











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Para muchos invertebrados la entrada de materia orgánica a los cuerpos de agua constituye una importante fuente de alimento y hábitat (Quinn, et al., 2000). La hojarasca que llega a los arroyos como materia orgánica en partículas gruesas (MOPG) genera una dinámica permanente en los procesos de producción, permitiendo que las comunidades biológicas se estructuren en ríos de pequeño orden (Quinn, et al., 2000; Rincón, et al., 2005; Guevara, et al., 2009). Tan pronto las hojas ingresan al sistema acuático, se inicia el proceso de descomposición por la acción de factores mecánicos, fisicoquímicos y biológicos que permiten que el material depositado sea aprovechado en forma inmediata (Eyes-Escalante, et al., 2012). Entre los factores biológicos se destaca el papel de los macroinvertebrados en la transformación de la materia orgánica, ya que aceleran el proceso de fragmentación y descomposición (Quinn, et al., 2000; Guevara, et al., 2009).
La colonización de la hojarasca por macroinvertebrados acuáticos es un proceso de sucesión ecológica o degradativa en el que se da un reemplazo progresivo de una comunidad por otra, lo que implica no solo el cambio en la composición de la biota sino también de su biomasa y de las condiciones ambientales (Brower & Zar, 1984). Este reemplazo progresivo facilita el flujo de nutrientes a medida que la MOPG es metabolizada y mineralizada (Begon, et al., 2000). Además de la acción de los macroinvertebrados en la degradación de la hojarasca, existe una combinación de factores que contribuyen a su dinámica en los sistemas acuáticos, entre los que se encuentran las propiedades fisicoquímicas del agua, el uso del suelo y la composición química de las hojas (Medina-Villar, et al., 2015). Algunos compuestos secundarios de las hojas pueden ser tóxicos, interferir con la digestión o dar un sabor amargo, y actúan como un impedimento en el momento de ser consumidos o colonizados por diversos organismos, lo que puede afectar la densidad y diversidad de los macroinvertebrados (Graça, 2001; Medina-Villar, et al., 2015).
Además de las características de la hoja, los cambios en la vegetación ribereña por actividades antropogénicas tienen efecto en la cantidad y la calidad de la materia orgánica que entra a los cuerpos de agua, así como en el tiempo requerido para su fragmentación (Golladay, et al., 1987; Webster, et al., 1990, Tuchman & King, 1993; Delong & Brusven, 1994). Las alteraciones en los aportes de tipo alóctono pueden conllevar un impacto en las comunidades acuáticas, especialmente en quebradas donde la principal fuente de energía se basa en la materia orgánica de origen terrestre (Cummins, et al., 1989; Graça, 1993). Este es el caso de la minería, la cual es una de las perturbaciones antropogénicas que causa afectaciones tanto en los cuerpos de agua como en sus zonas ribereñas, pues la descarga de sedimentos como resultado de esta actividad acelera la erosión y altera el tipo y la abundancia relativa de los microhábitats acuáticos (Hammond, et al., 2013).
Una forma de evaluar los efectos de estas actividades antropogénicas sobre el recurso hídrico es a través de la colonización de macroinvertebrados acuáticos con el uso de sustratos artificiales o naturales, que incluyen desde tejas de barro y bandejas de roca y sedimento hasta hojas artificiales o naturales en bolsas de malla plástica (Carvalho, et al., 2008). A pesar de los esfuerzos de distintos investigadores para determinar el papel de los macroinvertebrados en la descomposición de la MOPG, los estudios en el trópico siguen siendo pocos comparados con los realizados en Norte América y Europa (Marchant, 1989; Cortés, et al., 1997; Quinn, et al., 2000; Graça, 2001; Waters, et al., 2005). En Colombia los trabajos recientes de diversos investigadores en varias regiones (Guevara, et al., 2008; González, 2009; Eyes-Escalante, et al., 2012; Meza-Salazar, et al., 2012; Gutiérrez-López, et al., 2016), destacan el papel ecológico de los macroinvertebrados acuáticos en la colonización, la descomposición de la hojarasca, el reciclaje de nutrientes, y la transferencia de energía a través de la cadena trófica, así como su valor de bioindicadores de la calidad del agua (Zúñiga-Céspedes, et al., 2018).
En el departamento de Caldas se conocen pocos trabajos relacionados con la descomposición de hojarasca por macroinvertebrados acuáticos. Hasta ahora las investigaciones se han enfocado en la cuenca del río Chinchiná (Meza-Salazar, et al., 2012; Gutiérrez-López, et al., 2016; Rubio-M., et al., 2016), por lo que es preciso profundizar en el tema debido a la importancia de esta cuenca para la región, ya que ofrece muchos bienes y servicios ambientales para el desarrollo social y económico de la subregión Centro-Sur (Secretaria Técnica de los Pactos, 2012). El objetivo de este estudio fue evaluar la colonización de macroinvertebrados acuáticos en hojas individualizadas y mixtas (1:1) de Tibouchina lepidota y Eucalyptus sp. en la quebrada La Elvira, la cual ha sido afectada por la actividad minera aurífera en Caldas. La hipótesis propuesta es que las actividades resultantes de la minería ejercen un efecto negativo sobre la colonización de macroinvertebrados acuáticos debido a las alteraciones estructurales (pérdida de vegetación ribereña), físicas y químicas en la quebrada.
Materiales y métodos
Área de estudio
La quebrada La Elvira se localiza en el municipio de Manizales (Caldas, Colombia) y forma parte de la cuenca del río Chinchiná; allí se seleccionaron dos estaciones de muestreo, una denominada referencia y la otra, de minería. La estación de referencia se ubica a 2.766 m de altitud (05°03' 10,9'' N, 75°24' 33,6'' O), presenta vegetación ribereña compuesta principalmente por las familias Asteraceae, Urticaceae y Solanaceae y no ha sufrido impacto antropogénico evidente. En la misma quebrada, aguas abajo, se localiza la estación de minería, a 2.725 m de altitud (05°03' 4,4'' N, 75°24' 33,1'' O), la cual presenta efectos visibles de las actividades de minería aurífera y una zona ribereña donde predominan pastos como Pennisetum clandestinum y algunas herbáceas como Erigeron bonariensis, Hypochaeris radicata y Taraxacum officinale.
Variables ambientales y fisicoquímicas
En cada estación se determinó la velocidad del agua, el ancho y profundidad del cauce y la vegetación ribereña. Se registraron algunas variables fisicoquímicas in situ como el pH, la temperatura del agua, el oxígeno disuelto y el porcentaje de saturación de oxígeno disuelto con un equipo multiparamétrico (Lutron™). Además, se tomaron muestras puntuales de agua para el cálculo de variables fisicoquímicas asociadas con procesos mineros y los efectos potenciales sobre la integridad del recurso hídrico. Los análisis se hicieron en el laboratorio certificado ACUATEST S.A. (Manizales, Colombia).
Colonización de macroinvertebrados acuáticos
Se emplearon 192 bolsas plásticas (litterbags de 15 x 15 cm y ojo de malla de 6 mm), las cuales contenían 5 g de hojas secas senescentes de Eucalyptus sp. (planta exótica), Tibouchina lepidota (Bonpl.) Baill. (planta nativa) y una mezcla (1:1) de las dos especies para determinar si un recurso heterogéneo puede ser más atractivo para los invertebrados. En cada punto de muestreo (en adelante referencia y minería) se ubicaron 96 paquetes con hojas (3 tratamientos x 8 eventos de muestreo x 4 réplicas), dispuestos de forma alterna: Eucalyptus sp., T. lepidota y mixto (Eucalyptus sp. + T. lepidota). Se retiraron cuatro réplicas de cada tratamiento en tiempos predeterminados (3, 7, 15, 30, 45, 60, 70, 90 días). En cada evento de muestreo las bolsas retiradas se introdujeron en bolsas plásticas de cierre hermético con alcohol al 96 % y se transportaron al Laboratorio de Ecología de la Universidad de Caldas. Los organismos se separaron cuidadosamente de las hojas mediante lavado manual en tamices (250-150 ^m) y, posteriormente, se identificaron hasta el nivel de género con las claves y descripciones taxonómicas de Merritt & Cummins (1996), Roldán (1996), Domínguez, et al. (2006) y, Domínguez & Fernández (2009). Las hojas se secaron en un horno a 70 °C durante 48 horas, y ya secas se pesaron en una balanza electrónica de precisión (Pioneer de Ohaus™; 0,0001 g).
Análisis de datos
Para el cálculo de las tasas de descomposición de los diferentes tratamientos (Eucalyptus sp., T. lepidota, mixto), se empleó la ecuación de Olsen (1963):
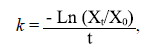
donde k es la constante de descomposición (día1), Xt es el peso de la hojarasca en un tiempo dado (g), X0 es el peso inicial de la hojarasca en el tiempo cero (g) y t, el tiempo expresado en días.
La densidad de macroinvertebrados (ind./m2) se estimó a partir de los organismos recolectados en las bolsas de hojarasca en cada evento de muestreo. Para comparar la riqueza y la densidad de los macroinvertebrados y el porcentaje de remanencia de masa seca entre tratamientos, se realizó un MANOVA no paramétrico con el programa RStudio, versión 0.96.122 (RStudio, 2012). La diversidad se estimó con base en el número efectivo de morfotipos o diversidad orden q (qD) (Chao & Jost, 2012):
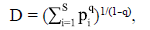
donde pi es la abundancia relativa (abundancia proporcional) de la i-ésima especie, S es el número de especies y q el orden de la diversidad. Cuando q equivale a 0, se obtiene la riqueza, la cual está definida por la incidencia de los morfotipos raros en el muestreo (singletons y doubletons). Cuando q ≈ 1, se obtiene la diversidad típica, que equivale a la exponencial del índice de entropía de Shannon y no presenta sesgo por los morfotipos raros o abundantes del muestreo. Por último, cuando q es igual a 2, el valor de diversidad indicará el número efectivo de morfotipos más abundante en el muestreo y equivale al inverso del índice de entropía de Simpson (Moreno, et al., 2011). La comparación de la diversidad (qD) se realizó bajo la misma completitud o cobertura de muestreo (Ĉm), la cual indica la proporción de la comunidad que está representada en el número de especies (o morfotipos) capturadas y varía entre 0 (baja completitud) y 100 % (alta completitud) (Chao & Jost, 2012).
Para el análisis de la sucesión degradativa se aplicó la fórmula propuesta por Brower & Zar (1984): RNj=Nj/ΣN, donde RNj es la densidad relativa de una especie en un periodo sucesional j (día de muestreo), Nj es la densidad de la especie en un periodo j y ΣN es la suma de las densidades de las especies en todos los días de muestreo; con estos valores se llevó a cabo un ANOVA multifactorial sin interacción, se realizó la prueba de Fisher y, posteriormente, gráficas para evidenciar dichas respuestas.
Resultados
Variables ambientales y fisicoquímicas
Los resultados de las variables ambientales y fisicoquímicas fueron contrastantes entre las zonas de referencia y minería. El ancho de la vegetación ribereña fue mayor en la de referencia que en la de minería (>15 m y 2 m, respectivamente), así como los registros de Escherichia coli (540 Vs. 337 UFC/100 mL), oxígeno disuelto (~1,8 veces mayor; 9,3 Vs. 5,2 mg O2/L) y sólidos suspendidos totales (63 Vs. 56 mg/L). En la estación de minería se registraron valores más altos para los sólidos disueltos totales (395 Vs. 111 mg/L), sulfatos (56 Vs. 21 mg/L), hierro (1,3 Vs. 0,4 mg/L), cloruros (2,9 Vs. 2,5 mg/L), fosfatos (1,2 Vs. 0,7 mg/L), grasas y aceites (0,9 Vs. 0,4 mg/L), conductividad (132 Vs. 108 |iS/m), y coliformes totales (3.156 Vs. 2.983 UFC/100 mL). Las demás variables no mostraron diferencias relevantes o registraron valores por debajo del límite de detección (Tabla 1S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2768).
Colonización de macroinvertebrados y pérdida de masa seca
En todos los tratamientos se recolectaron 18.070 individuos distribuidos en ocho órdenes, 24 familias y 34 géneros. En la estación de referencia se recolectaron 8.342 organismos: 2.197 asociados con hojas de Eucalyptus sp., 3.105 con T. lepidota y 3.040 con el sustrato mixto. En la estación de minería se recolectaron 9.728 organismos, 2.737 en Eucalyptus sp., 3.513 en T. lepidota y 3.478 en el tratamiento mixto. En ambas estaciones la familia más representativa fue Chironomidae (subfamilia Orthocladiinae), con 7.634 (78,5 %) en la estación de minería y 5.499 (66 %) en la de referencia (Tabla 2S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2769).
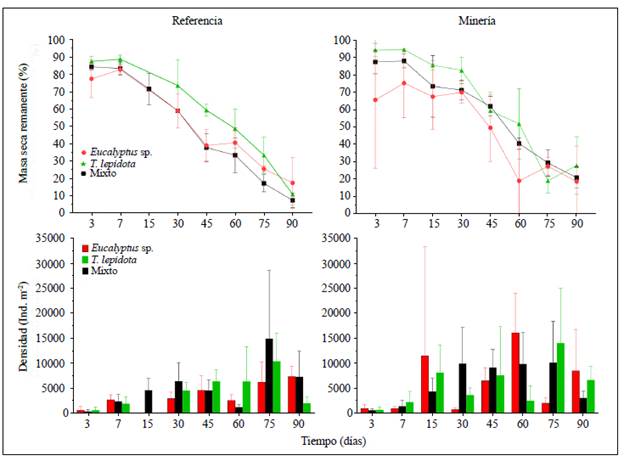
El porcentaje de masa seca remanente presentó diferencias significativas (p<0,001) entre las estaciones, los tratamientos y los eventos de muestreo. En el día 90 la estación de referencia presentó el mayor porcentaje de remanencia en el tratamiento de Eucalyptus sp., con 17,4 %, y menor porcentaje en el tratamiento mixto, con 7,3 %. En la estación de minería el mayor porcentaje de remanencia se presentó en el tratamiento con T. lepidota, con 27,8 %, y el menor, con Eucalyptus sp., 18,6 % (Figura 1). Las tasas de descomposición mostraron el siguiente patrón (Tabla 1S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2768): Eucalyptus sp. (estación de minería) > T. lepidota (estación de referencia) > mixta (estación de referencia). Ambas estaciones presentaron diferencias significativas en las tasas de descomposición entre los eventos de muestreo (referencia, p=0,025; minería, p<0,001), pero no en la riqueza de géneros (referencia, p=0,825; minería, p=0,108) (Tabla 3S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2770).
En la estación de referencia los valores más altos de densidad para los tres tratamientos se registraron en el día 75: enZ lepidota se recolectaron 27.689 ind./m2; en Eucalyptus sp., 15.511 ind./m2 y en el tratamiento mixto, 16.089 ind./m2. En la estación de minería las mayores densidades se registraron en el día 60 en el tratamiento mixto, con 21.067 ind./m2, en el día 75 en Eucalyptus sp., con 13.644 ind./m2, y en el día 45 en T. lepidota, con 16.800 ind./m2 (Figura 1). La densidad fue significativamente diferente entre las estaciones de muestreo (p<0,01) (Tabla 3S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2770).
Diversidad
La mayor riqueza usando la diversidad de orden q=0 (0D) se registró en la estación de referencia, con 42 taxones: en el tratamiento mixto se registraron 28 morfotipos, seguido por T. lepidota, con 25 y, por último, Eucalyptus sp., con 22. En la estación de minería se registraron 38 taxones: en T. lepidota, 22, enEucalyptus sp., 21 y en el tratamiento mixto, 20 morfotipos. La riqueza presentó diferencias significativas entre las estaciones (p<0,001) (Figura 2).
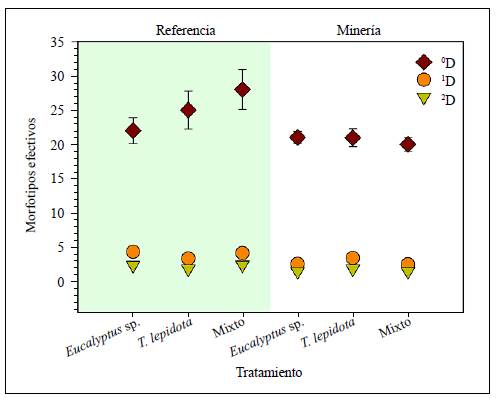
Figura 2 Diversidad de macroinvertebrados acuáticos con base en los números de Hill (promedio ± DE) asociados con la descomposición de hojas bajo tres tratamientos en dos estaciones de muestreo en la quebrada La Elvira (Manizales, Colombia)
La mayor diversidad q=1 (1D) se registró en la estación de referencia en el tratamiento de Eucalyptus sp., con 4,3 morfotipos efectivos, en tanto que la menor se registró en la estación de minería en el tratamiento mixto con 2,5 morfotipos efectivos (Figura 2). Estos resultados evidencian que el tratamiento con la especie exótica en la estación de referencia fue 1,8 (55 %) veces más diverso que el tratamiento mixto en la estación de minería. Con la medida de diversidad q=2 (2D), que es sensible a las especies más abundantes, los resultados fueron similares, es decir, los valores más altos se presentaron en la estación de referencia en Eucalyptus sp. y en el tratamiento mixto y, el valor más bajo se encontró en la estación de minería en el tratamiento mixto, con 1,4 morfotipos efectivos (Figura 2).
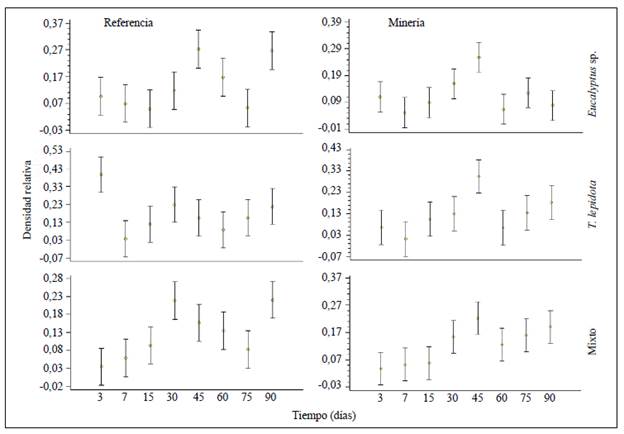
En la estación de referencia, en Eucalyptus sp., la mayor densidad relativa se presentó a los 45 días (0,27 ind./m2) y a los 90 (0,26 ind./m2); en el tratamiento mixto, en los días 30 (0,21 ind./m2) y 90 (0,22 ind./m2), y en T. lepidota, en los días 3 (0,39 ind./m2) y 30 (0,22 ind./m2) (Figura 3). En la estación de minería los datos mostraron un patrón similar: el tratamiento con Eucalyptus sp. presentó la mayor densidad relativa a los 45 días (0,23 ind./m2) y a los 30 días (0,15 ind./m2); el de T. lepidota y el mixto en los días 45 y 90 (0,30, 0,18; 0,22, 0,19 ind./m2, respectivamente) (Figura 3). El proceso de degradación de las hojas permitió evidenciar cinco categorías de colonización (temprana, tardía, inconsistente, intermedia y permanente), con algunos géneros representativos para cada categoría (Figura 1S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2771).
Discusión
Las altas densidades registradas para la subfamilia Orthocladiinae tanto en la estación de referencia como en la de minería, es un resultado relativamente común en las zonas templadas y tropicales donde se ha evaluado la dinámica de los macroinvertebrados en sistemas acuáticos bajo influencia minera (Cranston, et al., 1997; Czechowski, et al., 2020; De Bisthoven, et al., 2005). Además, estos organismos cumplen una función ecológica como recolectores (Henriques-Oliveira, et al., 2003; Galizzi, et al., 2012), pues se alimentan de la materia orgánica en partículas finas (MOPF) disponible en este tipo de experimentos desde el inicio de la descomposición de las hojas, y es empleada como potencial fuente de energía o como hábitat (Nessimian & Henriques-de-Oliveira, 2005; Callisto, et al., 2007; Chará-Serna, et al., 2010). La mayor abundancia de esta subfamilia también se ha reportado en otros estudios con paquetes de hojarasca en zonas templadas (Casas, et al., 2000; Abelho, 2008) y en el Neotrópico (Mathuriau & Chauvet, 2002; Carvalho & Uieda, 2004; Nessimian & Henriques-Oliveira, 2005; Callisto, et al., 2007; Carvalho, et al., 2008; Meza-Salazar & Rubio, 2010). Estos organismos pueden tolerar las condiciones de elevada acidez (pH=3,3) resultantes de procesos de minería en desarrollo o asociadas con procesos de lixiviación de minas abandonadas (Czechowski, et al., 2020; De Bisthoven, et al., 2005). En la estación de minería se presentaron valores comparativamente más altos de sólidos totales disueltos (~4 veces más altos), sulfato, hierro, cloruros y fosfatos (Tabla 1S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2768), asociados con la mayor presencia de estos quironómidos.
Los valores más altos de densidad, riqueza y diversidad de macroinvertebrados en la estación de referencia se asociaron con registros más bajos de masa seca remanente, principalmente en el tratamiento mixto, lo que corrobora nuestra predicción de que el sustrato más heterogéneo puede facilitar la colonización de una mayor riqueza o densidad de macroinvertebrados. Como es común, al final del experimento el material vegetal se encuentra en un estado avanzado de descomposición, lo que favorece el aumento de la MOPF disponible para los macroinvertebrados (Capello, et al., 2004). Además, la mayor presencia de vegetación ribereña en esta estación (Tabla 1S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2768) contribuyó a la entrada de material alóctono adicional disponible para los macroinvertebrados como alimento y refugio (Fisher & Likens, 1973; Abelho, 2001; Carvalho & Uieda, 2004; Ribeiro & Uieda, 2005). Por el contrario, en la estación de minería los valores de masa seca remanente al final del experimento fueron relativamente altos y se asociaron con menores valores de riqueza, densidad y diversidad de macroinvertebrados, así como con registros mayores de conductividad, sulfatos, grasas y aceites, y sólidos disueltos totales (Tabla 1S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2768). Además, la escasa vegetación ribereña no contribuye con el aporte de hojarasca para facilitar la presencia, la densidad y la biomasa de la fauna bentónica como se ha observado en otras microcuencas altoandinas (Guevara, et al., 2018). Estos resultados coinciden con los de otros estudios en los que se ha encontrado que, ante la ausencia de macroinvertebrados, los procesos de fragmentación y disolución de los compuestos de las hojas son más lentos (Quinn, et al., 2000; Graça, et al., 2002; Eyes-Escalante, et al., 2012).
La mayor masa seca remanente de Eucalyptus sp. en la estación de referencia se asoció con las características fisicoquímicas y la interacción de la comunidad de macroinvertebrados durante la colonización de las hojas y el reemplazo de los taxones. A pesar de no contar con información sobre la calidad química de las hojas, varios autores han resaltado que estas diferencias se relacionan con sus componentes secundarios, entre los que se encuentran fenoles y terpenos con actividad antimicótica, lo que interfiere con la acción fúngica y con la alimentación de los macroinvertebrados, los cuales no toleran estos compuestos, condicionando así su fragmentación (Graça, 2001; Graça, et al., 2002; Medina-Villar, et al., 2015). Asimismo, estos resultados coinciden con lo reportado por otros autores sobre la menor fragmentación de las hojas de especies vegetales exóticas en comparación con las hojas de especies nativas (Eyes-Escalante, et al., 2012). Si bien en condiciones naturales este suele ser el patrón en la descomposición, en el caso de la estación de minería la mayor masa remanente fue para T. lepidota. Durante el experimento se observó que este mayor porcentaje estaba representado por el material disponible de la venación de la hoja, aunque los demás componentes ya se habían degradado. Es probable que el cambio en la masa remanente de esta hoja entre las dos estaciones se relacione con alguna característica intrínseca de la planta, que podría reaccionar con algún factor químico presente en mayor cantidad en dicha estación (Tabla 1S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2768).
En la estación de referencia la sucesión degradativa evidenció que los tratamientos de Eucalyptus sp. y el mixto tuvieron "máximos de colonización" similares, lo que podría deberse a que en los periodos intermedio y final ocurre un aumento en la heterogeneidad del recurso alimentario, permitiendo que una mayor cantidad de organismos puedan aprovecharlo (Capello, et al., 2004; da Silva, et al., 2011). En el tratamiento con T. lepidota el patrón fue inverso, es decir, su periodo de colonización máximo se presentó al inicio y en el intermedio, lo que indica la estrecha relación entre los macroinvertebrados y esta planta nativa. En la estación de minería el patrón fue relativamente homogéneo: en Eucalyptus sp. el máximo periodo fue el intermedio, y en T. lepidota y el tratamiento mixto fueron el intermedio y el final.
Las categorías de colonizadores tempranos e intermedios correspondieron a Chirono-minae y Baetodes, respectivamente, ambos organismos colectores, lo que indica que se ali-mentan de MOPF, la cual se encuentra desde el inicio del proceso (Meier, et al., 1979; Nessimian & Henriques-de-Oliveira, 2005; Chará-Serna, et al., 2012 y Oliveira, et al., 2014), también reportaron la presencia de la subfamilia Chironominae desde los primeros días de muestreo, lo que quizá se deba a que los taxones menos especializados llegan primero al área, modificándola y permitiendo así que otros más especializados lleguen después (Carvalho & Uieda, 2004). Por otra parte, la presencia de Baetodes en el periodo intermedio del estudio se debió probablemente a que este organismo se alimenta de materia orgánica ya condicionada, es decir, que ha pasado por un proceso de descomposición previo (Quinn, et al., 2000; Carvalho & Uieda, 2004), como fue reportado por Capello, et al. (2004) en un estudio en el río Paraná en Argentina. Entre los colonizadores permanentes se encontró el género Atopsyche, descrito como depredador por Chará-Serna, et al. (2010), el cual se presentó durante todos los eventos de muestreo. El recambio de organismos y potenciales presas durante la fase de colonización y descomposición a lo largo del experimento contribuyó a una mayor oferta alimentaria.
El organismo representante de la categoría de colonizadores que apareció como inconsistente fue Simulium, catalogado por Cummins, et al. (2005) como colector-filtrador. Su fuente de alimento son las partículas suspendidas en la columna de agua y, generalmente, muestra un comportamiento inestable cuando se ve afectado por cambios en los factores abióticos o por las fluctuaciones en la oferta del recurso (Motta, et al., 2016). El representante de los colonizadores tardíos fue Maruina, clasificado como raspador (Cummins, et al., 2005), cuya alimentación consiste en perifiton y algas adheridas a cualquier superficie. Sin embargo, se ha resaltado que los organismos raspadores son escasos en las quebradas de cabecera (Chará-Serna, et al., 2010), por lo tanto, la mayor densidad hacia el final de los muestreos se relacionaría con el hecho de que en la superficie de las bolsas de malla plástica utilizadas para el estudio pueden crecer algas o perifiton que sirve de alimento.
Conclusiones
El proceso de colonización de macroinvertebrados acuáticos sobre hojas individualizadas y mixtas de Tibouchina lepidota y Eucalyptus sp. evidenció que un sustrato más heterogéneo en zonas sin impacto antropogénico evidente (estación de referencia), puede facilitar el reemplazo progresivo de macroinvertebrados asociados con una mayor riqueza o densidad de determinados grupos (coleópteros [Heterelmis, Anchytarsus], dípteros [Probezzia], anélidos [Naididae] y tricópteros [Nectopsyche]), comparado con el de zonas impactadas por actividades mineras. Sin embargo, aunque en la estación de minería se obtuvo una menor riqueza total, la densidad global fue comparativamente mayor (Tabla 2S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2769), la cual se reflejó en la presencia de algunos efemerópteros (Baetodes), dípteros (Orthocladiinae, Chironominae, Maruina) y tricópteros (Hydroptila). Asimismo, estos resultados pueden estar asociados con la mayor concentración de determinadas variables fisicoquímicas en áreas con actividades mineras y con una mayor cobertura de la vegetación ribereña en áreas de referencia (Tabla 1S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2768).
La hipótesis propuesta fue coherente con los resultados encontrados. Sin embargo, el análisis debe hacerse con precaución, tanto el de las variables fisicoquímicas como el de los descriptores biológicos (riqueza, diversidad, densidad, pérdida de masa seca y tasas de descomposición). En general, se encontró una mayor densidad de macroinvertebrados colonizadores asociada con la especie nativa (T. lepidota) que con la exótica (Eucalyptus sp.), independientemente del efecto (referencia o minería) en la quebrada de estudio (Tabla 2S,https://www.raccefyn.co/index.php/raccefyn/article/view/1059/2769).
El presente estudio permitió dilucidar cómo se dan los reemplazos progresivos de unos organismos por otros en la sucesión degradativa de hojas de T. lepidota y Eucalyptus sp. en la quebrada La Elvira (Manizales, Colombia). Además, contribuye al conocimiento de la dinámica de la colonización de macroinvertebrados acuáticos y su papel funcional en la descomposición de MOPG en sistemas lóticos altoandinos. Esta información es fundamental para predecir futuros efectos de otras presiones antropogénicas, como la agricultura y la ganadería, sobre la vegetación ribereña y su relevancia para el flujo de nutrientes y descriptores ecológicos de los macroinvertebrados acuáticos en sistemas lóticos.

