y se le encuentra entre el nivel del mar y los 1500 (1850) m. Esta especie es remplazada ecológicamente por S. indicus en alturas mayores (Pohl et al. 1994, y observaciones personales).
Nombres vulgares. Colombia: Castilleda (G. Bonet 37), cutupen (J. Idrobo 5219), espartillo (J. Idrobo 4358), guayacán (E. Pérez-Arbeláez 337); Venezuela: tucupene (L. Williams 13310).
Uso. Forraje (fuente: especímenes C. Chardon 123, J. Idrobo 5219 y H. Pittier 14484).
Número cromosómico. 2n = 24 (Pohl et al. 1994, Peterson et al. 2003, 2004).
Observaciones. Sporobolus jacquemontii es frecuentemente confundida con S. indicus, pues además de presentar un hábito y una morfología similares, comparten con frecuencia ciertos hábitats, principalmente ruderales. No obstante, éstas pueden separarse fácilmente ya que la gluma superior en S. indicus es aguda a obtusa, mientras que en S. jacquemontii es erosa a denticulada y truncada; por otra parte, en S. indicus las espiguillas son más grandes (1,9-2,7 mm de largo) que en S. jacquemontii, en la que las mismas alcanzan dimensiones un poco menores (1,3-2,0 mm de largo). Además, en S. indicus las inflorescencias se muestran contraidas, mientras que en S. jacquemontii las ramificaciones de las inflorescencias divergen del eje principal de las mismas (en ocasiones levemente plegadas al eje principal), correspondiendo en S. jacquemontii, en la mayoría de los casos, con panojas piramidales.
Material examinado
COLOMBIA. Antioquia: Mun. Medellín, 1500 m, 3 jul 1930, W. Archer 361 (COL, US). Mun. Arboletes, orilla de playa, 30 sep 1986, J. Betancur et al. 308, 314 (COL, HUA). Mun. San Jerónimo, km 10 of road San Jerónimo-Medellín, 1225 m, 19 sep 1987, J. Zarucchi et al. 5404 (COL, HUA, MO). Arauca: Mun. Arauca, inmediaciones del campus de la Universidad Nacional de Colombia, hacienda El Cairo, carretera Arauca-Tame, km 9, 200-300 m, 13 jun 2003, D. Giraldo-Cañas et al. 3489, 3504 (COL). Bolívar: Cartagena, Mamonal, 30 oct 1963, G. Bonet 37 (COL). Carretera Sincelejo-Tolú, 30 m, 3 jun 1950, H. García-Barriga 13473 (COL, US). Caldas: Río Magdalena, estación Pita, 200 m, 3 mar 1961, J. Idrobo 4358 (COL). Caquetá: Mun. Florencia, Barrio Villa Natalia, en un campo pedregoso en estado de sucesión temprana y dominado por gramíneas, ca. 280 m, 27-30 jun 2005, D. Giraldo-Cañas et al. 3942, 3947 (COAH, COL). Casanare: Mun. El Yopal, en áreas urbanas del centro de la ciudad, 400 m, 31 oct 2007, D. Giraldo-Cañas 4130 (COAH, COL). Llanos orientales, 15 km al sur de El Yopal, cerca del río Charte, 3 abr 1963, J. Blydenstein 700 (COL). Carretera a El Yopal, cerca del río Cherté, ca. 400 m, 19 oct 1962, J. Idrobo 5219 (COL). Chocó: Mun. Quibdó, en la ribera derecha del río Atrato, en inmediaciones del Malecón y la Plaza de Mercado, 90 m, 6 nov 2005, D. Giraldo-Cañas 3971, 3976 (CAUP, COL, HUA). Córdoba: 5 km carretera San Benito-San Juan, 35 m, 17 jul 1973, J. Idrobo & A. Cleef 6632 (COL). Cundinamarca: Mun. Nilo, hacienda La Guaira, inmediaciones del río Pagüey, 350 m, 16 oct 2004, D. Giraldo-Cañas et al. 3779 (COL).Provincia Gualiva, Mun. Villeta, Cordillera Oriental andina, vertiente occidental, ca. 900 m, 17-19 jun 2006, D. Giraldo-Cañas et al. 4081 (COL). Mun. Nariño, ca. 500 m, jul 1930, E. Pérez-Arbeláez 337 (COL, US). Guaviare: Mun. San José del Guaviare, trocha Nuevo Tolima, en cercanías del Batallón José Joaquín París, 250 m, mar 1996, D. Giraldo-Cañas & R. López 2568 (COAH, COL). Mun. San José del Guavire, Inspección La Fuga, 200 m, mar 1996, D. Giraldo-Cañas & R. López 2632 (COAH). La Guajira: Alrededores de Uribia, 40 m, 28 feb 1962, C. Saravia 212-b (COL). Meta: Mun. Villavicencio, estación biológica «Roberto Franco», 500 m, 25 ene 1986, J. Fernández et al. 4890 (COL). Quindío: Mun. Armenia, jardineras de la Avenida del Museo Quimbaya, 1500 m, 20-26 mar 2005, D. Giraldo-Cañas & J. García-Ulloa 3871 (COL, HUA). Vía Armenia-Barcelona, 1250 m, 3 may 2000, C. Agudelo et al. 4171 (COL). San Andrés, Providencia y Santa Catalina: Isla de San Andrés, cerca de la fábrica de grasa de coco El Cove, 26 jul 1966, J. Torres et al. 405 (COL). Santander: Along edge of Quebrada Honda, near Estación Las Palmas, 500-800 m, 6 jul 1953, J. Langenheim 3245 (COL, US). Tolima: Cordillera Central andina, Mun. Ibagué, jardineras y andenes de la Plaza de Bolívar, ca. 1200 m, 11-12 jun 2005, D. Giraldo-Cañas 3906 (COL, HUA). Valle del Cauca: Plana del Valle, bordes de la carretera entre Palmira y Cali, ca. 1000 m, 25 abr 1963, López Filgueiras 8463 (US). Mun. Palmira, predios de la Facultad de Agronomía, Universidad Nacional de Colombia, ca. 1000 m, 15 ene 1964, López Filgueiras 8499 (COL, US). Vichada: Mun. Puerto Carreño, afloramientos rocosos entre Punta de Lajas y el cerro El Bita, ribera del río Orinoco, 40-100 m, 4-5 ene 2004, D. Giraldo-Cañas & C. Parra 3657 (COL).
ECUADOR. Esmeraldas: San Lorenzo, around houses, 0-10 m, 22 ene 1987, I. Grignon 84204 (US). Los Ríos: 2 km S of Babahoyo, 300 m, 5 may 1990, P. Peterson & C. Annable 9036 (US). 2 km S of Babahoyo on road to Guayaquil near Inmetal Ceramientos Corporation Yard, 200 m, 5 may 1990, P. Peterson & C. Annable 9043 (US). Pichincha: 13 km E of Alluriquín (32 km E of Santo Domingo) on road to Quito, 980 m, 5 jun 1990, P. Peterson & E. Judziewicz 9534 (US).
PERÚ. Loreto: Prov. Coronel Portillo, cerca de Neshuya, entre Pucallpa y Tingo María, 250 m, 16 dic 1968, R. Ferreyra 17581 (US).
VENEZUELA. Amazonas: Depto. Atures, 8 km of Puerto Ayacucho, 85 m, 13-15 abr 1978, G. Davidse & O. Huber 15050 (MO, US). Cerca de la desembocadura del río Sanariapo, 120 m, 23 may 1940, L. Williams 13080 (US). Anzoátegui: Las Cañas, río Caris, en las sabanas del bajo, 12 sep 1940, H. Pittier 14484 (US). Apure: Llanos del Alto Apure, sabanas de Hato Matapalo, 150 m, 7 may 1911, A. Jahn 195-A (US). Barinas: La Yuca, cercanías de Barinas, abr 1956, M. Ramia 1070 (US). Bolívar: Sabana de El Tigre, cerca del río Cuchivero, 100 m, 13 jun 1940, L. Williams 13310 (US). Carabobo: Vicinity of Valencia, 400-800 m, 22 jun 1920, H. Pittier 8887 (US). Cojedes: San Carlos, 19 jun 1940, C. Chardon 123 (US). Distrito Federal: Ingomar (Turmerito), 950 m, jun 1936, H. Pittier 13684 (US), 13694 (US). Guarico: At intersection of Río Orituco and road from Calabozo to Cazorla, ca. 90 m, 4 nov 1973, G. Davidse 3718 (COL, MO). Monagas: Río Guarapiche, ca. 9 km WSW of Jusepin, 160 m, 25 may 1967, R. Pursell et al. 9020 (US). Sucre: Dtto. Sucre, Parque Nacional Mochima, pueblo de Mochima, 9 sep 1982, B. Garófalo et al. 1078 (US). Táchira: La Mulera, 1250 m, 24 sep 1942, F. Tamayo 2303 (US). Trujillo: Llanos de Monay, 610 m, 28 may 1944, J. Steyermark 56777 (US).
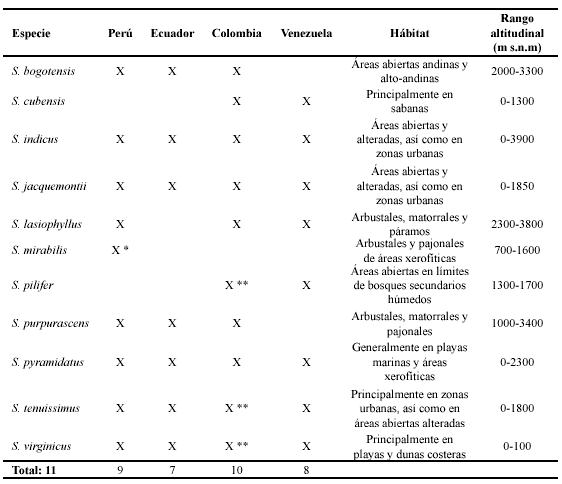
5. Sporobolus lasiophyllus Pilg., Bot. Jahrb. Syst. 37: 504. 1906. TIPO: Perú. Junín: In montibus ad oppidum Tarma, in eadem formatione, 3600 m, feb 1903, Weberbauer 2381 (lectotipo US-1061515!, aquí designado). Fig. 2 C-G.
Plantas perennes, cespitosas, generalmente de porte grácil. Cañas erectas, hasta de 0,4 m de altura, simples, raramente ramificadas. Hojas principalmente basales, follaje muy denso ya que los entrenudos son extremadamente cortos; vainas densa, corta y suavemente pilosas, anchas; lígulas ciliadas, 0,5-0,7 mm de largo; láminas planas a frecuentemente plegadas, glabras a glabrescentes, escabriúsculas marginalmente, pilosas cerca de la región ligular, 2-18 cm de largo, 0,15-0,35 cm lat. Inflorescencias en panojas abiertas, piramidales, laxas, conspicuamente exertas, 4-14 (-20) cm long, con ramificaciones verticiladas, filiformes, hasta de 5 cm de largo, floríferas desde su porción medial. Espiguillas 3,1-3,8 mm de largo, verde oscuras a negruzcas, con matices metálicos, cortamente pediceladas, los pedicelos 1-1,5 mm de largo; gluma inferior 2-3 mm de largo, aguda; gluma superior tan larga como la espiguilla o subigual, aguda; lema tan larga como la gluma superior o subigual, de ápice agudo; pálea como la lema; estambres 3, anteras 1,5-2 mm de largo.Fruto obovoide-elíptico a oblongo, lateralmente comprimido, ca. 1,5 mm de largo
Distribución geográfica y ecológica. Sporobolus lasiophyllus es una especie endémica de la región andina de Perú, Colombia y Venezuela. Se le encuentra frecuentemente en matorrales y arbustales altoandinos así como en subpáramos y páramos de zonas principalmente secas, y del mismo modo, es frecuente en áreas de pendientes pronunciadas y suelos arenosos, entre los 2300 y los 3800 m de altitud. Posiblemente se le encuentre también en Ecuador, pero hasta el momento no hemos detectado ejemplares de ese país.
Observaciones. Esta especie es fácilmente reconocible por sus hojas basales con vainas densamente pilosas y por sus inflorescencias piramidales laxas con espiguillas verde-negruzcas con matices metálicos. Por otra parte, hay que destacar que en ocasiones las cañas muestran alternancia de macroblastos y braquiblastos, naciendo, a partir de los nudos de estos últimos, hojas de vainas muy apretadas, con algunas raíces desarrolladas en la base del respectivo nudo. Esta situación la observamos en los ejemplares Peterson et al. 21870, 21879 (US), recolectados en Perú.
Material examinado
COLOMBIA. Antioquia: Un kilómetro al norte de Santa Rosa de Osos, 2600 m, 25 sep 1948, J. Araque M. & F. Barkley 18A003 (MEDEL, US). Bogotá D. C.: Bogotá, cerro Guadalupe, 2900 m, nov 1911, B. Apollinaire & B. Arthur 59 (US). Bogotá, cerro de Monserrate, 3100 m, 8 oct 1944, N. Fassett 25861 (COL, US). Macizo de Bogotá, cerro entre la quebrada Las Delicias y la quebrada Las Ninfas, páramo, 3000-3180 m, 25 jun 1939, J. Cuatrecasas 5634 (US). Bogotá, Vitelma, 2800 m, 4 jun 1944, F. Hermann 11359 (US). Bogotá, páramos en la vía a La Calera, 30 may 1926, G. Woronow & S. Juzepczuk 5117 (US). Boyacá: Mun. Tota, laguna de Tota, sitio Playa Blanca, carretera Tota-Aquitania, 3000-3100 m, 22 jun 2003, D. Giraldo-Cañas et al. 3534 (COL). Carretera Socha-Los Pinos, km 41, 2km al N del Alto Las Pilas y 4 km al ENE de Socha, 2970 m, 22 may 1973, A. Cleef 9878 (COL, US). Mun. Aquitania, borde del cerro El Órgano, lago de Tota, 3025 m, 13 may 1976, O. Rangel & J. Aguirre 341 (COL). Cundinamarca: Mun. Sesquilé, vereda Las Espigas, finca La Esperanza, matorrales altoandinos, ca. 2900 m, 24 ago 2003, D. Giraldo-Cañas et al. 3588 (COL). Mun. Soacha, cerros de La Chucúita, 2650 m, sep 1940, H. García-Barriga 10308 (HUA). Región de la laguna de La Herrera, 2600-2800 m, 3 may 1985, V. Wijninga & R. Vink 162 (COL). Santander: Páramo de Berlín, carretera Pamplona-Bucaramanga, 6 km al SW de Berlín, 3320 m, 13 jun 1973, A. Cleef 10188 (COL). Páramo de Los Puentes, above La Baja, 3500-3700 m, 25 ene 1927, E. Killip & A. Smith 18236 (US). Páramo de Berlín, 3650 m, 8 ago 1968, L. Mora 4545 (COL). Departamento desconocido, J. C. Mutis 5495 (US).
PERÚ. Ancash: Prov. Bolognesi, cerros al SE de Chiquián, 3680 m, 28 abr 1952, E. Cerrate 1394 (US). Prov. Recuay, Cordillera Blanca, 16 km NE of Catac on road towards Chavin de Huantar, 3630 m, 21 mar 1997, P. Peterson & N. Refulio Rodríguez 13831 (US). Prov. Sihaus, 31 km N of Pomabamba on road towards Sihaus, 3780 m, 24 mar 1997, P. Peterson & N. Refulio Rodríguez 13904 (US). Prov. Pallasca, 17 km N of Huandoval and 6 km S of Pallasca, 3360 m, 27 mar 1997, P. Peterson & N. Refulio Rodríguez 13924 (US). Ayacucho: Prov. Huamanga, S of Ayacucho ca. 23 km on Hwy 3 towards Abancay, 3575 m, 15 mar 2007, P. Peterson et al. 20493 (US). Cajamarca: 6 km S of Cajabamba on road towards Huamachuco, 2780 m, 30 mar 1997, P. Peterson & N. Refulio Rodríguez 13999 (US). Prov. Cajamarca, 1 km S of Huambocancha on road towards Cajamarca, 2770 m, 14 mar 2000, P. Peterson & N. Refulio Rodríguez 14848 (US). Prov. Cajabamba, WSW of Cajabamba ca. 23 air km, SW of Araqueda ca. 9 air km, along road branching south away from road towards Ventana (in La Libertad), 3207 m, 24 mar 2008, P. Peterson & R. Soreng 21870 (US). Cajamarca to Celendin, 3 km NE of Encanada, 3200 m, 22 mar 1988, S. Renvoize & S. Lægaard 4850 (US). Piura: Prov. Huancabamba, 14 km E of Sondor on road towards Tabaconas, 2540 m, 31 mar 2000, P. Peterson & N. Refulio Rodríguez 15157 (US).
VENEZUELA. Mérida: Páramo de Don Pedro, 3000 m, 10 jul 1944, V. Badillo 999 (US). Páramo de La Negra, between Bailadores and La Grita, 2745-3045 m, 7 jul 1944, J. Steyermark 57074 (US). Trujillo: Alrededores de La Morita, arriba de Jajó, 2800 m, ago 1958, L. Aristeguieta 3507 (US).
6. Sporobolus mirabilis Pilg., Bot. Jahrb. Syst. 56 (Beibl. 123): 27. 1920. TIPO: Perú. Cajamarca: Prov. Jaen, zwischen dem Tale des Shumba, eins rechten Nebenflusses des Chinchipe und der Ortschaft Jaen, grassteppe mit Staucheren, 700-800 m, sehr Haufig, stellenweise reine Bestande bildend, Apr 1912, Weberbauer 6186 [holotipo B, isotipos BAA-2910 fragm. ex B!, US-81956!]. Fig. 5 A-F.
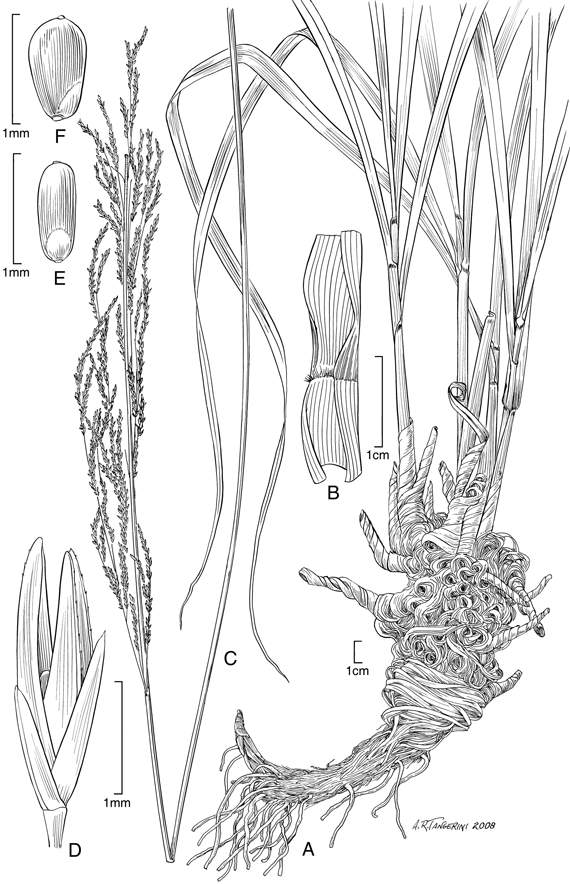
Plantas perennes, robustas, rizomatosas, los rizomas cortos y vigorosos. Cañas erectas, hasta de un metro de altura. Hojas principalmente basales, numerosas; vainas glabras, ciliadas hacia la porción ligular, pajizas a amarillentas, las vainas de las hojas viejas adheridas a la base de las plantas a manera de rizos numerosos o rotas a manera de numerosas fibras; lígulas densamente ciliadas, 0,5-0,7 mm de largo; láminas aplanadas, glabras, de ápice casi setiforme, hasta de 90 cm de largo, 0,6-0,9 cm lat. Inflorescencias en panojas angostas, densas, exertas, 27-35 cm de largo, ramificaciones alternas, de orden consecutivo, delicadas, las primarias hasta de 20 cm de largo, las secundarias hasta de 4 cm de largo.Espiguillas 2,2-2,8 mm de largo, pajizas, cortamente pediceladas, pedicelos 0,4-0,8 mm de largo; glumas desiguales, más cortas que la espiguilla; gluma inferior 1,1-1,4 mm de largo, aguda; gluma superior 1,9-2 mm de largo, aguda, uni-nervia; lema 2,2-2,8 mm de largo, lanceolada, aguda, escabrosa; pálea como la lema; estambres 2, anteras ca. 1 mm de largo. Fruto no visto.
Distribución geográfica y ecológica. Sporobolus mirabilis es una especie endémica de Perú, la cual sólo se conoce de Cajamarca. Ésta crece en áreas de pajonales en una matriz de arbustales así como en áreas xerofíticas de Cactaceae y Acacias, entre los 700 y los 1600 m de altitud. Al parecer, esta especie es poco frecuente dada la escasez de colecciones.
Observaciones. Sporobolus mirabilis es fácilmente reconocible por su hábito robusto, sus grandes inflorescencias y por sobre todo, por las vainas de las hojas viejas que permanecen adheridas a las plantas, las cuales se disponen en forma de numerosos rizos o rotas a manera de numerosas fibras, lo que le da a ésta un aspecto único entre las especies del género.
Material examinado
PERÚ. Cajamarca: 10 km from Balsas towards Celendin, 1550 m, 25 mar 1988, S. Renvoize & S. Lægaard 4941 (US).
7. Sporobolus pilifer (Trin.) Kunth, Enum. Pl. 1: 211. 1833. Vilfa pilifera Trin., Gram. Unifl. Sesquifl. 157. 1824. TIPO: Nepal, Wallich 3774 (lectotipo LE, designado por Baaijens & Veldkamp, Blumea 35: 441. 1991; isolectotipos BM, K, US-81956!). Fig. 3 C-D.
Sporobolus ciliatus J. Presl, Reliq. Haenk. 1 (4,5): 242. 1830. TIPO: Panamá, Hab. in Panama, Haenke s.n. (holotipo ?, isotipo MO!).
Sporobolus rupestris Kunth, Revis. Gramin. 1 : 267, t. 45. 1830. TIPO: Brasil, in humidis petrosis pr. Friburgo, 1836, a cl. Beyrich l. dr. ill. a Roemer (holotipo ?, isotipo LE-TRIN-1737.01!).
Plantas anuales, gráciles, cespitosas. Cañas erectas, hasta de 30 cm de altura, simples o ramificadas. Hojas gráciles, principalmente basales; vainas carinadas, glabras o laxamente pilosas, las márgenes ciliadas; lígulas 0,2-0,3 mm de largo, ciliadas; láminas aplanadas a convolutas, de base cordiforme, laxamente pilosas en ambas caras, ciliadas, 1,3-6 cm de largo, 0,2-0,4 cm lat. Inflorescencias en panojas espiciformes, erectas, exertas, delgadas a gráciles, 2-9 cm de largo, con las ramificaciones verticiladas en los nudos inferiores, floríferas desde su base, glandulares. Espiguillas 1,8-2 mm de largo, pardas; gluma inferior 0,6-0,7 mm de largo, aguda, frecuentemente enervia, ocasionalmente uninervia; gluma superior 1,6-1,7 mm de largo, aguda, uninervia, un poco más ancha que la gluma inferior; lema 1,7-1,9 mm de largo, aguda, glumiforme; pálea aguda, como la lema; estambres 3, anteras 0,5-0,7 mm de largo. Fruto 1-1,3 mm de largo, aplanado, redondeado en el ápice.
Distribución geográfica y ecológica. Esta especie se distribuye desde Nicaragua y Costa Rica hasta Venezuela, las Guayanas y Brasil, así como en Asia y África. No obstante, aquí se registra por primera vez en territorio colombiano. En Colombia sólo la hemos registrado en los departamentos de Huila y Norte de Santander, en caminos sombreados de un bosque secundario húmedo así como en colinas áridas, entre los 1300 y los 1700 m de altitud.
Número cromosómico. 2n = 36, 40 (Zhenlan & Phillips 2006), 54 (Pohl et al. 1994).
Observaciones. Sporobolus pilifer es frecuentemente escrito en las determinaciones de material de herbario como Sporobolus piliferus, y aún esta escritura aparece en diversas publicaciones botánicas.
Material examinado
COLOMBIA. Huila: Mun. San Agustín, parque arqueológico San Agustín, en caminos sombreados, ca. 1700 m, 1-5 jul 2005, D. Giraldo-Cañas et al. 3919 (COL, HUA, US). Norte de Santander: East of Ocaña, arid clay hills, Cordillera Oriental, 1300 m, 16 dic 1943, F. Hermann 10897 (US, dos ejemplares). Departamento desconocido, año 1842, I. Linden 1565 (US).
VENEZUELA. Miranda: Sebastopul, 17 oct 1938, V. Badillo 123 (US).
8. Sporobolus purpurascens (Sw.) Ham., Prodr. Pl. Ind. Occid. 5. 1825. Agrostis purpurascens Sw., Prodr. 25. 1788. TIPO: Jamaica, Swartz s.n. (holotipo S!, isotipo BM). Figs. 1 A-B y Figs. 6 A-D.

Plantas perennes, cespitosas. Cañas erectas, ocasionalmente geniculadas, raramente ramificadas en los nudos inferiores, hasta de 0,75 m de altura. Hojas principalmente basales; vainas redondeadas, estriadas, glabras, frecuentemente ciliadas; lígulas 0,5-1 mm de largo, ciliadas; láminas aplanadas a plegadas, atenuadas, abaxialmente glabras, usualmente ciliadas a ciliado-pectinadas, especialmente en la base, 4-25 cm de largo, 0,2-0,5 cm lat. Inflorescencias en panojas exertas, contraídas, cilíndricas a espiciformes, 6-20 cm de largo, con ramificaciones en verticilos bien definidos, las ramificaciones floríferas desde su base. Espiguillas cortamente pediceladas, purpúreas a café oscuras o verde oscuras, brillantes con matices metálicos, 2,7-4,1 mm de largo; gluma inferior 1,6-2,5 mm de largo, lanceolada, aguda, uninervia o enervia; gluma superior 2,7-4,1 mm de largo, ovada, aguda a acuminada, uninervia, raramente 2-3-nervia, los nervios con asperezas dispuestas con su punta hacia la región distal; lema 2,6-3,9 mm de largo, glumiforme, uninervia; pálea (2,3-) 2,5-3,8 mm de largo, glumiforme; estambres 3, anteras 0,9-1,2 mm de largo. Fruto 1,6-2,1 mm de largo, obovado a elíptico, lateralmente comprimido, redondeado en el ápice, rugoso.
Distribución geográfica y ecológica. Esta especie se distribuye desde el sur de los Estados Unidos de América y el Caribe hasta Perú y Brasil. Sporobolus purpurascens crece en ambientes de pajonales y matorrales de sustratos arenosos o rocosos, principalmente de áreas semiáridas o subxerofíticas así como en páramos, y en ocasiones se le encuentra como especie ruderal, entre los 1000 y los 3400 m de altitud.
Número cromosómico. 2n = 60 (Pohl et al. 1994, Peterson et al. 2003, 2004).
Observaciones. Sporobolus purpurascens es muy similar a S. bogotensis, aunque difieren básicamente en la forma de la panoja, la cual es estrechamente cilíndrica-espiciforme y con ramificaciones provistas de espiguillas desde su porción basal en S. purpurascens, mientras que en S. bogotensis la panoja es ovoide a piramidal con las ramificaciones sólo floríferas en su mitad distal. Adicionalmente, las láminas de S. purpurascens tienen las márgenes basales conspicuamente ciliadas, siendo glabras o escasamente pilosas en S. bogotensis. Es interesante destacar que varios especímenes de varias localidades altoandinas de Bogotá y Cundinamarca (Colombia) [Giraldo-Cañas 3245, 3256, Idrobo et al. 419, McKee 10521, Triana 58], presentan la gluma superior 2 ó 3-nervia, característica que no se ha hallado en los demás ejemplares del continente americano, los que poseen gluma superior uni-nervia. Adicionalmente, estos ejemplares con gluma superior 2 ó 3-nervia poseen las panojas más densas y más comprimidas entre todos los representantes de la especie. En el ejemplar H. García-Barriga 10923 (COL) hay mezcla de individuos pertenecientes a S. purpurascens y a Agrostis sp.
Material examinado
COLOMBIA. Bogotá D. C.: Localidad de Usme, cuenca alta del río Tunjuelito, carretera Usme-La Base-El Hato, desvío a Truchas El Danubio, en matorrales altoandinos, 3200-3300 m, 23 sep 2001, D. Giraldo-Cañas & C. Acosta 3245, 3256 (CAUP, COL, HUA, US). Cordillera de Bogotá, 3000 m, 1 nov 1856, J. Triana 58 (COL, US). Localidad de Usme, al sur de Usme, entre La Regadera y El Hato, estación agrícola experimental Usme, 3000-3100 m, 27 jun 1950, J. Idrobo et al. 419 (COL, US). Boyacá: Mun. Villa de Leyva, camino entre la hacienda "Torcoroma de Arriba" y el cañón de Las Clusias, matorrales degradados, ca. 2500 m, 18 jun 2003, D. Giraldo-Cañas et al. 3548 (COL). Desierto 4 km W de Villa de Leyva, 2200 m, 26 dic 1971, A. Cleef 340 (US). Mun. Sáchica, 1990 m, ago 1964, C. Saravia 4300, 4364 (COL). Duitama, 10 dic 1929, Toro 44 (US). Cauca: Entre Popayán y Río Blanco, 1780-1850 m, 6 ene 1947, J. Cuatrecasas 23409 (US). El Tambo, 1745 m, 7 may 1935, H. García-Barriga 4423 (COL, US). El Ramal to Río Sucio, W of Popayán, 1600-1900 m, 3 jul 1922, F. Pennell & E. Killip 8152 (US). Mun. Popayán, alrededores de la ciudad, vía al oriente, 1800 m, 18 oct 1999, B. Ramírez 12392 (CAUP, COL). Mun. Popayán, sector nortede la ciudad, 1800 m, 27 ago 2000, B. Ramírez 13458 (CAUP, COL). De Balboa a Argelia, 15,5 km al E de Balboa, ca. 3000 m, 27 jun 1989, F. Zuloaga & X. Londoño 4193 (COL, MO, US). Tierradentro, Aguacate, 1600 m, año 1983, J. Wood 3923 (COL). Around Tacueyó, Río Palo Valley, Huila group, Central Cordillera, 1800 m, ene 1906, H. Pittier 1024 (US). Cundinamarca: Mun. Mosquera, laguna La Herrera, 2680 m, 14 dic 1943, H. García-Barriga 10923 (COL, US), 10925 (COL, US). Road to Páramo de Guasca, 3100 m, 23 jul 1963, H. McKee 10521 (US). Ubaté, 2700 m, año 1983, J. Wood 3502 (COL). Huila: Mun. San Agustín, parque arqueológico San Agustín, en sitios abiertos y bordes de caminos, ca. 1700 m, 1-5 jul 2005, D. Giraldo-Cañas et al. 3917 (COL, US). Nariño: Carretera Túquerres-Pasto, cerca del río Juanambú, 1380-1820 m, 1-5 dic 1962, C. Saravia & R. Jaramillo 1930 (COL). Norte de Santander: Páramo de Fontibón, 2700 m, 21 jul 1940, J. Cuatrecasas & H. García-Barriga 10098 (COL). Valle del Cauca: San Antonio above Cali, road to Buenaventura, Western Cordillera, 1900 m, dic 1906, H. Pittier 746 (US). Mun. Yumbo, entre Pubenza y Lulalo, 1100 m, 21 nov 1962, C. Saravia 1737 (COL). Departamento desconocido, Lehmann BT-1030 (US).
ECUADOR. Azuay: Gualaceo, on road to Remigio Crespo, 2400 m, 29 dic 1986, I. Grignon 84103 (US). 5.6 km S of La Paz on the Pan American Highway, and 36.3 km N of Ona, 3000 m, 24 abr 1990, P. Peterson et al. 8891 (US). 7 km S of Chordeley on road to Sigsig, 2220 m, 28 abr 1990, P. Peterson et al. 8943 (US). Carchi: North of Juncal, 2600 m, 18 ene 1987, I. Grignon 84193 (US). Imbabura: Angochagua, 2800 m, 20 nov 1949, M. Acosta Solís 4576 (US). Loja: Alturas orientales de Loja, 2250 m, 2 may 1946, R. Espinosa 275 (US). Mountain range W of Loja, behind Barrio de La Reina del Cisne, along road to Catamayo, 2300-2400 m, 22 feb 1987, I. Grignon 84278 (US). Between La Toma and Loja, 1800-2600 m, 4 sep 1923, A. Hitchcock 21418 (US). Between Loja and San Lucas, 2100-2600 m, 6 sep 1923, A. Hitchcock 21463 (US). 10 km S of Loja on road to Zumba, then 1-2 km E on road to Parque Nacional Podocarpus, 2500 m, 31 may 1990, P. Peterson & E. Judziewicz 9384 (US). 8 km S of Vilcabamba on road to Zumba, 1870 m, 31 may 1990, P. Peterson & E. Judziewicz 9397 (US). 4 km E of Purinuma on road to Malacatos, 2300 m, 1 jun 1990, P. Peterson & E. Judziewicz 9453 (US). Pichincha: Alturas de Quito, 2800-3000 m, 15 dic 1945, M. Acosta Solís 11331 (US). Tumbaco, 2400 m, 29 may 1939, E. Asplund 6563 (US). Zamora: Zamora-Huaico, E de Loja, Cordillera Oriental, 2600-2800 m, 2 abr 1951, M. Acosta Solís 20503 (US).
PERÚ. Cajamarca: Prov. Cutervo, 1 km W of Cutervo on road towards Socota, 2460 m, 20 mar 2000, P. Peterson & N. Refulio Rodríguez 14998 (US). Prov. Cutervo, Dist. San Andrés, a 3 km al NE de la localidad de San Andrés, 2250 m, 18 jun 1980, I. Sánchez Vega 2320 (US). San Martín: San Roque, 1350-1500 m, ene-feb 1930, L. Williams 7533 (US). Piura: Prov. Huancabamba, 14 km E of Sondor on road towards Tabaconas, 2540 m, 31 mar 2000, P. Peterson & N. Refulio Rodríguez 15152 (US).
9. Sporobolus pyramidatus (Lam.) Hitchc., Man. Grasses W. Ind. 84. 1936. Agrostis pyramidata Lam., Tabl. Encycl. 1 : 161. 1791. TIPO: West Indies, Martinique, J. Richard s.n. (holotipo P, isotipo LE-TRIN-1645.01, fragm. et illust.!). Fig. 7 A-B.
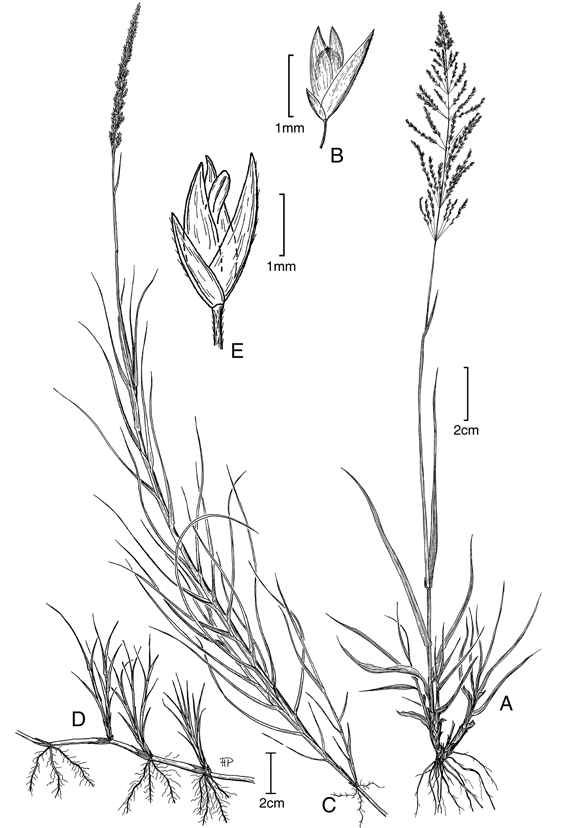
Vilfa arguta Nees, Fl. Bras. Enum. Pl. 2: 395. 1829. Sporobolus argutus (Nees) Kunth, Enum. Pl. 1: 215. 1833. TIPO: Brasil, F. Sellow s.n. (sintipo LE-TRIN-1676.01!).
Plantas anuales o perennes, cespitosas. Cañas erectas a decumbentes, ocasionalmente ramificadas, hasta de 0,65 m de altura. Hojas principalmente basales; vainas redondeadas, estriadas, glabras, ciliadas; lígulas 0,3-0,7 mm de largo, ciliadas; láminas aplanadas a convolutas, abaxialmente glabras a glabrescentes, adaxialmente pilosas, ciliadas en sus porciones inferiores, 4-20 cm de largo, 0,2-0,6 cm lat. Inflorescencias en panojas contraídas cuando inmaduras a piramidales en la madurez, exertas, 4-20 cm de largo, con las ramificaciones verticiladas a subverticiladas y frecuentemente con glándulas conspicuas, las ramificaciones distales floríferas en toda su extensión, las basales con espiguillas en su mitad superior. Espiguillas pediceladas, 1,2-1,7 (-2) mm de largo, pardo-amarillentas, grisáceas o argénteas, lateralmente comprimidas; gluma inferior subulada, lanceolada, obtusa a aguda, enervia o uninervia, 0,3-0,8 mm de largo, más corta que la ½ ó 1/3 de la espiguilla; gluma superior ovada a lanceolada, agudo-acuminada, uninervia, tan larga como la espiguilla o ligeramente más corta que ésta; lema glumiforme, tan larga como la gluma superior o ligeramente menor, ovada, lanceolada, aguda, uninervia; pálea hialina, como la lema y tan larga como ésta o levemente más corta; estambres 3, anteras 0,2-0,5 mm de largo. Fruto elipsoide a oblongo, lateralmente comprimido, redondeado en el ápice, 0,6-1 mm de largo, finamente estriado.
Distribución geográfica y ecológica. Esta especie es nativa del continente americano (Peterson et al. 2003) y se distribuye desde los Estados Unidos de América y el Caribe hasta Argentina y Chile. Sporobolus pyramidatus crece en áreas alteradas, en caminos, zonas ruderales, áreas pendientes de tierras bajas (aunque raramente puede alcanzar ambientes abiertos a alturas que no sobrepasan los 2300 m), aunque principalmente se le encuentra en playas marinas y en ambientes xerofíticos.
Nombre vulgar. Teatino (R. Echeverri 927, 1077).
Número cromosómico. 2n = 24, 36, 54 (Peterson et al. 2003, 2004).
Material examinado
COLOMBIA. Atlántico: Palmar de Varela, abr 1939, A. Dugand 1231 (US). Ribera del río Magdalena, Las Flores, 30 oct 1967, A. Dugand 7106 (COL). Puerto Colombia, Sabanilla, 21 nov 1912, A. Hitchcock 9937 (US). Puerto Colombia, 10 m, 6 jul 1963, H. McKee 10438 (COL, US). Bolívar: Cartagena, 20 nov 1912, A. Hitchcock 9894, 9915, 9918 (US). La Guajira: Mun. Uribia, corregimiento Nazareth, Playa Nueva York, y laguna de Los Patos-Puerto Estrella, 0-5 m, 29 dic 2004, J. Betancur et al. 11228 (COL). Mun. Uribia, corregimiento Nazareth, Playa Nueva York, 0-5 m, 6 ene 2005, J. Betancur et al. 11367 (COL). Pájaro, 10 m, 28 nov 1959, J. Cuatrecasas 25467 (US). Cerrejón, 200 m, 14 abr 1949, O. Haught 6587 (US). Wind swept plains of Riohacha, 1914, J. Sinclair s.n. (US-865576). Cabo de La Vela, ca. 5 m, 28 mar 1964, C. Saravia & M. de Saravia 3500 (COL). Magdalena: Near Fonseca, 130 m, 1 oct 1944, O. Haught 4381 (US). Santa Marta, parque nacional natural Tayrona, Playa Brava, 0-3 m, dic 1991, J. Martínez et al. 16 (COL). Región de Santa Marta, Ciénaga, en Aguacoca, 20 m, 16 feb 1950, R. Romero Castañeda 1960 (COL). Ciénaga, alrededores de Aguacoca, 20 m, 21 feb 1959, R. Romero Castañeda 7260 (COL, US). Isla de Salamanca, cerca del Riñón de Majagualito, ca. 5 m, 14 sep 1966, M. Schnetter & R. Schnetter 38 (COL). Santa Marta, 1898-1899, H. Smith 114, 2574 (US). Tolima: Mun. Espinal, finca El Palmar, 17 mar 1964, R. Echeverri 927 (COL). Mun. Espinal, en Calicheros en frente del molino San Martín, 27 jun 1964, R. Echeverri 1077 (COL). Departamento desconocido, J. C. Mutis 5478 (US); año 1843, I. Linden 1587 (US); Moritz 1520 (US).
ECUADOR. Azuay: 10.2 km N of Ona on the Pan American Highway at the bridge crossing the Río León, 1880 m, 24 abr 1990. P. Peterson et al. 8920 (US). Carchi: 11 km S of Mira on road to Ibarra, 1850 m, 12 may 1990, P. Peterson et al. 9094 (US). 64 km N of Cayambe on the Pan American Highway towards Tulcán, 2250 m, 12 may 1990, P. Peterson et al. 9102 (US). Islas Galápagos: Santa Cruz, Charles Darwin Research Station, near main tortoise pen, 15 abr 1983, P. Bentley 321 (US). Santa Cruz, on open spot near the two craters behind Santa Rosa, near Puerto Suárez, ca. 15 m, 13 jul 1974, H. van der Werff 1308 (COL). Guayas: Punta Carnero, road and hills side, dry rock area at the ocean, 10-30 m, 17 mar 1973, L. Holm-Mielsen et al. 2036 (COL). Pacific coast 400 m E of Punta Carnero, 0-5 m, 17 mar 1973, L. Holm-Mielsen et al. 2007, 2062 (COL). 1 km NW of Chanduy towards Atahualpa, 3-6 m, 18 mar 1973, L. Holm-Mielsen et al. 2148 (COL). Imbabura: Tababuela in Chota valley, N of Ibarra, 1700 m, 9 mar 1984, S. Lægaard 51759 (COL). Manabi: sin datos, Eggers 15701 (US). Oro: Between Machala and Puerto Bolívar, 0-10 m, 26 ago 1923, A. Hitchcock 21103 (US). Pichincha: 15 km SW of Tabacundo, 13 ene 1987, I. Grignon 84153 (US).
PERÚ. Amazonas: Prov. Bagua, Entre Bagua Grande y Chamaya, monte bajo con cactáceas y Croton, 500-600 m, 22 abr 1964, R. Ferreyra 15641 (US). Cajamarca: Prov. Cajamarca, 40 km N of Cajabamba and 13 km S of Ichocan, 2230 m, 30 mar 1997, P. Peterson & N. Refulio Rodríguez 14003 (US). Prov. Cajamarca, cerca de la localidad de Choropampa, km 124 de la carretera de penetración Pacasmayo-Cajamarca, 1700 m, 7 mar 1981, I. Sáncez Vega 2364 (US). Piura: Cerca de Bocapan, entre Talaraí y Zorritos, 100 m, 19 abr 1949, R. Ferreyra 5936 (US). Tumbes: Prov. Tumbes, El Salto, 15 m, 27 abr 1955, I. Infantes 5077 (US). Entre Tumbes y Pizarro, 21 may 1959, Velarde 7405 (US).
VENEZUELA. Anzoátegui: Alrededores del campamento de Guaraguao, Puerto La Cruz, 29 jun 1941, H. Pittier 14910 (US), 14921 (COL, US). Aragua: Ocumare de la costa, feb 1943, T. Lasser 701 (US). Carabobo: Vicinity of El Palito, road from Puerto Cabello to San Felipe, 24 sep 1920, H. Pittier 9082 (US). Carretera El Palito-Puerto Cabello, 12 ene 1980, B. Trujillo & E. Sulbarán 16129 (COL). Distrito Federal: Vicinity of El Valle, near Caracas, 28 ago 1921, H. Pittier 9709 (US). Falcón: Morón, 18 jul 1952, H. Gines 4187 (US). Lara: Around Barquisimeto, 600 m, 29 jun 1913, H. Pittier 6389 (US). Mérida: Savannas of Lagunillas, 1000 m, oct 1921, A. Jahn 661 (US). Sucre: Dtto. Sucre, parque nacional Mochima, islas Caracas del Este, 9 sep 1982, B. Garofalo 1023 (US), 1027 (US). Playa Manzanillo, W de Cumanagoto y SW de Cumaná, 1 m, 8 sep 1973, J. Steyermark et al. 108139 (COL).
10. Sporobolus tenuissimus (Mart. ex Schrank) Kuntze, Revis. Gen. Pl. 3 (3): 369. 1898. Panicum tenuissimum Mart. ex Schrank, Denkschr. Königl.-Baier. Bot. Ges. Regensburg 2: 26. 1822. TIPO: Cultivado en Munich a partir de material procedente de Brasil, Martius s.n. (holotipo M). Fig. 6 E-F.
Plantas anuales, gráciles, cespitosas. Cañas erectas, ramificadas en la base, hasta de 0,5 m de altura, raramente hasta de 0,8 m. Hojas gráciles, principalmente basales; vainas carinadas, glabras; lígulas 0,1-0,2 mm de largo, ciliadas; láminas aplanadas o involutas, acuminadas, glabras a glabrescentes, márgenes lisas, 2,5-15 cm de largo, 0,2-0,3 cm lat. Inflorescencias en panojas gráciles, abiertas, piramidales a más comúnmente oblongas, exertas, 7-30 cm long, con numerosas ramificaciones difusas y capilares, no verticiladas. Espiguillas 0,8-1 mm de largo, grisáceas, conspicuamente pediceladas, pedicelos 0,5-5 mm de largo; gluma inferior 0,1-0,3 mm de largo, oblonga, aguda, enervia, frecuentemente erosa; gluma superior 0,3-0,5 mm de largo, ovado-lanceolada, aguda, obtusa, enervia a uninervia, raramente erosa; lema 0,8-1 mm de largo, ovado-oblonga, aguda a obtusa, 1-3-nervia, los nervios apenas perceptibles; pálea elíptica, tan larga como la lema o casi igual que ella; estambres 3, anteras 0,2-0,3 mm de largo. Fruto obovoide, lateralmente comprimido, truncado en el ápice, de superficie alveolada, 0,5-0,7 mm de largo
Distribución geográfica y ecológica. Según Tovar (1993), Pohl et al. (1994), Lægaard & Peterson (2001), Peterson et al. (2001, 2003), Veldkamp (2003) y Zhenlan & Phillips (2006), S. tenuissimus es nativa del continente americano y se distribuye en el sur de los Estados Unidos de América así como en México, Mesoamérica, Venezuela, Guayanas, Ecuador, Perú (aunque no pudimos estudiar ningún espécimen peruano), Bolivia, Brasil, Paraguay y las Antillas, e introducida en África y Asia (Peterson et al. 2003); recientemente fue reportada para Singapur (Duistermaat 2004) y Thailandia (Veldkamp 2003). Las colecciones aquí consideradas son los primeros registros para Colombia (véase Peterson et al. 2001), país en el que se encuentra ampliamente distribuida. Esta especie crece en áreas abiertas y alteradas, tanto urbanas como rurales, aunque en Colombia por lo regular la hemos hallado en grietas de aceras y pavimentos de áreas urbanas, entre el nivel del mar y los 1800 m. Como dato curioso, Veldkamp (2003) comenta que en Malasia S. tenuissimus crece frecuentemente junto con Eragrostis amabilis (L.) Wight & Arn. ex Nees [= Eragrostis tenella (L.) P. Beauv. ex Roem. & Schult.], lo que concuerda con nuestras observaciones en los sitios en los que la hemos encontrado en Colombia.
Nombre vulgar. Pelo de vieja (Guzmán & Suárez 60).
Número cromosómico. 2n = 12, 40 (Peterson et al. 2003, Roodt & Spies 2003, Veldkamp 2003, Zhenlan & Phillips 2006).
Usos. En el norte de Sumatra el grano se usa para aliviar desórdenes estomacales (Veldkamp 2003). Según Veldkamp (2003), en África se emplea, en algunas ocasiones, como ornamental, lo que concuerda con algunas áreas colombianas, en las que en virtud de las delicadas y bellas inflorescencias de esta especie, se le conserva en macetas y jardineras.
Observaciones. Sporobolus tenuissimus es fácilmente reconocible por su porte grácil, por sus delicadas y exertas inflorescencias piramidales a más comúnmente oblongas, las cuales poseen numerosas ramificaciones difusas, capilares y alternas, así como por sus diminutas espiguillas.
Material examinado
COLOMBIA. Antioquia: Mun. Itagüí, creciendo en las grietas laterales de la Avenida Santa María, a la altura del barrio Simón Bolívar, entre las calles 64 y 65, cerca de la quebrada Doña María, 1550 m, 2 ene 2002, D. Giraldo-Cañas et al. 3291-A (COL, HUA). Mun. Medellín, predios internos del Jardín Botánico "Joaquín Antonio Uribe", 1550 m, 2 ene 2002, D. Giraldo-Cañas et al. 3295 (COL, HUA). Mun. Medellín, en predios de la Plaza Botero, 1550 m, 4 ene 2003, D. Giraldo-Cañas et al. 3430 (COL, HUA). Arauca: Mun. Arauca, inmediaciones de las instalaciones de la Universidad Nacional de Colombia, hacienda El Cairo, formaciones de sabanas inundables con pequeñas "matas de monte", carretera Arauca-Tame, km 9, 200-300 m, 13 jun 2003, D. Giraldo-Cañas et al. 3519 (COL). Boyacá: Mun. Santa María, en grietas de aceras del casco urbano, 900 m, 27 sep 2007, D. Giraldo-Cañas et al. 4120 (COL, HUA). Caquetá: Mun. Florencia, Barrio Villa Natalia, en un campo pedregoso en estado de sucesión temprana y en la que dominan muchas especies y géneros de gramíneas, ca. 280 m, 27-30 jun 2005, D. Giraldo-Cañas et al. 3950 (COAH, COL). Casanare: Mun. El Yopal, en áreas urbanas del centro de la ciudad, ca. 400 m, 31 oct 2007, D. Giraldo-Cañas 4136 (COAH, COL). Cauca: Mun. Popayán, predios internos y jardineras abandonadas de la Facultad de Educación de la Universidad del Cauca, 1750 m, 7-11 nov 2004, D. Giraldo-Cañas et al. 3801 (COL). Mun. Popayán, jardines externos del Banco de la República, 1750 m, 7-11 nov 2004, D. Giraldo-Cañas et al. 3816 (COL). Chocó: Mun. Quibdó, en imediaciones del Malecón y la Plaza de Mercado, ribera derecha del río Atrato, 90 m, 6 nov 2005, D. Giraldo-Cañas 3973 (CAUP, COL). Córdoba: Mun. Lorica, Estación Piscícola de la CVS, 150 m, 20 feb 2005, D. Giraldo-Cañas et al. 3840 (COL). Mun. San Pelayo, carretera San Pelayo-Lorica, en huerta casera, 150 m, 26 feb 2005, D. Giraldo-Cañas et al. 3869 (COL). Cereté, oct 1969, Guzmán & Suárez 60 (COL). Cundinamarca: Provincia Alto Magdalena, Mun. Nilo, en las afueras del Estadio Municipal, camino a la hacienda La Guaira, cerca del río Pagüey, 350 m, 16 oct 2004, D. Giraldo-Cañas et al. 3782 (COL). Provincia Gualiva, Mun. Villeta, Cordillera Oriental andina, vertiente occidental, ca. 900 m, 17-19 jun 2006, D. Giraldo-Cañas et al. 4082 (COL). Huila: Mun. Pitalito, en grietas de aceras de la zonba urbana, ca. 1550 m, 1-5 jul 2005, D. Giraldo-Cañas et al. 3922 (COL). Mun. Timaná, en grietas de los andenes del área urbana, ca. 1550 m, 1-5 jul 2005, D. Giraldo-Cañas et al. 3925 (COL). Meta: Estación Carimagua, 150 m, 18 jul 1991, C. Grollier 55 (COAH). Quindío: Mun. Quimbaya, rastrojos abiertos en los límites con un cultivo de café, ca. 1500 m, 20-26 mar 2005, D. Giraldo-Cañas & J. García-Ulloa 3879 (COL, HUA). Mun. Quimbaya, en las grietas del pavimento del parque principal, ca. 1500 m, 20-26 mar 2005, D. Giraldo-Cañas & J. García-Ulloa 3888 (COL). Mun. Armenia, en las grietas del pavimento de la Plaza de Bolívar, ca. 1500 m, 20-26 mar 2005, D. Giraldo-Cañas & J. García-Ulloa 3892 (COL). Santander: Mun. Barbosa, en las grietas de pavimentos y aceras, cerca de la plaza principal, ca. 1700 m, 20 jul 2003, D. Giraldo-Cañas et al. 3562 (COL). Tolima: Mun. Ibagué, jardineras y grietas de aceras de la Plaza de Bolívar, ca. 1200 m, 11-12 jun 2005, D. Giraldo-Cañas et al. 3905 (CAUP, COL). Vichada: Mun. Puerto Carreño, en antejardines y grietas de pavimentos del casco urbano, en inmediaciones de la Plaza Central y la Gobernación, 50 m, 6 ene 2004, D. Giraldo-Cañas & C. Parra 3669 (CAUP, COL).
ECUADOR. Pichincha: Alluriquin, 850 m, 7 feb 1987, I. Grignon 84211 (MO). Road Quito-Santo Domingo, at Tinelandia, 600 m, 17 ago 1984, S. Lægaard 52665 (COL). Alluriquin, 850 m, 22 sep 1984, S. Lægaard 53034 (COL, US). 21 km W of Santo Domingo on road to Chone, then from Nuevo Israel, 320 m, 5 jun 1990, P. Peterson & E. Judziewicz 9523 (US).
VENEZUELA. Distrito Federal: La Florida, near Caracas, 900 m, 27 nov 1938, A. Alston 5234 (MO). Monagas: Caripite, 30 mar 1940, A. Chase 12590 (US). Sucre: Cumaná, Funck 229 (US).
11. Sporobolus virginicus (L.) Kunth, Revis. Gramin. 1: 67. 1829. Agrostis virginica L., Sp. Pl. 1: 63. 1753. TIPO: Estados Unidos de América. Virginia: J. Clayton 507 (lectotipo LINN-84.30, designado por Hitchcock, Contr. U.S. Natl. Herb. 12: 119. 1908; isolectotipos BM, US-76294, fragm. ex BM!). Fig. 7 C-E.
Plantas perennes, fuertemente rizomatosas, los rizomas escamosos y reptantes, e incluso estoloníferas. Cañas erectas a decumbentes, hasta de 0,6 m de altura, enraizadas y ramificadas en los nudos inferiores. Hojas principalmente caulinares, conspicuamente dísticas, las basales sobrelapadas; vainas imbricadas, redondeadas, glabras o pilosas, ciliadas; lígulas 0,2-0,4 mm de largo, ciliadas; láminas aplanadas a involutas, lineares, abaxialmente glabras, adaxialmente pilosas, márgenes escabrosas, terminando en una punta muy fina, 2-16 cm de largo, 0,2-0,5 mm lat. Inflorescencias en panojas densas, contraídas a espiciformes, lanceoladas a lineares, erectas, 2,5-9 cm de largo, con las ramificaciones cortas y floríferas desde su base, no verticiladas. Espiguillas 1,7-2,5 (-3,1) mm de largo, grisáceas o pajizas, lateralmente comprimidas; gluma inferior lanceolada, aguda, acuminada, uninervia, (1,2-) 1,5-2,3 mm long; gluma superior tan larga como la espiguilla o levemente más corta, ovado-elíptica, aguda a acuminada, uninervia; lema ovada, aguda, uninervia, glumiforme, ligeramente más corta que la gluma superior; pálea ovada, aguda, hialina, tan larga como la lema o ligeramente más corta; estambres 3, anteras 0,9-1,4 mm de largo. Fruto subgloboso a obovado o elíptico, ligeramente comprimido, redondeado en el ápice, 0,7-1,1 mm de largo
Distribución geográfica y ecológica. Según Tovar (1993), Pohl et al. (1994), de Castro Boechat & Longhi-Wagner (1995), Lægaard & Peterson (2001) y Peterson et al. (2001, 2003), S. virginicus crece desde los Estados Unidos de América y el Caribe hasta Brasil y Chile, así como en las áreas paleotropicales (Clayton 1965). A pesar de esta amplia distribución, no se conocía su presencia en Colombia (véase Peterson et al. 2001), la cual destacamos en esta contribución. Esta especie crece principalmente en las playas marinas, dunas y en ambientes de suelos salinos de tierras bajas.
Nombres vulgares. Faxtil (P. Lowy & F. González 180), hierba amarga (N. de López 431), paja (E. Mancera 1), pasto de amor (R. Echeverry 3409), pasto de salitre (N. de López 431). Según Luces (1942), a S. virginicus se le conoce en Venezuela con el nombre de saladilla.
Número cromosómico. 2n = 18, 20, 30 (Peterson et al. 2003, 2004, Roodt & Spies 2003, Zhenlan & Phillips 2006).
Observaciones. Pohl et al. (1994: 276) y Peterson et al. (2003: 121) comentan que esta especie raramente produce semillas, probablemente porque es autoincompatible y la mayoría de las poblaciones son clonales y de un solo genotipo.
Material examinado
COLOMBIA. Atlántico: Barranquilla, Las Flores-La Playa, saline marshes, 0-2 m, 13 sep 1959, J. Cuatrecasas 24286 (COL, US). Puerto Colombia, playa de Miramar, 10 jul 1962, A. Dugand 6099 (COL). Puerto Colombia, Sabanilla, 21 nov 1912, A. Hitchcock 9925 (US). Bolívar: Cartagena, en las playas de las inmediaciones de El Laguito, cerca de los apartamentos "Cristóforo Colombo", 0-5 m, 16 oct 2002, D. Giraldo-Cañas 3308 (COL). Cartagena, hacia La Boquilla, 30 jul 1943, A. Dugand & R. Jaramillo 3411 (US). Archipiélago de San Bernardo, isla Panda, 24 jun 2002, C. Flórez & P. Sarmiento 67 (COL), 16 abr 2003, C. Flórez 89 (COL). Cartagena, sitio San Andrés, 31 mar 1995, F. Arias & H. Guevara 75 (COL). Cartagena, sandy sea-beach, 0-2 m, 1-3 oct 1922, F. Pennell 12014 (US). Cartagena, 18 km SW of crossing Canal del Dique at Pasacaballos, Isla Barú, along playa Mohán, 0-2 m, 6 ago 1985, J. Zarucchi & H. Cuadros 3964 (MO, US). Magdalena: Santa Marta, Rodadero de Santa Marta, 1 m, 11 nov 1979, R. Echeverry 3409 (COL, US). Playas en los alrededores de Santa Marta, 15 m, 9 ene 1948, J. Giacometto et al. 18C023 (US). En la margen izquierda del río Magdalena, 13 jul 1970, N. de López 431 (COL). Pozos Colorados, 29 oct 1987, E. Mancera 1 (COL). Ciénaga, alrededores de Aguacoca, 20 m, 21 feb 1959, R. Romero-Castañeda 7250 (US). Santa Marta, sea level, 1898-1901, H. Smith 113 (US). San Andrés, Providencia y Santa Catalina: Providencia, Iron Wood Hill, 21 jul 1990, P. Lowy & F. González 180 (COL). San Andrés, 363 m, 17 jul 1966, J. Torres et al. 190 (COL). Sucre: Islas de San Bernardo, isla Boquerón, 0-1 m, 11 ago 1979, H. Bernal 109 (COL). Departamento desconocido, sin datos y sin fecha, Moritz 1564 (US).
ECUADOR. Esmeraldas: Atacames, 0-50 m, 28 ene 1987, I. Grignon 84208 (US). Islas Galápagos: Santa Cruz, Pelican Bay, 10 ene 1983, P. Bentley 149 (US). Guayas: Guayaquil, sandy seashore at Estero Salado, 6 feb 1955, E. Asplund 15345 (US). Manabi: Coast N of Crusita, 0-5 m, 24 sep 1984, S. Lægaard 53071 (COL, US). San Clemente, old salinas, dune-depressions, low dunes, 0-5 m, 10 mar 1988, S. Lægaard & S. Renvoize 70701 (COL). Oro: Near Puerto Bolívar, 30 abr 1956, J. Hjerting & K. Rahn 629 (US).
PERÚ. Arequipa: Prov. Camana, shore of Pacific Ocean, 1 km ESE of Puerto Camana, sea level, 14 ene 1963, H. Iltis et al. 1524 (US). Prov. Islay, 1 km S of Mollendo, sea level, 23 abr 1939, W. Eyerdam 25372 (US). La Libertad: Prov. Trujillo, Facultad de Medicina de Trujillo, 60 m, 12 ene 1967, E. Sánchez 6281 (US). Lambayeque: Prov. Chiclayo, 1-3 m, 21 jun 1947, F. Fosberg 27973 (US), 27982 (US). Lima: Cerca de Venanilla y Ancón, 5 m, 12 oct 1947, R. Ferreyra 2475 (US).
VENEZUELA. Anzoátegui: Mun. Peñalver, costa arenosa cerca del Hatillo, laguna de Unare, mar Caribe, 0-5 m, oct 1996, A. Fernández 9658 (HUA). Aragua: Ocumare, 29 may 1937, H. Pittier 14015 (US). Carabobo: Puerto Cabello, sin fecha, H. Curran & M. Haman 1160 (US). Puerto Cabello, 19 ago 1897, Lehmann 8835 (US). Distrito Federal: Macuto, 0-3 m, 8 oct 1944, J. Steyermark 59317-a (US). Puerto Caribe, ene 1937, F. Tamayo 177 (US). Playas arenosas de Macuto y Caribe, ago 1940, F. Tamayo 1358 (US). Falcón: Distrito Silva, Cayo del Medio, al SE de Tucacas, 1 m, 3 sep 1974, J. Steyermark & B. Manara 110695 (COL). Miranda: Tocaripua, 0-10 m, 27 feb 1949, H. Gines 4526 (US). Paparo, at the mouth of Río Grande del Tuy, Barlovento, near sea level, 11-16 jun 1913, H. Pittier 6346 (US). Nueva Esparta: El Robledar, dic 1951, H. Gines 2641 (US). Sucre: Cumaná, sin fecha, Funck 601 (US). Zulia: Santa Rosa, near Maracaibo, 6 oct 1922, H. Pittier 10476 (US). Maracaibo, sin fecha, Plée 23 (COL). Playas del caribe, departamento Vargas, 5 ago 1943, T. Lasser 756 (COL). Isla Zapara, Barra del Lago de Maracaibo, 22 ago 1921, A. Jahn 501 (US).
ESPECIE DUDOSA
Sporobolus humifusus (Kunth) Kunth, Revis. Gramin. 1: 67. 1829. Vilfa humifusa Kunth, Nov. Gen. Sp. (quarto ed.) 1: 137. 1816. TIPO: Venezuela. Sucre: Cumaná, F. Humboldt & A. Bonpland s.n. (holotipo P, isotipos B, BAA).
De esta especie no pudimos tener acceso a ningún material, y al parecer, sólo se conoce del material tipo. Además, la descripción original de ésta es muy corta y no detalla ni resalta ninguna característica que le sea exlusiva a la especie. Todo lo anterior no nos permite hacer conjeturas acerca de dicho binomio.
ESPECIES EXCLUIDAS
Sporobolus brasiliensis (Raddi) Hack., Bull. Herb. Boissier, ser. 2, 4 (3): 278. 1904. Aira brasiliensis Raddi, Agrostogr. Bras. 36. 1823. TIPO: Brasil. Rio de Janeiro, Raddi s.n. (holotipo PI, isotipo LE) (= Eragrostis airoides Nees).
Eragrostis airoides Nees, una enigmática especie sudamericana (presente en Argentina, Bolivia, Brasil, Colombia, Paraguay, Uruguay, Venezuela y el Caribe), frecuentemente tratada como Sporobolus brasiliensis (Raddi) Hack. (Peterson & Giraldo-Cañas 2008), en virtud de su número cromosómico básico de x = 9, sus espiguillas paucifloras (1-3 antecios) y la morfología de su cariopsis, pero la posesión de espiguillas con no sólo un antecio sino frecuentemente con dos o tres y el modo de desarticulación de la espiguilla, son caracteres que respaldan la inclusión de S. brasiliensis en el género Eragrostis (Peterson & Giraldo-Cañas 2008).
Sporobolus domingensis (Trin.) Kunth, Enum. Pl. 1: 214. 1833. Vilfa domingensis Trin., Neue Entdeck. Pflanzenk. 2: 59. 1821. Sporobolus pyramidatus (Lam.) Hitchc. subesp. domingensis (Trin.) Cat. Guerra, Fontqueria 44: 143. 1996. TIPO: Hab. in Domingo, Cl. Poiteau s.n. (holotipo LE-TRIN-1694.01!, isotipo US-91124!).
Sporobolus domingensis se distribuye en el sur de los Estados Unidos de América, México y el Caribe, y fue citada para Venezuela por Peterson et al. (2001: 207). No obstante, no pudimos detectar ejemplares venezolanos de la misma y por lo tanto, la excluimos de este tratamiento taxonómico.
AGRADECIMIENTOS
Queremos manifestar nuestro profundo agradecimiento a la Universidad Nacional de Colombia y a su Instituto de Ciencias Naturales, así como al Smithsonian Institution y al Herbario Nacional de los Estados Unidos de América (US) por las facilidades permanentes para llevar a cabo los estudios en gramíneas. A J. F. Veldkamp (L), S. Renvoize (K) y S. Lægaard (AAU) por su colaboración, comentarios y por el obsequio de valiosa bibliografía. El Smithsonian Institution, así como el Herbario Nacional de los Estados Unidos de América (US), financiaron el viaje y la estadía del primer autor en Washington. Agradecemos a la Utah State University por su permiso para usar las ilustraciones que aparecieron en la Flora of North America (volumen 25), las cuales fueron realizadas por Linda A. Vorobik y Hana Pazdirkova; A Simon Lægaard por permitirnos usar la ilustración que apareció en la Flora of Ecuador [214. Gramineae (part 2) subfam. Chloridoideae]; A Lynn G. Clark por permitirnos usar una de las figuras que apareció en la Flora Costaricensis (Family #15, Gramineae). Por último, queremos agradecer a Alice R. Tangerini (US) por la elaboración de las restantes figuras y por la preparación de las planchas. Esta contribución es derivada del proyecto "Estudios sistemáticos en gramíneas de Colombia. Parte II" de la Universidad Nacional de Colombia, sede Bogotá.
LITERATURA CITADA
1. ASTEGIANO, M. 1986. La cleistogamia y casmogamia en Sporobolus indicus (Poaceae). Kurtziana 18: 69-76. [ Links ]
2. BAAIJENS, G. & J. VELDKAMP. 1991. Sporobolus (Gramineae) in Malesia. Blumea 35 (2): 393-458. [ Links ]
3. BOR, N. 1960. Pooideae-Sporoboleae. En: The grasses of Burma, Ceylan, India and Pakistan: 622-636. Pergamon Press, Londres. [ Links ]
4. BRANDENBURG, D. 2003. Notes on free pericarps in grasses (Poaceae). J. Ky. Acad. Sci. 64 (2): 114-120. [ Links ]
5. BROWN, R. 1810. Prodromus Florae Novae Hollandiae. Londres. [ Links ]
6. CAMPBELL, C., J. QUINN, G. CHEPLICK & T. BELL. 1983. Cleistogamy in grasses. Annu. Rev. Ecol. Syst. 14: 411-441. [ Links ]
7. CHASE, A. & Z. LUCES DE FEBRES. 1972. Primer libro de las gramíneas. Instituto Interamericano de Ciencias Agrícolas de la OEA, Lima. [ Links ]
8. CIALDELLA, A. & A. VEGA. 1996. Estudios sobre la variación estructural de las espiguillas en géneros de la tribu Paniceae (Poaceae). Darwiniana 34: 173-182. [ Links ]
9. CLAYTON, W. 1965. Studies in the Gramineae: VI. Kew Bull. 19 (2): 287-296. [ Links ]
10. CLAYTON, W. & S. RENVOIZE. 1986. Genera Graminum: Grasses of the world. Kew Bull., Additional series 13: 1-389. [ Links ]
11. COLBRY, V. 1957. Diagnostic characteristics of the fruits and florets of economic species of North American Sporobolus. Contr. U.S. Natl. Herb. 34 (1): 1-24. [ Links ]
12. COLUMBUS, J. T., R. CERROS-TLATILPA, M. S. KINNEY, M. E. SIQUEIROS-DELGADO, H. L. BELL, M. P. GRIFFITH & N. F. REFULIO-RODRÍGUEZ. 2007. Phylogenetics of Chloridoideae (Gramineae): A preliminary study based on nuclear ribosomal internal transcriber spacer and chloroplast trnL-F sequences. Aliso 23: 565-579. [ Links ]
13. CONNOR, H. 1979. Breeding systems in the grasses. New Zealand J. Bot. 17: 547-574. [ Links ]
14. CONNOR, H. 1981. Evolution of reproductive systems in the Gramineae. Ann. Missouri Bot. Gard. 68: 48-74. [ Links ]
15. CONNOR, H. 1987. Reproductive biology in the grasses. En: T. Soderstrom, K. Hilu, C. Campbell & M. Barkworth (eds.), Grass systematics and evolution: 117-132. Smithsonian Institution Press, Washington D. C. [ Links ]
16. DÁVILA ARANDA, P. & E. MANRIQUE FORCECK. 1990. Glosario de términos agrostológicos. Cuadernos del Instituto de Biología 5, Universidad Nacional Autónoma de México UNAM, México, D. F. [ Links ]
17. DE CASTRO BOECHAT, S. & H. LONGHI-WAGNER. 1995. O gênero Sporobolus (Poaceae: Chloridoideae) no Brasil. Acta Bot. Bras. 9 (1): 21-86. [ Links ]
18. DUISTERMAAT, H. 2004. New grass (Poaceae) records for Singapore, including Panicum laxum new fos Asia. Garden's Bulletin Singapore 56 (1): 29-42. [ Links ]
19. ELLIS, R. 1977. Distribution of Kranz syndrome in the souther African Eragrostoideae and Panicoideae according to bundle sheat anatomy and cytology. Agroplantae 9: 73-110. [ Links ]
20. FONT QUER, P. 1993. Diccionario de Botánica. Ed. Labor, S. A., Barcelona. [ Links ]
21. GIRALDO-CAÑAS, D. 2008. Sistemática del género Axonopus (Poaceae: Panicoideae: Paniceae) y revisión de las especies de la serie Barbigeri. Serie Biblioteca José Jerónimo Triana 17: 1-211. Instituto de Ciencias Naturales, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá D. C. [ Links ]
22. GOOSSENS, A. 1938. A study of the South African species of Sporobolus R. Br., with special reference to leaf anatomy. Trans. Royal Soc. South Africa 26 (2): 173-223. [ Links ]
23. GOULD, F.& R. SHAW. 1992. Gramíneas. Clasificación Sistemática. AGT Editor S.A., México D. F. [ Links ]
24. HILU, K. & L. ALICE. 2001. A phylogeny of Chloridoideae (Poaceae) based on matK sequences. Syst. Bot. 26 (2): 386-405. [ Links ]
25. HOLMGREN, P., N. HOLMGREN & L. BARNETT. 1990. Index Herbariorum. Part I: The Herbaria of the World. The New York Botanical Garden, Nueva York. [ Links ]
26. KIESLING, R. 2002. Código Internacional de Nomenclatura Botánica (Código de Saint Louis). Edición en español, Instituto de Botánica Darwinion-Missouri Botanical Garden, Buenos Aires. [ Links ]
27. LÆGAARD, S. & P. PETERSON. 2001. Gramineae (part 2, Subfam. Chloridoideae). En: G. Harling & L. Andersson (eds.), Flora of Ecuador 68: 1-131. [ Links ]
28. LIU, Q., N. ZHAO & G. HAO. 2005a. Inflorescense structures and evolution in subfamily Chloridoideae (Gramineae). Plant Syst. Evol. 251: 183-198. [ Links ]
29. LIU, Q., N. ZHAO, G. HAO, X. HU & Y. LIU. 2005b. Caryopsis morphology of the Chloridoideae (Gramineae) and its systematic implications. Bot. J. Linnean Soc. 148: 57-72. [ Links ]
30. LUCES, Z. 1942. Géneros de las gramíneas venezolanas. Ministerio de Agricultura y Cría, Boletín Técnico Nro. 4, Tipografía Garrido, Caracas. [ Links ]
31. MCCLURE, F. & T. SODERSTROM. 1972. The agrostological term Anthecium. Taxon 21: 153-154. [ Links ]
32. MORA-OSEJO, L. E. 2004. Morfología, sistemática y evolución de las Angiospermae. Departamento de Biología-Instituto de Ciencias Naturales, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá D. C. [ Links ]
33. MORRONE, O. & F. ZULOAGA. 1991. Estudios morfológicos en el subgénero Dichanthelium de Panicum (Poaceae: Panicoideae: Paniceae), con especial referencia a Panicum sabulorum. Ann. Missouri Bot. Gard. 78: 915-927. [ Links ]
34. NICORA, E. & Z. RÚGOLO DE AGRASAR. 1987. Los géneros de gramíneas de América austral. Ed. Hemisferio Sur, Buenos Aires. [ Links ]
35. ORTIZ-DÍAZ, J-J. & A. CULHAM. 2000. Phylogenetic relationships of the genus Sporobolus (Poaceae: Eragrostideae) based on nuclear ribosomal DNA ITS sequences. En: S. Jacobs & J. Everet (eds.), Grasses: Systematics and evolution: 184-188. CSIRO, Melbourne. [ Links ]
36. PARODI, L. 1964. Gramíneas bonaerenses. Clave para la determinación de los géneros y enumeración de las especies. Colección Harold Cyril Martin, Talleres Gráficos Ernesto Zeiss, Buenos Aires. [ Links ]
37. PETERSON, P. M., T. COLUMBUS, & S. PENNINGTON. 2007. Classification and biogeography of New World grasses: Chloridoideae. En: J. T. Columbus, E. A. Friar, J. M. Porter, L. M. Prince & M. G. Simpson (eds.), Monocots. Comparative Biology and Evolution. Poales. Aliso 23: 580-594. [ Links ]
38. PETERSON, P. M. & D. GIRALDO-CAÑAS. 2008. Eragrostis (Poaceae: Chloridoideae: Eragrostideae) in Colombia. J. Bot. Res. Inst. Texas 2 (2): 875-916. [ Links ]
39. PETERSON, P. M., S. HATCH & A. WEAKLEY. 2003. Sporobolus. En: M. Barkworth, K. Capels, S. Long & M. Piep (eds.), Flora of North America-North of Mexico 25: 115-139. [ Links ]
40. PETERSON, P. M., R. SORENG, G. DAVIDSE, T. FILGUEIRAS, F. ZULOAGA & E. JUDZIEWICZ. 2001. Catalogue of New World grasses (Poaceae): II. Subfamily Chloridoideae. Contr. U.S. Natl. Herb. 41: 1-255. [ Links ]
41. PETERSON, P. M., J. VALDÉS-REYNA & J. ORTIZ-DÍAZ. 2004. Sporobolus (Poaceae: Chloridoideae: Cynodonteae: Zoysieae: Sporobolinae) from northeastern Mexico. Sida 21 (2): 553-589. [ Links ]
42. PETERSON, P. M., R. D. WEBSTER & J. VALDÉS REYNA. 1995. Subtribal classification of the New World Eragrostideae (Poaceae: Chloridoideae). Sida 16: 529-544. [ Links ]
43. PETERSON, P. M., R. D. WEBSTER & J. VALDÉS-REYNA. 1997. Genera of New World Eragrostideae (Poaceae: Chloridoideae). Smithsonian Contr. Bot. 87: 1-50. [ Links ]
44. PILGER, R. 1956. Sporobolinae. En: H. Melchior & E. Werdermann (eds.), Gramineae II, Unterfamilien: Micraiodeae, Eragrostideae, Orizoideae, Olyroideae. Die Naturlichen Pflanzenfamilien, 2a. edición, 14: 53-67. [ Links ]
45. PINTO-ESCOBAR, P. 2002. Las gramíneas en Colombia. Revista Acad. Colomb. Cienc. 26 (98): 59-68. [ Links ]
46. POHL, R., C. REEDER & G. DAVIDSE. 1994. Sporobolus. En: G. Davidse, M. Sousa & A. Chater (eds.), Flora Mesoamericana 6: 273-276. [ Links ]
47. REINHEIMER, R., M. E. ASTEGIANO & A. C. VEGETTI. 2005. Tipología de las inflorescencias en especies de Sporobolus (Sporobolinae-Eragrostideae-Poaceae). Kurtziana 31 (1-2): 7-19. [ Links ]
48. ROODT, R. & J. SPIES. 2003. Chromosome studies in the grass subfamily Chloridoideae. I. Basic chromosome numbers. Taxon 52: 557-566. [ Links ]
49. ROTH, I. 1977. Fruits of Angiosperms. Gebrüder Borntraeger, Berlín-Stuttgart. [ Links ]
50. RUA, G. 1999. Inflorescencias, bases teóricas para su análisis. Sociedad Argentina de Botánica, Buenos Aires. [ Links ]
51. RÚGOLO DE AGRASAR, Z. & M. PUGLIA. 2004. Plantas de la Argentina, silvestres y cultivadas. Gramíneas ornamentales. Editorial LOLA, Buenos Aires. [ Links ]
52. SCHWABE, H. 1949. Contribución al estudio anatómico de las especies argentinas del género Sporobolus y sus relaciones con los géneros afines. Bol. Soc. Argent. Bot. 2 (4): 253-270. [ Links ]
53. SIMON, B. & S. JACOBS. 1999. Revision of the genus Sporobolus (Poaceae, Chloridoideae) in Australia. Australian Syst. Bot. 12: 375-448. [ Links ]
54. SORENG, R. & J. DAVIS. 1998. Phylogenetics and character evolution in the grass family (Poaceae): Simultaneous analysis of morphological and chloroplast DNA restriction site character sets. Bot. Rev. 64: 1-85. [ Links ]
55. SOUSA, M.& S. ZÁRATE. 1988. Flora Mesoamericana. Glosario para Spermatophyta, español-inglés. Universidad Nacional Autónoma de México, México D. F. [ Links ]
56. STAPF, O. 1898. Gramineae. En: W. Thiselton-Dyer (ed.), Flora Capensis 7: 315, 578-589. [ Links ]
57. TOVAR, O. 1993. Las gramíneas (Poaceae) del Perú. Ruizia 13: 1-480. [ Links ]
58. VALDÉS-REYNA, J. & S. HATCH. 1991. Lemma micromorphology in the Eragrostideae (Poaceae). Sida 14 (4): 531-549. [ Links ]
59. VAN DEN BORRE, A. & L. WATSON. 1997. On the classification of the Chloridoideae (Poaceae). Australian Syst. Bot. 10: 491-531. [ Links ]
60. VAN DEN BORRE, A. & L. WATSON. 2000. On the classification of the Chloridoideae: Results from morphological and leaf anatomical data analyses. En: S. Jacobs & J. Everet (eds.), Grasses: Systematics and evolution: 180-183. CSIRO, Melbourne. [ Links ]
61. VEGETTI, A. & J. PENSIERO. 1993. Tipología de la inflorescencia de Panicum sección Panicum (Poaceae: Paniceae). Bol. Soc. Argent. Bot. 29: 7-10. [ Links ]
62. VELDKAMP, J. F. 2002. Revision of Eragrostis (Gramineae, Chloridoideae) in Malesia. Blumea 47: 157-204. [ Links ]
63. VELDKAMP, J. 2003. Miscellaneous notes on Thai Gramineae. Blumea 48: 495-501. [ Links ]
64. WATSON, L. & M. DALLWITZ. 1999. Grass genera of the World: descriptions, illustrations, identification, and information retrieval; including synonyms, morphology, anatomy, physiology, phytochemistry, cytology, classification, pathogens, world and local distribution, and references. http://biodiversity.uno.edu/delta/. Versión: 28 de mayo de 1999. [ Links ]
65. ZHENLAN, W. & S. PHILLIPS. 2006. Sporobolus. Flora of China 22: 482-485. [ Links ]
66. ZULOAGA, F. & T. SODERSTROM. 1985. Classification of the outline species of New World Panicum (Poaceae: Paniceae). Smithsonian Contr. Bot. 59: 1-63. [ Links ]
Recibido: 05/06/2008
Aceptado: 23/11/2008

