










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232
Caldasia vol.34 no.2 Bogotá July/Dec. 2012
ANTONIO GALÁN DE MERA
JOSÉ ALFREDO VICENTE ORELLANA
ELIANA LINARES PEREA
JOSÉ CAMPOS DE LA CRUZ
CARLOS TRUJILLO VERA
FRANCISCO VILLASANTE BENAVIDES
Departamento de Biología (Botánica), Facultad de Farmacia, Universidad San Pablo-CEU, Apartado 67, 28660 Boadilla del Monte, Madrid, España. agalmer@ceu.es. Autor para correspondencia: avicore@ceu.es
Estudios Fitogeográficos del Perú, Sánchez Cerro 219, Manuel Prado, Paucarpata, Arequipa, Perú. elialinper@hotmail.com
Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Apartado 140434, Lima 14, Perú. joricampos@yahoo.es
Escuela de Geografía, Facultad de Ciencias Histórico Sociales, Universidad Nacional de San Agustín, Arequipa, Perú. cartruve@hotmail.com
Departamento de Biología, Facultad de Ciencias Biológicas y Agropecuarias, Universidad Nacional de San Agustín, Arequipa, Perú. fvillabe@gmail.com
RESUMENEl presente trabajo trata de establecer una relación entre las comunidades de Cactáceas y su distribución en las vertientes occidentales de los Andes del Perú. Para ello empleamos distintos valores climáticos de precipitación y evapotranspiración potencial, y los índices climáticos de termicidad y aridez. Sobre una matriz con la composición florística de las comunidades aplicamos el índice de Sørensen obteniendo un dendrograma que explica cómo se agrupan las comunidades según provincias biogeográficas y pisos bioclimáticos. Aplicando sobre las distintas localidades seleccionadas un Análisis de Componentes Principales (ACP) podemos deducir cómo se agrupan en función de la altitud y los valores e índices climáticos. Las poblaciones de Cactáceas situadas al norte del paralelo 8°Sur son las más influidas por la humedad aportada por la corriente de El Niño; bajo el paralelo 8°Sur, las localidades situadas en las vertientes occidentales andinas dependen de la escasa humedad estival (HE), aunque en el sur también puede ser evidente la invernal (HI) al recibir las lluvias de depresiones aisladas que se desprenden de borrascas sub-antárticas entre julio y agosto; no obstante, los andenes de cultivo ancestrales reflejan un clima más húmedo que el actual. En la costa y áreas basales próximas al desierto abiótico, la aridez es muy elevada aunque aún están presentes las comunidades con Neoraimondia arequipensis. Las comunidades situadas al norte del paralelo 8° Sur pertenecen a la provincia biogeográfica Pacífica, las del centro a la provincia Ancashino-Paceña, las de los departamentos de Arequipa, Moquegua y Tacna, a la provincia Oruro-Arequipeña, y las del Desierto Pacífico a la provincia Limeño-Ariqueña.
Palabras clave. Cactaceae, biogeografía, bioclimatología, El Niño, corriente de Humboldt, Andes, Perú.ABSTRACT
Using climatic values of precipitation and potential evapotranspiration, as well as climate indices of thermicity and aridity, in this work we try to establish a relationship between cactus communities and their distribution in the western slopes of the Peruvian Andes. Using the Sørensen index on a matrix with the floristic composition of plant communities, we obtained a dendrogram that explains their grouping according to biogeographic provinces and bioclimatic belts. Using a Principal Component Analysis (PCA) on selected localities, we can infer how they are grouped according to the altitudinal gradient and climatic values and indices. Cactus populations located north of the 8° parallel south are the most influenced by the humidity provided by El Niño stream; those populations south of the 8° parallel south, on the western slopes of the Andes, depend on the low summer humidity (HE), although it can also be evident the winter humidity (HI) due to rainfalls of isolated depressions that emerge from sub-Antarctic storms between July and August. However, the crop terraces of ancestral cultures reflect a more humid ancient climate. In the coast and basal areas of the Andes, next to the abiotic desert, the aridity is very high, although communities with Neoraimondia arequipensis are still present. Communities located north of the 8° parallel south belong to the Pacific biogeographic province, those of the central Peru belong to the Ancash-La Paz province, those of the Arequipa, Moquegua and Tacna departments to the Oruro-Arequipa province, and finally those communities of the Pacific Desert belong to the Lima-Arica province.
Key words. Cactaceae, biogeography, bioclimatology, El Niño, Humboldt stream, Andes,Peru.Recibido: 14/01/2012
Aceptado: 01/10/2012
INTRODUCCIÓN
La Cordillera Andina se extiende a lo largo de 9000 km en América del Sur. En Perú se inicia en la Cordillera del Cóndor, al norte, y finaliza al sur en la Cordillera del Barroso, dentro de los Andes tropicales. Su vegetación está influida por la situación anticiclónica debida a la corriente fría de Humboldt, la sombra de lluvias causada por las altitudes de la cordillera, las lluvias irregulares de la corriente de El Niño y, al oriente, las lluvias amazónicas (Galán de Mera et al. 1997, Fagan 2009). El rasgo más destacado de su relieve son sus grandes altitudes con el vulcanismo terciario y cuaternario en el sur (Lissón 1980)(p.ej. Ampato -6288 m-, Coropuna -6305 m-, Huascarán -6768 m-, Solimana -6093 m-) y valles muy profundos (Colca, Cotahuasi), que aumenta la diversidad de comunidades presididas por Cactáceas, y una acción antrópica que se refleja en la ganadería extensiva y en un uso agrícola del suelo ancestral sustentado en los "ayllus" (unidad básica de población pre-hispanica) y, posteriormente, en las poblaciones de estructura española (Murra 2009).
El origen de la vegetación presidida por Cactáceas en los Andes occidentales parece ir asociada al Mioceno, pues en esta época ya se evidenciaban en el norte de Chile fenómenos de aridez de origen anticiclónico (Hartley 2003) potenciados por el desarrollo de la corriente de Humboldt y la orogenia andina (Garzione et al. 2008). La separación gradual de la Antártida de América del Sur tuvo lugar durante el Eoceno, hace entre 50 y 30 Ma (Livermore et al. 2005), lo que aisló a la Antártida de las aguas marinas cálidas y contribuyó a la expansión de los glaciares, al enfriamiento del mar, y por tanto, al fortalecimiento de la corriente fría de Humboldt. Por otra parte, el cierre del corredor oceánico de América Central a finales del Plioceno (Ibaraki 1997) produjo una expansión de la corriente de Humboldt hacia el norte coincidiendo con el comienzo del periodo de hiperaridez en el desierto de Atacama (Luebert & Gajardo 2005).
En el oeste de América del Sur hace unos 15 Ma, la combinación entre la situación anticiclónica debida a la corriente de Humboldt y la sombra de lluvias de los Andes, diferenciaron elementos florísticos pre-adaptados a la aridez de las vertientes occidentales andinas (Graham 2009); por ello, posiblemente en el Mioceno comienza la especiación y endemicidad de las Cactáceas en los Andes occidentales, acompañadas de algunos xerófitos (Villagrán & Hinojosa 2005), como Prosopis L., Acacia Willd., Loxopterygium Hook., Bulnesia Gay (Solbrig 1976), Larrea Cav. (Schwarzer et al. 2010) o Bougainvillea Comm. ex Juss. (Linares Perea et al. 2010), cuya vegetación quedó separada por la Cordillera.
Schwarzer et al. (2010) sugieren una estrecha relación entre los efectos del vulcanismo moderno y las comunidades vegetales ante la explosión del volcán Huaynaputina hace 400 años y que, en general, el vulcanismo terciario y cuaternario en el sur del Perú originó la formación de profundos valles interandinos que aislaron incluso a endemismos orientales formándose otros asociados a determinadas condiciones geomorfológicas procedentes de eventos volcánicos de diferente edad. Estos eventos coinciden con la interpretación filogenética de otros autores (Hershkovitz & Zimmer 1997) que sugieren, con evidencias de ADN de cloroplastos de Cactáceas, un origen de esta familia hace unos 30 Ma, a mediados del Terciario, y una diversificación posterior a finales de este periodo coincidente con el desarrollo del Desierto Pacífico. Similares resultados aportan las paleomadrigueras de roedores fósiles del sur del Perú (Holmgren et al. 2001) que indican un predominio de Cactáceas durante el Holoceno.
La vegetación occidental actual del sur del Perú se puede sintetizar con la de las lomas costaneras del Desierto Pacífico (250- 1000 m), las formaciones de cactus columnares (1200- 3100 m), separada de la anterior por un desierto abiótico que puede tener una extensión de 100 Km de ancho, arbustedas y matorrales (3100- 3800 m), pajonales de la puna (3800- 4800 m), y las comunidades ligadas a la geomorfología periglacial (> 4800 m), que en realidad son las formaciones vegetales de todo el occidente peruano (Weberbauer 1912, 1945) bajo la influencia de la corriente de Humboldt. Las comunidades con cactus están situadas en las lomas del desierto costero y en los pisos basales de los Andes. Su diversidad es debida a la endemicidad y a las condiciones ecológicas. Por ello, el objetivo de este trabajo es profundizar en cómo están relacionadas con diferentes parámetros e índices climáticos, cuyos valores están influidos por la corriente de Humboldt, El Niño, o las lluvias andinas de origen amazónico.
MATERIALES Y MÉTODOS
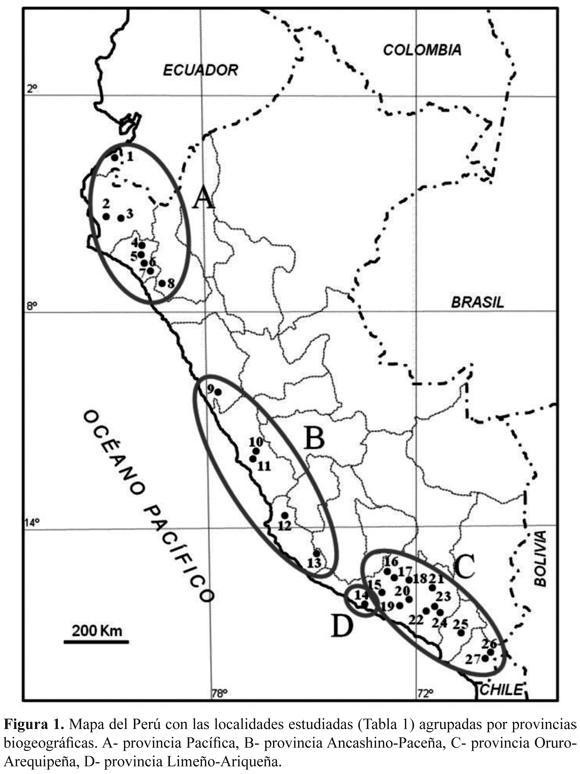
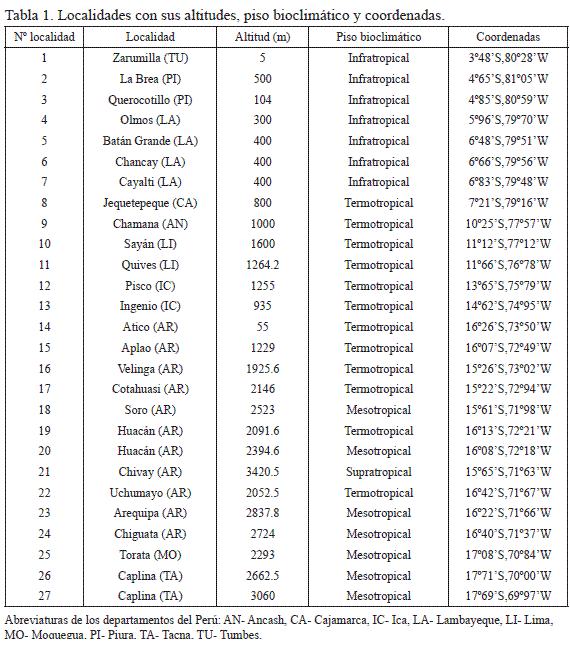
El territorio estudiado y muestreos seleccionados. Se seleccionaron 27 localidades a lo largo de la costa y de las vertientes occidentales de los Andes peruanos (Fig. 1, Tabla 1). En ellas se han levantado muestreos siguiendo el método fitosociológico de Braun-Blanquet (1932), tomando como base trabajos ya publicados (Galán de Mera & Vicente Orellana 1996, Galán de Mera & Gómez Carrión 2001, Galán de Mera et al. 2002, 2004, 2009), y nuevos inventarios realizados entre los años 2008 y 2011. En total se han manejado 88 muestreos y 146 especies.
Fuentes botánicas. Las especies de Cactáceas en el Perú han sido estudiadas por varios autores (Rauh 1958, Britton & Rose 1963, Ritter 1981, Ostolaza Nano 2011), aunque Rauh (1958) y Ritter (1981) son los que aportan una mayor cantidad de datos sobre su distribución geográfica permitiendo elaborar una discusión sobre sus causas. Para la actualización de la nomenclatura de las plantas que intervienen en las comunidades presididas por Cactáceas hemos seguido a Brako & Zarucchi (1993) y la base de datos Tropicos (2012). En particular, las Cactáceas siguen a Hunt (2006) excepto para Corryocactus puquiensis Rauh & Backeb., Echinopsis schoenii (Rauh & Backeb.) Friedrich & G.D. Rowley, Oreocereus piscoensis (Rauh & Backeb.) F. Ritter, O. tacnaensis F. Ritter y Weberbauerocereus torataensis F. Ritter, en que no hemos considerado la sinonimia al tratarse de plantas morfológicamente diferentes y donde a veces su distribución ni siquiera abarca las localidades típicas del taxón sinonimizado. Para la distribución de endemismos hemos consultado a Arakaki et al. (2006).
Para la determinación de las plantas de los muestreos hemos utilizado los herbarios AQP, F, MO, NY, US y USM; los especímenes colectados se conservan en AQP.
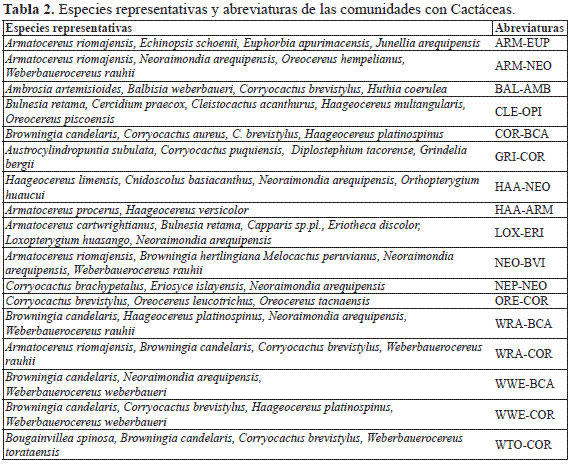
Las comunidades vegetales con Cactáceas estudiadas a lo largo de las vertientes occidentales andinas son las que figuran en la Tabla 2 donde quedan identificadas por sus especies más representativas.
Datos climáticos. Para intentar estudiar la influencia del clima sobre las comunidades de Cactáceas, hemos utilizado parámetros e índices climáticos con valores de temperatura, precipitación y humedad: Precipitación anual en mm (P), precipitación estival (PE), precipitación invernal (PI), evapotranspiración potencial anual (ETP), evapotranspiración potencial estival (ETPE), evapotranspiración potencial invernal (ETPI), humedad estival (HE), humedad invernal (HI), aridez estival (1/HE) y aridez invernal (1/HI) (Tabla 3).
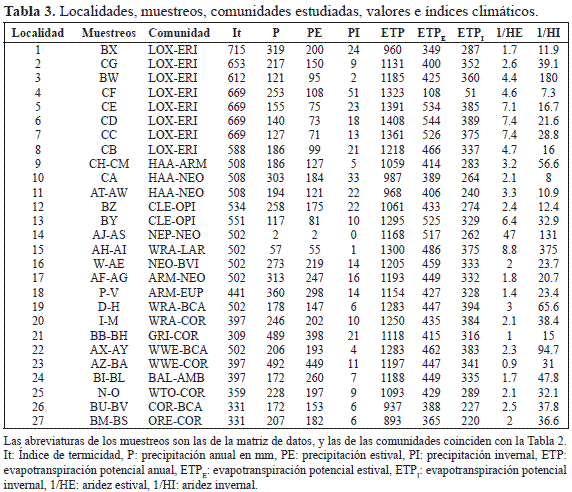
Como índice de temperaturas hemos calculado el Índice de Termicidad (It) con el fin de aplicar el modelo de los "pisos bioclimáticos" de Rivas-Martínez et al. (1999) en localidades próximas o con altitud semejante adonde se han levantado los muestreos (Tabla 3). Los valores de It son coincidentes con cinturas altitudinales y latitudinales de flora y vegetación, y en ellas se pueden seleccionar bioindicadores (Kuentz et al. 2007, Galán de Mera et al. 2010). En el territorio estudiado podemos distinguir 4 pisos bioclimáticos: infratropical (It > 610), termotropical (It = 610-471), mesotropical (It = 470-311) y supratropical (It = 310-171).
Siguiendo el modelo climático de predicción de la vegetación de Galán de Mera & Vicente Orellana (2007) para los valores e índices relativos a la humedad de los territorios (HE, HI), tomamos como meses estivales diciembre, enero, febrero y marzo, y como meses invernales junio, julio, agosto y septiembre. La evapotranspiración potencial (ETP) es muy importante para predecir los tipos de vegetación, ya que está directamente relacionada con la humedad del suelo y la radiación solar. ETP incluye la transpiración de las plantas y la evaporación del suelo (Schultz 1995). Consideramos a la humedad de una localidad como el cociente entre la precipitación y ETP. Sin embargo, como la precipitación en las estaciones meteorológicas del desierto costero es demasiado baja como para que tenga valores significativos, utilizamos los valores inversos de la humedad (aridez), 1/HE y 1/HI.
Los datos meteorológicos para determinar las características climáticas de las distintas localidades han sido tomados de Müller (1982), Rivas-Martínez & Rivas-Saenz (1996-2009) y Senamhi (2012). Sin embargo, siguiendo el sistema de superficies interpoladas de Hijmans et al. (2005), hemos utilizado la base de datos SamSam Water (2012), con la que hemos interpolado los valores de precipitación (P, PE y PI), y evapotranspiración potencial (ETP, ETPE y ETPI). Con ello, podemos conocer los valores e índices relativos a la humedad en el punto exacto de realización de los muestreos.
Análisis estadístico. Se construyó una matriz de 88 muestreos ordenados entre D y CM en el eje X (Tabla 3) y 146 especies en el eje Y que están presentes o ausentes en cada muestreo. Sobre esta matriz hemos utilizado el índice de similitud de Sørensen (Sørensen 1948) y un Análisis de Componentes Principales (ACP) elaborados con el programa informático PAST 1.89 (Hammer et al. 2009).
El índice de similitud de Sørensen produce un dendrograma donde podemos observar la distancia entre diferentes muestreos de vegetación. Con el ACP podemos interpretar los datos climáticos que afectan de una forma u otra a las diferentes comunidades de Cactáceas, usando vectores con los que podemos deducir la influencia de las variables climáticas sobre las localidades seleccionadas.
RESULTADOS
Pisos bioclimáticos. De acuerdo con los valores del Índice de Termicidad (It), las áreas de Perú que se extienden sobre el paralelo 8° Sur pertenecen al piso infratropical (localidades 1-7). El resto de las localidades de la Fig. 1, hasta el departamento de Arequipa, pertenecen al piso termotropical (8-15 y 22), mientras que las comunidades situadas hacia el interior de Arequipa (18, 20, 23 y 24), Moquegua (25) y Tacna (26 y 27) pertenecen al mesotropical, excepto las situadas en las zonas basales de los valles interandinos (16, 17 y 19), que vuelven a ser termotropicales. La localidad 8 queda en el límite entre los pisos infra- y termotropical, y la 21 se encuentra en el piso supratropical.
Dendrograma de Sørensen. El dendrograma de similitud de Sørensen (Fig. 2) muestra dos grandes ramificaciones, A: muestreos que no llevan Corryocactus brevistylus (K. Schum. ex Vaupel) Britton & Rose, ni especies de Browningia Britton & Rose ni de Weberbauerocereus Backeb., y B: muestreos donde estos elementos están presentes.
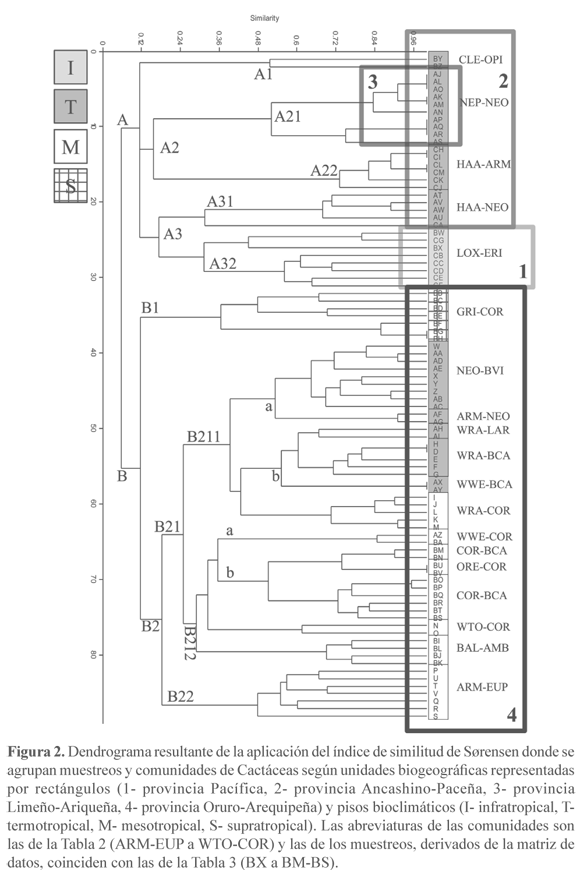
En A, la ramificación A1 son los muestreos termotropicales del departamento de Ica con Bulnesia retama (Gillies ex Hook. & Arn.) Griseb., Cercidium praecox (Ruiz & Pav. ex Hook.) Harms, Cleistocactus acanthurus (Vaupel) D.R. Hunt, Haageocereus multangularis (Haw.) F. Ritter y Oreocereus piscoensis. A2 coincide con los muestreos del piso termotropical con poca riqueza florística, sobre todo de arbustos, y que pertenecen a las comunidades de Eriosyce islayensis (Foerster) Katt. y Neoraimondia arequipensis Backeb. (A21) y a las de Haageocereus versicolor (Werderm. & Backeb.) Backeb. y Armatocereus procerus Rauh & Backeb. (A22)(Fig. 3A); la ramificación A3 corresponde a inventarios termotropicales ricos en arbustos (Grabowskia boerhaaviifolia (L.f.) Schltdl., Trixis cacalioides (Kunth) D. Don.), como ocurre en la comunidad de Haageocereus limensis (Salm-Dyck) F. Ritter y Neoraimondia arequipensis del centro del Perú (A31), y al piso infratropical con comunidades donde están presentes Eriotheca discolor (Kunth) A. Robyns, Loxopterygium huasango Spruce ex Engl. y Neoraimondia arequipensis (A32).

B presenta dos grandes ramificaciones: B1 con la comunidad supratropical de Grindelia bergii (Hieron.) Cabrera y Corryocactus puquiensis, que al llevar plantas que alcanzan una mayor altitud (Austrocylindropuntia subulata (Muehlenpf.) Backeb., Diplostephium tacorense Hieron., Proustia oblongifolia D. Don o Tunilla soehrensii (Britton & Rose) D.R. Hunt & Iliff), muestra una mayor distancia, y B2, con inventarios, la mayoría termo- y mesotropicales, donde están Armatocereus riomajensis Rauh & Backeb., Corryocactus brevistylus, Euphorbia apurimacensis Croizat, Weberbauerocereus rauhii Backeb., W. torataensis y W. weberbaueri (K. Schum. ex Vaupel) Backeb. La división de B2 nos lleva a una gran diversidad de comunidades en el sur del Perú ligada a los valles interandinos de origen volcánico. B21 comparte elementos occidentales como Ambrosia artemisioides Meyen & Walp. ex Meyen, Browningia candelaris (Meyen) Britton & Rose, Corryocactus aureus (Meyen) Hutchison o Haageocereus platinospinus (Werderm. & Backeb.) Backeb. La comunidad interandina de Armatocereus riomajensis y Euphorbia apurimacensis (B22) (Fig. 3B) está compuesta por una flora muy particular donde están presentes los endemismos Echinopsis schoenii y Junellia arequipense (Botta) Botta.
La diferencia entre B211 y B212 la marca la presencia en B211 de Weberbauerocereus rauhii, y la mayor parte de esta ramificación se corresponde con inventarios termotropicales, de tal forma que las comunidades de Browningia candelaris, Corryocactus brevistylus, Weberbauerocereus rauhii, W. weberbaueri y Neoraimondia arequipensis son las más térmicas y de menor altitud en las vertientes occidentales de los Andes de Arequipa. Bajo B211, a y b son muestreos termotropicales y tienen como elemento común a Neoraimondia arequipensis, un bio-indicador termo-infratropical; a, es una rama constituida por la comunidad interandina de Neoraimondia arequipensis y Browningia hertlingiana (Rauh) Buxb., y la comunidad de Armatocereus riomajensis y N. arequipensis, que tienen en común Armatocereus riomajensis, N. arequipensis, Oreocereus hempelianus (Gürke) D.R. Hunt y Weberbauerocereus rauhii; la primera incluye a Melocactus peruvianus Vaupel, un elemento común con las comunidades del centro y del norte. b reúne a las comunidades termotropicales con Weberbauerocereus donde además puede intervenir Larrea divaricata Cav. B212 corresponde a la vegetación mesotropical caracterizada por tres endemismos que se extienden entre los departamentos de Arequipa y Tacna; en los alrededores de Arequipa (a) Weberbauerocereus weberbaueri, en Tacna (b) Oreocereus tacnaensis, y en Moquegua (c) W. torataensis.
ACP y condiciones climáticas. Del ACP de la Fig. 4 deducimos que las localidades se encuentran agrupadas por pisos bioclimáticos y altitudes, y que muestran una mayor o menor lejanía de los vectores 1/HE y 1/HI que significan la aridez estival o invernal. El grupo I son las estaciones infratropicales con mayor índice It, por el contrario, también son las situadas a menor altitud (A); según nos desplazamos hacia los valores positivos del eje 1, pasamos al grupo T (termotropical), M (mesotropical), y S (supratropical), a la vez que se va ascendiendo en altitud.
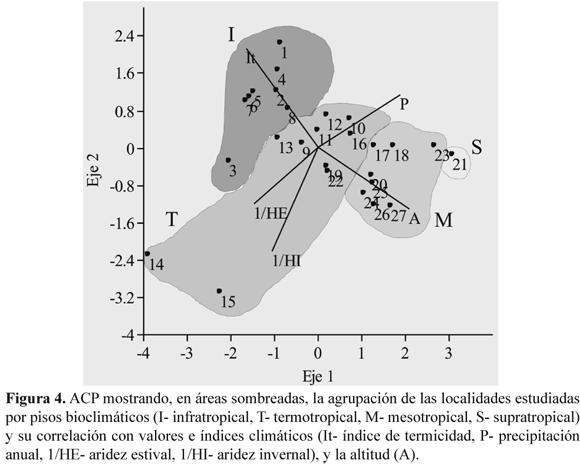
Las localidades 1-12 son las que están más alejadas de la aridez invernal, y conforme los valores del eje 2 son más elevados, la humedad estival (HE) e invernal (HI) se van haciendo mayores debido a la influencia de la corriente El Niño que se pone de manifiesto, sobre todo en 1 (Zarumilla-Tumbes), 2 ( La Brea-Piura) y 4 (Olmos-Lambayeque), con una vegetación formada por Armatocereus cartwrightianus (Britton & Rose) Backeb. ex A.W. Hill, Bulnesia retama, Bursera graveolens (Kunth) Triana & Planch., Capparis L. sp.pl., Cercidium praecox, Eriotheca discolor, Loxopterygium huasango y Neoraimondia arequipensis. Las localidades 9-13 están situadas por debajo del paralelo 8° Sur; aquí la humedad invernal (HI) también es importante ya que procede de las nieblas aportadas por la corriente de Humboldt, y la humedad estival (HE) es debida a las escasas precipitaciones andinas, aunque en 12 (Pisco-Ica) y 13 (Ingenio-Ica) es algo mayor, coincidiendo con la presencia de Bulnesia retama y Cercidium praecox. Todas estas localidades tienen en común a Cnidoscolus basiacanthus (Pax & K. Hoffm.) J.F. Macbr., Espostoa melanostele (Vaupel) Borg, Gravouwskia boerhaaviifolia, Neoraimondia arequipensis y Orthopterygium huaucui (A. Gray) Hemsl. En 9 (Chamana, Ancash) hay una mayor aridez ya que los muestreos pertenecen a las partes secas basales del río Fortaleza, destacando Haageocereus versicolor y Armatocereus procerus en su composición florística.
Las localidades de los grupos M y S presentan sobre todo influencia de la humedad estival (HE), aunque en menor grado que 1 (Zarumilla-Tumbes). En M, cuanto más nos adentramos en los valores positivos del eje 2 la aridez estival (1/HE) se va haciendo menor, y al desplazarnos hacia la derecha en el eje 1 HI tiene una cierta importancia, pues 16 (Velinga-Arequipa), 17 (Cotahuasi-Arequipa) y 18 (Soro-Arequipa) pertenecen a valles interandinos, más próximos a las vertientes orientales de los Andes. 19 (Huacán-Arequipa) y 22 (Uchumayo-Arequipa) presentan mayor aridez estival (1/HE) e invernal (1/HI) puesto que están al finalizar el desierto abiótico en las partes más bajas de los Andes, y esta aridez se extiende hasta Atico-Arequipa (14) y Aplao-Arequipa (15). Atico está en la costa y Aplao igualmente al inicio de las estribaciones andinas con la originalidad de la presencia de Larrea divaricata.
En los grupos M, S y las localidades 15-17, y 19 y 22 de T encontramos vegetación donde interviene Corryocactus brevistylus en el piso meso- y supratropical, Neoraimondia arequipensis, en el termotropical, y en general, Browningia candelaris, Weberbauerocereus rauhii, W. torataensis y W. weberbaueri. En 14, N. arequipensis se acompaña de Eriosyce islayensis, con una aridez estival e invernal muy elevadas, y en 16 podemos destacar a Browningia hertlingiana.
DISCUSIÓN
Comunidades de Cactáceas y territorios biogeográficos. Los grupos A y B del dendrograma (Fig. 2) marcan dos grandes territorios, A, donde Corryocactus brevistylus está ausente, y B, donde C. brevistylus está presente en todas las comunidades limitando la vegetación de Cactáceas hasta unos 3500 m de altitud dentro de la provincia Oruro-Arequipeña (Galán de Mera et al. 2009), que se extiende también por las laderas occidentales de los Andes chilenos adyacentes (Luebert & Gajardo 2005). En B, el paquete de plantas que acompañan a C. brevistylus, en especial Armatocereus riomajensis, Browningia candelaris, Weberbauerocereus rauhii y W. weberbaueri, son sustituidas por las de la rama A: Armatocereus procerus, Cnidoscolus basiacanthus, Espostoa melanostele u Orthopterygium huaucui, de la provincia Ancashino-Paceña, y éstas por Armatocereus cartwrightianus, Bursera graveolens, Capparis sp. pl., Cercidium praecox o Loxopterygium huasango de la provincia Pacífica (A32) (Galán de Mera et al. 2002). Estos bosques y arbustedas de la provincia Pacífica se encuentran relacionados con los de Ecuador (Linares-Palomino 2006); así los trazos de A. cartwrightianus que muestran Loaiza S. & Morrone (2011) se continúan desde Perú indicando una distribución andina occidental, y la extensión de la provincia biogeográfica Pacífica hacia Ecuador, coincidiendo con lo que estos autores llaman "trazo generalizado costero seco".
En todos los Andes occidentales del Perú, desde las menores altitudes hasta unos 2100 m, se encuentra Neoraimondia arequipensis, que al ser un endemismo peruano (Brako & Zarucchi 1993) de los pisos bioclimáticos infra- y termotropical (Galán de Mera et al. 2010) que se extiende de norte a sur, es quien mantiene la originalidad de las comunidades de Cactáceas en el territorio. Aunque Rauh (1958) y Ritter (1981) llevan esta especie hasta Chile, no está citada en su flora (Marticorena & Quezada 1985). Las comunidades con Eriosyce islayensis de la rama A21 pertenecen a la provincia costera Limeño-Ariqueña.
En análisis de la distribución de Cactáceas de Bolivia que presenta Navarro (1996), que también utiliza el modelo de los pisos bioclimáticos y define unidades biogeográficas, las especies enumeradas no se encuentran en nuestro territorio estudiado, pues aunque el altiplano peruano limita con el occidente boliviano, la cordillera establece una barrera biogeográfica entre los valles de ambos lados. La vegetación de los valles suroccidentales de Bolivia está relacionada con la del Chaco (Navarro & Maldonado 2005); la presencia de Armatocereus Backeb. y Browningia Britton & Rose en los Andes áridos occidentales peruanos tal vez tenga su origen en los bosques secos del norte del Perú (Loaiza S. & Morrone 2011), lo que hace que sus comunidades sean muy diferentes florísticamente a las bolivianas.
En el mapa de la Fig. 1 podemos dibujar círculos de distribución de Cactáceas que coinciden con estas unidades biogeográficas: A- Armatocereus cartwrightianus, Austrocylindropuntia pachypus (K. Schum.) Backeb., Espostoa lanata (Kunth) Britton & Rose (provincia Pacífica); B- Armatocereus procerus, Espostoa melanostele, Mila Britton & Rose sp. pl. (provincia Ancashino-Paceña); C- Armatocereus riomajensis, Browningia candelaris, Corryocactus brevistylus, Weberbauerocereus rauhii, W. weberbaueri (provincia Oruro-Arequipeña), y D- Corryocactus brachypetalus (Vaupel) Britton & Rose, Eriosyce islayensis y Haageocereus decumbens (Vaupel) Backeb. (provincia Limeño-Ariqueña).
Recientemente Ostolaza Nano (2011), divide N. arequipensis en tres subespecies, cuya distribución vuelve a coincidir con estas unidades biogeográficas (Fig. 1): N. arequipensis subsp. arequipensis, de la provincia Oruro-Arequipeña y Limeño-Ariqueña (C y D respectivamente), N. arequipensis subsp. gigantea (Werderm. & Backeb.) Ostolaza, de la provincia Pacífica (A), y N. arequipensis subsp. roseiflora (Werderm. & Backeb.) Ostolaza, de la provincia Ancashino-Paceña (B).
Distribución de comunidades de Cactáceas de Norte a Sur. Del ACP (Fig. 4) deducimos cómo las localidades 1-8 son las más influidas por la humedad estival (HE) e invernal (HI) relacionadas con la corriente de El Niño, cuya humedad persiste durante más de un año (Huertas Vallejos 2009, Fagan 2010). Los bosques xerófilos de este territorio, donde intervienen Cactáceas, son debidos a la humedad aportada por las lluvias procedentes del Océano Pacífico y a la aportada desde la Cuenca Amazónica (Koepcke 1961, Ferreyra 1983). Como ya indicaron Galán de Mera et al. (1997), hacia el sur va disminuyendo progresivamente la riqueza de árboles y arbustos hasta los 8° Sur (latitud de la Cordillera Blanca con el Huascarán, 6768 m). La falta de lluvias occidentales, como consecuencia de la sombra de lluvias de los Andes y de la corriente de Humboldt, disminuye la presencia de arbustos en el círculo de Cactáceas B y C de la Fig. 1. No obstante, en el valle del río Ingenio (Ica), las comunidades con Neoraimondia arequipensis albergan, de forma relictual, Bulnesia retama y Cercidium praecox (Weberbauer 1945, localidad 13 del ACP, muestreo BY del dendrograma), lo que coincide con los lapsos de clima más húmedo detectados hace más de 5000 años (Arntz & Fahrbach 1996) y, posteriormente, fuertes episodios de El Niño, que vienen a producir una gran proliferación de la vegetación (Beresford-Jones et al. 2009).
Distribución de las comunidades de Cactáceas de Oeste a Este en el Sur del Perú. Recientemente han sido registradas en el occidente del sur del Perú algunas plantas, como Browningia hertlingiana [= B. viridis (Rauh & Backeb.) Buxb.] y Euphorbia apurimacensis, con una mayor distribución en el oriente peruano (Ritter 1981, Linares Perea 2005, Linares Perea et al. 2009). A ellas hay que añadir el nuevo registro de Larrea divaricata en los alrededores de Aplao (localidad 15 del ACP, muestreos BI y BH del dendrograma)(Fig. 2).
La presencia de Larrea divaricata en el seno de una comunidad de Weberbauerocereus rauhii y Browningia candelaris, refleja un clima anterior más húmedo y cálido semejante al de las sierras de Mendoza, Córdoba y La Rioja en Argentina (Hunziker 1952, Eskuche & Landolt 1986). En la actualidad, L. divaricata sobrevive en áreas con afloramientos pre-cámbricos y avalanchas volcánicas procedentes de erupciones del Terciario inferior, época que precede a la creciente aridez de las vertientes occidentales de los Andes a finales del Plioceno (Hartley 2003, Lamb & Davis 2003).
Las comunidades de Armatocereus riomajensis, Echinopsis schoenii y Euphorbia apurimacensis (localidad 18 del ACP, muestreos P-V del dendrograma) y de Neoraimondia arequipensis y Browningia hertlingiana (localidad 16 del ACP, muestreos W-AD del dendrograma) están situadas en valles interandinos transversales en el departamento de Arequipa, y correlacionadas en el ACP con la humedad estival de las lluvias andinas. Están asentadas sobre materiales volcánicos miocenos cuyo areal registró una creciente aridez por la elevación de la Cordillera y la formación de valles profundos debidos al vulcanismo terciario y cuaternario (Lissón 1980). Las localidades 18 (Soro-Arequipa) y 23 (Arequipa)(mesotropicales), y 21 (Chivay-Arequipa) (supratropical) y 24 (Chiguata-Arequipa)(mesotropical) son las más favorecidas por la humedad estival andina (HE), aunque también adquiere una cierta importancia la humedad invernal (HI) debido a las depresiones aisladas que se desprenden de las borrascas subantárticas entre julio y agosto (Mestanza Hernández 2005).
Comunidades de Cactáceas y uso del territorio. La correlación entre la humedad estival (HE) e invernal (HI), y las comunidades de Cactáceas occidentales en el sur del Perú se materializa con la presencia de grupos agro-pastoralistas desde hace unos 5000 años (Grosjean et al. 2005). Éstos usaban los pastos anuales que crecen en las comunidades de Cactáceas ejerciendo la trashumancia desde las partes altas (Rostworowski 2009), en muchas ocasiones favorecida por episodios de El Niño, coincidentes con sequías en las altitudes andinas y una mayor humedad estival en las partes bajas; incluso, se han encontrado espinas de cactus en zonas de combustión de los asentamientos humanos del piso termotropical, que pudieron ser usadas como elementos de las vestimentas u otros enseres domésticos (Núñez et al. 2005). Tal vez en épocas muy arcaicas comience a establecerse entre los pobladores el uso del máximo de pisos ecológicos que perdura hasta la actualidad (Murra 2009), con diferentes tipos de cultivos entre el piso termotropical y orotropical, como una originalidad de las culturas Inca y pre-Inca consecuente con los cambios climáticos.
Las fotografías de las Fig. 5A y Fig. 5B reflejan la importancia que debió tener la humedad estival (HE) en las localidades 16 (Velinga-Arequipa) y 20 (Huacán-Arequipa) durante la cultura Wari (s. VIII-X) que, con un clima aún más húmedo que el actual (Ortloff & Kolata 1993), cultivaban en andenes donde hoy la aridez es evidente, al estar invadidos por los elementos de las comunidades de Cactáceas.

AGRADECIMIENTOS
Este trabajo ha sido realizado con el apoyo de la Cátedra de Desarrollo Sostenible y Medio Ambiente de la Universidad San Pablo-CEU-Grupo Santander (2009-2011). Nuestro agradecimiento a la Universidad Nacional Mayor de San Marcos (Lima, Perú) y a la Universidad Nacional de San Agustín (Arequipa, Perú) por su apoyo constante. Gracias a los dos revisores anónimos cuyos comentarios han mejorado el manuscrito.
LITERATURA CITADA
1. Arakaki, M., C. Ostolaza, F. Cáceres & J. Roque. 2006. Cactaceae endémicas del Perú. Revista Peruana de Biología 13(2): 193-219. [ Links ]
2. Arntz, W.E. & E. Fahrbach. 1996. El Niño. Experimento climático de la naturaleza. Fondo de Cultura Económica, México D.F. 309 pp. [ Links ]
3. Beresford-Jones, D., H. Lewis & S. Boreham. 2009. Linking cultural and environmental change in Peruvian prehistory: Geomorphological survey of the Samaca Basin, Lower Ica Valley, Peru. Catena 78: 234-249. [ Links ]
4. Brako, L. & J.L. Zarucchi. 1993. Catálogo de las Angiospermas y Gimnospermas del Perú. Missouri Botanical Garden, San Luis. 1286 pp. [ Links ]
5. Braun-Blanquet, J. 1932. Plant Sociology. The study of plant communities. Koeltz Scientific Books, Koenigstein. 439 pp. [ Links ]
6. Britton, N.L. & J.N. Rose. 1963. The Cactaceae. Reimpresión de la 2a edición, 1937. Dover, Nueva York. 4 vol. [ Links ]
7. Eskuche, U. & E. Landolt. 1986. Contribution to the knowledge of flora and vegetation of northern Argentina. Veröffentlichungen des Geobotanischen Institutes der ETH, Stiftung Rübel Zürich 91: 14-140. [ Links ]
8. Fagan, B. 2009. El gran calentamiento. Gedisa, Barcelona. 350 pp. [ Links ]
9. Fagan, B. 2010. La corriente de El Niño y el destino de las civilizaciones. Inundaciones, hambrunas y emperadores. Gedisa, Barcelona. 347 pp. [ Links ]
10. Ferreyra, R. 1983. Los tipos de vegetación de la costa peruana. Anales del Jardín Botánico de Madrid 40: 241-256. [ Links ]
11. Galán de Mera, A., S. Baldeón, H. Beltrán, M. Benavente & J. Gómez. 2004. Datos sobre la vegetación del centro del Perú. Acta Botanica Malacitana 29: 89-115. [ Links ]
12. Galán de Mera, A. & J. Gómez Carrión. 2001. Las comunidades con Cactáceas del sur del Perú. Nuevos datos sobre la alianza Corryocaction brevistyli. Acta Botanica Malacitana 26: 240-246. [ Links ]
13. Galán de Mera, A., E. Linares Perea, J. Campos de la Cruz & J.A. Vicente Orellana. 2009. Nuevas observaciones sobre la vegetación del sur del Perú. Del Desierto Pacífico al Altiplano. Acta Botanica Malacitana 34: 107-144. [ Links ]
14. Galán de Mera, A., E. Linares Perea, C. Trujillo Vera & F. Villasante Benavides. 2010. Termoclima y humedad en el sur del Perú. Bioclimatología y bioindicadores en el departamento de Arequipa. Zonas Áridas 14: 71-82. [ Links ]
15. Galán de Mera, A., M.V. Rosa & C. Cáceres. 2002. Una aproximación sintaxonómica sobre la vegetación del Perú. Clases, órdenes y alianzas. Acta Botanica Malacitana 27: 75-103. [ Links ]
16. Galán de Mera, A. & J.A. Vicente Orellana. 1996. Las comunidades con Corryocactus brevistylus del sur del Perú. Phytologia 80: 40-47. [ Links ]
17. Galán de Mera, A. & J.A. Vicente Orellana. 2007. Cronosequences of vegetation- a bioclimatic theory for interpreting the patterns of relic vegetation types. Phytocoenologia 37: 471-494. [ Links ]
18. Galán de Mera, A., J.A. Vicente Orellana, J.A. Lucas García & A. Probanza Lobo. 1997. Phytogeographical sectoring of the Peruvian coast. Global Ecology and Biogeography Letters 6: 349-367. [ Links ]
19. Garzione, C.N., G.D. Hoke, J.C. Libarkin, S. Withers, B. MacFadden, J. Eiler, P. Ghosh & A. Mulch. 2008. Rise of the Andes. Science 320: 1304-1308. [ Links ]
20. Graham, A. 2009. The Andes: A Geological Overview from a Biological Perspective. Annals of the Missouri Botanical Garden 96: 371-385. [ Links ]
21. Grosjean, M., L. Núñez & I. Cartajena. 2005. Cultural response to climate change in the Atacama Desert. En: Smith, M. & P. Hesse (eds.). 23° South: The Archaeology and Environmental History of the Southern Desert: 156-171. National Museum of Australia. Camberra. [ Links ]
22. Hammer, Ø., D.A.T. Harper & P.D. Ryan. 2009. PAST- Paleontological Statistics, ver.1.89. University of Oslo, Oslo. 92 pp. [ Links ]
23. Hartley, A.J. 2003. Andean uplift and climate change. Quarterly Journal of the Geological Society of London 62(2): 497-500. [ Links ]
24. Hershkovitz, M.A. & E.A. Zimmer. 1997. On the evolutionary origins of the cacti. Taxon 46: 217-232. [ Links ]
25. Hijmans, R.J., S.E. Cameron, J.L. Parra, P.G. Jones & A. Jarvis. 2005. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25: 1965-1978. [ Links ]
26. Holmgren, C.A., J.L. Betancourt, K.A. Rylander, J. Roque, O. Tovar, H. Zeballos & E. Linares. 2001. Holocene Vegetation History from Fossil Rodent Middens near Arequipa, Peru. Quaternary Research 56: 242-251. [ Links ]
27. Huertas Vallejos, L. 2009. Injurias del tiempo. Desastres naturales en la historia del Perú. Universidad Ricardo Palma, Lima. 403 pp. [ Links ]
28. Hunt, D. 2006. The New Cactus Lexicon. Dh Books, Milborne Port. 373 pp. [ Links ]
29. Hunziker, J.H. 1952. Las comunidades vegetales de la cordillera de La Rioja. Revista de Investigación Agrícola 6(2): 167-181. [ Links ]
30. Ibaraki, M. 1997. Closing of the Central American Seaway and Neogene coastal upwelling along the Pacific coast of South America. Tectophysics 281: 99-104. [ Links ]
31. Koepcke, H-W. 1961. Synökologische Studien an der Westseite der peruanischen Anden. Ferd. Dümmlers, Bonn. 320 pp. [ Links ]
32. Kuentz, A., A. Galán de Mera, M-P. Ledru & J-C. Thouret. 2007. Phytogeographical data and modern pollen rain of the puna belt in southern Peru (Nevado Coropuna, Western Cordillera). Journal of Biogeography 34: 1762-1776. [ Links ]
33. Lamb, S. & P. Davis. 2003. Cenozoic climate change as a possible cause for the rise of the Andes. Nature 425: 792-797. [ Links ]
34. Linares-Palomino, R. 2006. Phytogeography and Floristics of Seasonally Dry Tropical Forests in Peru. En: Pennington, R.T. & J.A. Ratter (eds.). Neotropical Savannas and Seasonally Dry Forests: Plant Diversity, Biogeography, and Conservation: 257-279. CRC Press. Boca Raton. [ Links ]
35. Linares Perea, E. 2005. Cotahuasi, Perú. Un paraíso de Cactus. Quepo 19: 92-97. [ Links ]
36. Linares Perea, E., J. Campos de la Cruz, W. Nauray Huari, J.A. Vicente Orellana & A. Galán de Mera. 2010. Nuevas adiciones a la flora del Perú, V. Arnaldoa 17: 99-112. [ Links ]
37. Linares Perea, E., A. Galán de Mera, J. Campos de la Cruz, W. Nauray Huary & J.A. Vicente Orellana. 2009. Nuevas adiciones a la flora del Perú, IV. Arnaldoa 16: 75-79. [ Links ]
38. Lissón, G. 1980. Cómo se generó el suelo peruano. Contribución a la paleogeografía del Perú y las causas del relieve actual. En: Samame Boggio, M. (ed.). El Perú Minero, III. Geología: 61-82. Ingemmet. Lima. [ Links ]
39. Livermore, R., A. Nankivell, G. Eagles & P. Morris. 2005. Paleogene opening of Drake Passage. Earth and Planetary Science Letters 236: 459-470. [ Links ]
40. Loaiza S., C.R. & J.J. Morrone. 2011. Análisis panbiogeográfico de algunas Cactaceae del Ecuador. Gayana Botánica 68(2): 220-225. [ Links ]
41. Luebert, F. & R. Gajardo. 2005. Vegetación alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetación de la Puna meridional. Phytocoenologia 35: 79-128. [ Links ]
42. Marticorena, C. & M. Quezada. 1985. Catálogo de la flora vascular de Chile. Gayana Botánica 42: 5-157. [ Links ]
43. Mestanza Hernández, F. 2005. Episodios sinópticos de la nevada en la Sierra Sur-Año 2004. Boletín de la Sociedad Geográfica de Lima 118: 67-78. [ Links ]
44. Müller, M.J. 1982. Selected climatic data for a global set of standard stations for vegetation science. Dr. W. Junk, La Haya, Boston, Londres. 306 pp. [ Links ]
45. Murra, J.V. 2009. El mundo andino. Población, medio ambiente y economía. Instituto de Estudios Peruanos-Pontificia Universidad Católica del Perú, Lima. 511 pp. [ Links ]
46. Navarro, G. 1996. Catálogo ecológico preliminar de las cactáceas de Bolivia. Lazaroa 17: 33-84. [ Links ]
47. Navarro, G. & M. Maldonado. 2005. Geografía ecológica de Bolivia. Vegetación y ambientes acuáticos. Fundación Simón Patiño, Santa Cruz. 719 pp. [ Links ]
48. Núñez, L., M. Grosjean & I. Cartajena. 2005. Ocupaciones humanas y paleoambientes en la puna de Atacama. Universidad Católica de Chile-Taraxacum, San Pedro de Atacama. 480 pp. [ Links ]
49. Ortloff, CH. & A.L. Kolata. 1993. Climate and collapse: Agro-ecological perspectives on the decline of the Tiwanaku state. Journal of Archaeological Science 20: 195-221. [ Links ]
50. Ostolaza Nano, C. 2011. 101 cactus del Perú. Ministerio del Ambiente, Lima. 253 pp. [ Links ]
51. Rauh, W. 1958. Beitrag zur Kenntnis der Peruanischen Kakteenvegetation. Springer, Heidelberg. 542 pp. [ Links ]
52. Ritter, F. 1981. Kakteen in Südamerika, 4. Friedrich Ritter Selbstverlag, Spangenberg. 1692 pp. [ Links ]
53. Rivas-Martínez, S. & S. Rivas-Saenz. 1996-2009. Worldwide Bioclimatic Classification System (http://www.globalbioclimatics.org). Madrid. [ Links ]
54. Rivas-Martínez, S., D. Sánchez Mata & M. Costa. 1999. North American Boreal and Western temperate Forest Vegetation (Syntaxonomical sinopsis of the potential natural plant communities of North America, II). Itinera Geobotanica 12: 5-316. [ Links ]
55. Rostworowski, M. 2009. Historia del Tahuantinsuyu. Instituto de Estudios Peruanos, Lima. 359 pp. [ Links ]
56. SamSam Water Foundation. 2012. SamSam Water Climate Tool. (http://www.samsamwater.com). AA Zaandam, Holanda. [ Links ]
57. Schultz, J. 1995. The ecozones of the world. The ecological divisions of the geosphere. Springer, Berlin. 449 pp. [ Links ]
58. Schwarzer, C., F. Cáceres Huamaní, A. Cano, M.I. La Torre & M. Weigend. 2010. 400 years for long-distance dispersal and divergence in the northern Atacama desert- Insights from the Huaynaputina pumice slopes of Moquegua, Peru. Journal of Arid Environments 74: 1540-1551. [ Links ]
59. Senamhi. 2012. Servicio Nacional de Meteorología e Hidrología del Perú (http://www.senamhi.gob.pe). Lima. [ Links ]
60. Solbrig, O. 1976. The origin and floristic affinities of the South American temperate desert and semiarid regions. En: Goodall, D. (ed.). Evolution of desert biota: 7-50. University of Texas Press. Austin. [ Links ]
61. Sørensen, T. 1948. A method of stabilizing groups of equivalent amplitude in plant sociology based on the similarity of species content and its application to analysis of the vegetation on Danish commons. Biologiske Skrifter Kongeline Danske Videnskabernes Selskab 5: 1-34. [ Links ]
62. Tropicos. 2012. http://www.tropicos.org. Missouri Botanical Garden, San Luis. [ Links ]
63. Villagrán, C. & L.F. Hinojosa. 2005. Esquema biogeográfico de Chile. En: Llorente Bousquets, J. & J.J. Morrone (eds.). Regionalización biogeográfica en Iberoamérica y tópicos afines: 551-577. Universidad Nacional Autónoma de México. México DF. [ Links ]
64. Weberbauer, A. 1912. Pflanzengeographische Studien im südlichen Peru. Botanische Jahrbücher für Systematik 107: 27-46. [ Links ]
65. Weberbauer, A. 1945. El mundo vegetal de los Andes Peruanos (Estudio fitogeográfico). Ministerio de Agricultura, Lima. 777 pp. [ Links ]
