











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introduction
The establishment of forest plantations with fast-growth species, such as the genus Eucalyptus (Myrtaceae), represents a high profitability option for commercial reforestation (Obregon & Restrepo, 2006). Eucalypts have a great capacity of adaptation to a wide range of site conditions, different management systems, and different end uses (Moura, 2001); moreover, they cover more than 20 million hectares around the world (Ferreira, et al., 2019) mainly in tropical and sub-tropical countries. In Colombia, the genus Eucalyptus is one of the most used by forestry companies and it is estimated that approximately 57 thousand hectares are planted with this genus in the country (PROFOR, 2017).
One of the most important decisions to consider when establishing new forest plantations is the selection of the species to be planted, as well as the choice of an adequate planting spacing to ensure the maximization of wood production. The selection of suitable species is a fundamental stage in forestry (Rollan, et al., 2018). Choosing the right species for each environment favors the formation of high-yield plantations. In tropical regions, the most important selection factors for assigning a species are the level of productivity, the resistance to pests and diseases, the tolerance to drought, and the suitability of wood properties (Gonçalves, et al., 2013).
Several studies evaluating Eucalyptus growth in different environmental conditions have shown that the yield of the same species varies according to the place where it is grown (Coutinho, et al., 2004; Delgado-Matas & Pukkala, 2011; du Toit, et al., 2017; Ferreira, et al., 2017). Since each species develops in a specific environment, the species selection process needs to be site-specific, an approach that results in satisfactory yields according to the potential environmental site conditions (Del Quiqui, et al., 2001; Moura, 2001). In this regard, species trials are a useful tool to evaluate species and select the one with the highest productivity and the best adaptation to the planting site.
The climate in tropical dry forests is characterized by evapotranspiration that greatly exceeds precipitation during most of the year with subsequent water deficit (Pizano & García, 2014). These conditions make plant productivity dependent on the amount of water available and its efficient use by the organism (Yepes & Buckeridge, 2011). According to Gonçalves, et al. (2013; 2017), in the case of the Eucalyptus genus, the most drought-tolerant pure and hybrids species are Eucalyptus urophylla S.T. Blake, Eucalyptus camaldulensis Dehnh, Eucalyptus tereticornis Sm., E. granáis x E. urophylla, E. grandis x E. camaldulensis, and E. urophylla x E. camaldulensis. In Colombia, the frequent use of at least six eucalypt species by forest producers has been documented (Borralho & Nieto, 2012). However, there is still a lot of uncertainty regarding the species to be planted and their potential productivity, especially in marginal sites with water restrictions.
On the other hand, the selection of an adequate planting spacing is important because it affects site occupation, average log sizes, and wood quality, which are key components in the forest plantation economy (Akhtar, et al., 2008). Determining the planting density is a complex and important decision for producers since it affects the whole cycle given its significant effect on individual trees and stand growth (Guerra, et al., 2014). In eucalypt forest plantations the initial space between trees has been based on spacing trials, which usually range between 1000 and 2500 stems-ha-1 (Bernardo, et al., 1998; Meskimen & Franklin, 1978).
High tree densities provide higher growth per hectare implying more intense competition for resources and faster site occupation (Chen, et al., 2011; Forrester, et al., 2013). Additionally, in high-density plantations the maximum current (CAI) and mean annual increments (MAI) may also occur earlier during stand development. For the production of Eucalyptus pulp and paper, 3x2 m (1666 stems ha-1) and 3*2.5 m (1333 stems ha-1) spacings have frequently been implemented (Caterina, 2017). Previous studies have shown that higher densities also have effects on wood properties. For instance, Rocha, et al. (2016) found that the planting of E. grandis x E. camaldulensis clones in a 4.5 to 9.0 m2 (2222 to 1111 stems ha-1) spacing increased wood density and resulted in the production of raw materials more suitable for energy purposes.
Conversely, lower densities generated trees with larger sizes but took longer to occupy the site completely while the growth rate of the stand may be slower and the rotation age occurred later (Forrester, et al., 2013; Stape & Binkley, 2010). For the production of high-quality saw timber, a wider initial spacing (500 to 800 stemsha-1) is recommended in sites of good quality as it ensures a high potential for increasing diameter during the first three years (Nutto, et al., 2006).
Planting densities in Colombia have often been prescribed without considering the capacity of the site, the species, and the interaction site-species, which has led to a waste of the productive capacity of the sites (Barrios, et al., 2011). Therefore, it is important to evaluate the species, as well as their interaction with planting density, to maximize site productivity. In this context, the objective of this study was to evaluate the early silvicultural performance of nine species of Eucalyptus in three planting spaces under the environmental conditions of tropical dry forests in a site located in the inter-Andean valleys of Colombia.
Materials and methods
The species trial was conducted in an experimental area of the Universidad del Tolima located in the municipality of Armero, Tolima, Colombia (5°00'58.1"N; 74°54'27.2"W) (Figure 1) in a tropical dry forest (Holdridge, 1947) of the inter-Andean valleys at 227 meters above sea level, a mean annual temperature of 28 °C and average relative humidity of 71 %, the total annual precipitation is 1700 mm with a bimodal distribution: a rainy period from April to June and another one from October to December. As regards the climatic water balance (precipitation - evapotranspiration), there are eight months with climatic water deficit (Fernández, et al., 2014). The landscape geomorphology corresponds to that of piedmont with a terrace-fan relief, low slopes of 0-3 %, and soils with sandy clay loam texture (IGAC, 2004).
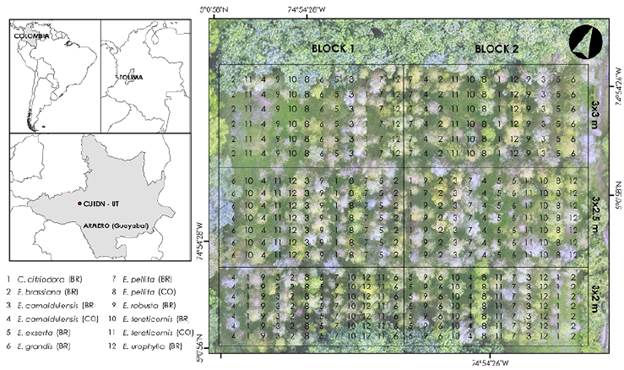
Figure 1 Study area location and experimental design of the eucalypt species trial in Armero (Guayabal), Tolima, Colombia. BR: Provenance from Brazil; CO: Provenance from Colombia
We used seeds of nine eucalypt species for seedlings production. All the nine eucalypt species were provenances from Brazil (BR). We also evaluated three provenances from Colombia (CO) (Table 1). The seedlings were kept in the nursery for three months until they reached an average height of 30 cm, approximately. Site preparation included sub-soiling at a 60 cm depth and the use of a hydro-retaining polymer for each individual at the moment of planting. We applied two fertilizations during the first year and carried out at least four yearly mechanical weed controls.
Table 1 Species and provenances of eucalypt evaluated
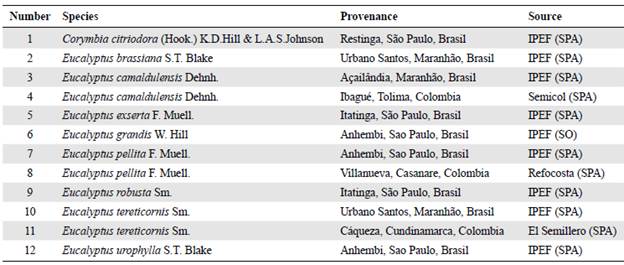
* IPEF: Instituto de Pesquisas e Estudos Florestais; SPA: Seed production area; SO: Seedling orchard
We used a split-plot design where the whole plots (main plots) were the planting spacing factor with three levels: 3*2 m (1666 stemsha-1), 3x2.5 m (1333 stemsha-1), and 3x3 m (1111 stemsha-1) while subplots were the species factor with 12 levels (Table 1), and two replicates (blocks). The stand densities evaluated are commonly associated with management prescriptions for pulp and saw timber production (Barrios, et al., 2011). Each subplot was comprised of a row of around 6 to 7 trees of each of the species under evaluation as recommended for species trials (Lott & Wardill, 2009) (Figure 1). The split-plot design is commonly used in forestry research, as it is suitable for two-factor experiments in which the levels of one of the factors require a larger plot size while the other requires smaller size plots (Jayaraman, 1999; Lott & Wardill, 2009).
We began taking measurements after 12 months of planting and repeated them at 18 and 24 months. The variables measured included diameter at breast height (dbh), total height, crown diameter, basal area, volume, and survival. We measured the dbh 1.3 m above ground level with a diameter tape, the total height with a laser hypsometer (Nikon Forestry 550), and the crown diameter with a laser range finder (Meco® Laser Distance Meter, DLDM60). We determined the volume (v) per individual tree as g x h x ff where (g) is the basal area, h the total height (m), and ff the form factor assumed as 0.5. This form factor was selected based on previous studies on eucalypts. Nieto & Gasca (2010) used a form factor of 0.5 to evaluate Eucalyptus pellita superior trees in Colombia. Delgado & Pukkala (2011) reported a form factor of 0.51 for different species of tropical Eucalyptus growing in Africa. Likewise, Hernández-Ramos, et al. (2017) reported a form factor of 0.45 for Eucalyptus urophylla in Tabasco, México, and Acosta (2018) one that varied from 0.46 to 0.54 for Eucalyptus grandis grown in Chimborazo, Ecuador.
The basal area and the volume per hectare were obtained from each treatment's plot level and then expanded to a hectare. The mean annual volume increment (MAI) corresponded to the quotient between the volume per hectare and the actual age of the trial. The survival (in percentage) was determined as the quotient between the number of live trees and the overall planted trees. Additionally, to evaluate the adaptation of eucalypt species to the environmental conditions of the study area, we evaluated the global phytosanitary status of each tree considering four levels of affectation modified from Carrasco, et al. (2019) (1= healthy; 2= mildly affected, i.e., < 25 % affected; 3= moderately affected, i.e., between 25 and 50 % affected, and 4= severely affected, i.e., > 50 % affected).
At 24 months of age, we submitted the experimental data to an analysis of variance using the Statistical Analysis System (SAS) software (SAS Institute Inc., 2009). The Kolmogorov-Smirnov normality test was used to assess the normality of the data and homogeneity of variances was verified by observing the fitted model residuals distribution. Prior to the statistical analysis, the survival data were transformed applying an arcsine square root transformation to meet normality. We used the following model for the split-plot design (Montgomery, 2004; Littell, et al., 2006):
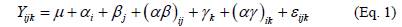
where Y jjk is the response variable, μ the overall mean, α. the effect of the ith planting spacing, β j . the block random effects, (αβ) ij . the random error of whole plots, y k the effect of kth species, (αy).k the interaction between planting spacing and species, and ε jjk the random error of subplots. When the analysis of variance found significant differences for some factors, we used a Tukey multiple comparison test to find the treatments that were different. All statistical analyses were performed considering a level of significance of α=0.05.
Results
The analysis of variance found significant differences between species for all the variables evaluated (p<0.05). Conversely, planting spacing only had a significant effect on the basal area (p<0.01) and the volume (p<0.05) (Table 2). The analysis of variance also provided evidence to discard the interaction between the planting spacing and species factors for all the variables evaluated (p>0.05) (Table 2).
Table 2 Analysis of variance of the main interactive effects of planting spacing and species
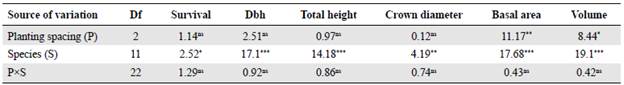
For each variable, the F-value and the significance level of the analysis of variance is shown: *, ** and *** indicate significant effects at p<0.05, p<0.01, and p<0.0001, respectively; ns: Non-significant
In general, all the species showed a good adaptation to the environmental conditions of the study area with survival rates between 67.9 % and 97.6 % (overall mean of 82.2 %). Among the species evaluated, we highlight the Brazilian provenances of E. camaldulensis and E.pellita with survival rates of 97.6 % and 92.5 %, respectively (Table 3). Conversely, E. robusta (BR) and E. tereticornis (BR) showed the lowest survival rates with 67.9 % (Table 3). When comparing the species from different origins, we found that the survival of the Brazilian provenances of E. camaldulensis and E.pellita was 19.8 and 10.3 % higher than the Colombian provenances, respectively. Contrarily, the survival rate of the Colombian provenance of E. tereticornis was 8.7 %, higher than the Brazilian provenance.
Table 3 Means and Tukey's grouping of survival and growth variables for eucalypt species at 24 months of planting. Means with different letters are statistically different (p<0.05).
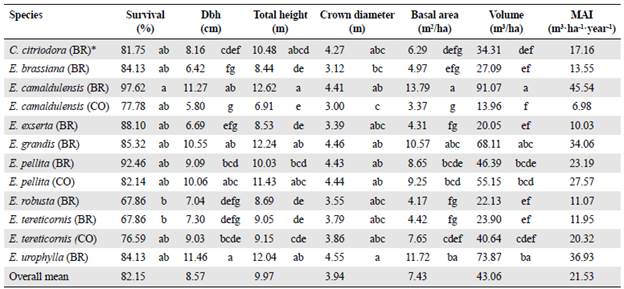
* BR: provenance from Brazil; CO: provenance from Colombia
The species with the highest volume after 24 months of planting were the Brazilian provenances of E. camaldulensis (91.07 m3 ha-1), E. urophylla (73.87 m3 ha-1), E. grandis (68.11 m3ha-1), and the Colombian (55.15 m3ha-1) and Brazilian provenances of E. pellita (46.39 m3 ha-1). These species also showed the best growth performance. Considering volume production, the Colombian provenances of E. pellita and E. tereticornis were slightly higher than those of Brazil. However, the Brazilian provenance of E. camaldulensis was far superior to that of Colombia (Table 3).
As for stand basal area and volume, the 3*2 m and 3x2.5 m treatments were not significantly different (p>0.05); however, they were significantly different from the 3x3 m treatment (p <0.05) (Table 2). The analysis showed that the planting spacings of 3x2 m and of 3x2.5 m maximized the production of the basal area and volume reaching an average of 8.52 m2ha-1 and 49.19 rrfha1, and 7.86 m2ha-1 and 46.13 m3 ha-1, respectively. Conversely, the basal area and volume production notoriously decreased (5.92 m2ha-1, 33.86 m3 ha-1) with the higher spacing (3x3 m).
Figure 2a shows the development of the mean annual volume increment (MAI) for each of the species under evaluation from the 12th to the 24th month: 24 months after planting, the MAI showed an average of 21.53 m3 ha-1 year-1 ranging from 6.98 to 45.54 m3 ha-1 year-1. Interestingly, the lowest MAI was that of the Colombian provenance of E. camaldulensis and the highest that of the Brazilian provenance of E. camaldulensis. As shown in figure 2a, the Brazilian provenances of E. camaldulensis, E. urophylla, E. grandis, and the Colombian and Brazilian provenances of E. pellita were the fastest-growing species among those under evaluation. Narrow planting spaces promoted a higher growth among the species evaluated reaching an average MAI of 24.59 and 23.06 m3ha-1 year-1 for 3x2 m and 3x2.5 m planting spacings, which were 45 and 36 % higher than the MAI reached by the 3x3 m planting spacing (16.93 m3 ha-1 year-1), respectively (Figure 2b).
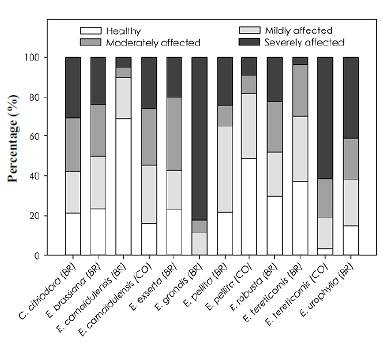
Although all the species showed some level of phytosanitary affectation, E. camaldulensis (BR) and E. pellita (CO) had the best adaptation to the study area with 69.2 % and 48.5 % of healthy trees while we found a higher percentage of trees severely affected for E. grandis (BR) (82.3 %) (Figure 3). The main damages were defoliation of the tree canopy caused by leafcutter ants and the occurrence of termites in the bark and semi-occluded dead branches of the trees. Additionally, bark lesions (e.g., bark splitting) were observed at the base of the stems in some trees, especially E. grandis specimens.
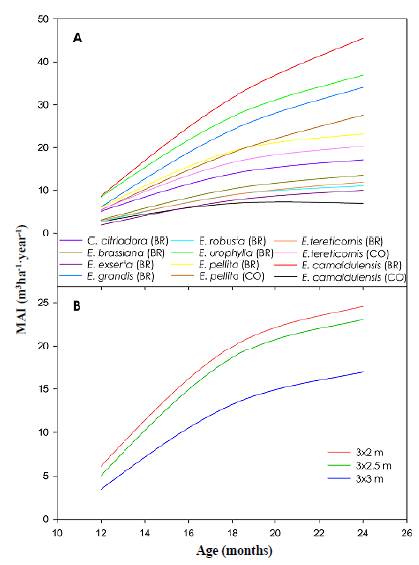
Figure 2 Development of the mean annual volume increment for each eucalypt species (A) and planting spacing (B). BR: Provenance from Brazil; CO: Provenance from Colombia.
Discussion
We found significant differences among the eucalypt species under study in a tropical dry forest site in terms of survival, dbh, total height, crown diameter, basal area, and volume. In general, high survival rates ranging between 67.86 and 97.62 % were obtained for all the species evaluated. Survival is a relevant variable for the evaluation of species adaptation; however, the selection and recommendation of any genotype must also consider growth and productivity, resistance to drought, pests, and diseases (Fernandes, et al., 2018).
Eucalyptus camaldulensis (Brazil provenance) was superior in terms of survival (18.8 % over the overall mean) and growth (111.5 % over the overall mean) compared to the other species evaluated. Bush, et al. (2013) and Costa, et al. (2012) also reported high survival rates among the provenances and progenies of this species. These authors have highlighted the great environmental plasticity of E. camaldulensis to adapt to nutrient-poor and low rainfall sites and tolerate moderately saline and humid soil conditions. Conversely, the Colombian provenance of E. camaldulensis was the species with the lowest growth and yield in the study. This contradictory result could be explained by the differences in the quality and the genetic improvement of the seeds between the Brazilian and Colombian provenances. However, previous studies have also reported wide variations among E. camaldulensis provenances. Del Quiqui, et al. (2001) registered survival rates between 75 and 96 % and an MAI between 28.81 and 40.21 m3-ha-1-year-1 among E. camaldulensis provenances in the state of Paraná, Brazil.
Other species showing good adaptation to the study area were E. urophylla, E. grandis, and E. pellita (Colombia and Brazil provenances), with survival rates of 84.1 %, 85.3 %, 82.1 %, and 92.5 % and an MAI of 36.93, 34.06, 27.57, and 23.19 m3ha-1 year-1, respectively. In a previous study with provenances of E. urophylla and E. grandis in Colombia, Wright & Osorio (1996) reported stem volumes 50 % higher for E. grandis than for E. urophylla. However, in the present study, the individual stem volume of E. urophylla was 16 % greater than that of E. grandis. For E. grandis,Albaugh, et al. (2015) have reported an MAI between 50 and 108 m3ha1year-1 in fertilization trials in six sites distributed in the Colombian Andes.
Our results agree with those reported by Silva, et al. (2019a; 2019b) when evaluating survival and growth in 3-year progeny trials of E. urophylla and E. grandis in different climatic zones of Brazil. These authors reported a survival rate of 51 % to 92 % and an MAI of 19 to 46 m3 ha-1 year-1 for E. urophylla and of 28 to 89 % and 20.5 and 48.4 m3-ha-1 -year-1 for E. grandis.
The provenances of E. pellita had a favorable adaptation to the test site with similar growth and slightly different survival. In our study, both provenances of E. pellita showed a higher MAI compared to that reported by Ferreira, et al. (2017) of 18.2 m3 -ha-1-year-1 for this species growing in the state of Rio de Janeiro, Brazil. The growth reached by the Colombian E. pellita provenance is consistent with that reported by Nieto, et al. (2016) in the Colombian Orinoco after 4 years of growth. However, Nieto & Gasca (2010) reported an average MAI of 19.5 m3-ha-1 -year-1 for E.pellita growing in Casanare, Colombia, which is lower than that reported in the present study. Nieto, et al. (2016) highlight the potential of E. pellita to adapt to high humidity site conditions. However, in our study, the species showed good adaptation to low rainfall compared to its natural distribution, which may be explained by the fact that although water deficits are common in our study area, they are not as severe as in other areas of the tropical dry forest of Colombia.
Eucalyptus tereticornis, C. citriodora, E. brassiana, E. robusta, and E. exserta yields were below the average in most of the variables evaluated. Although E. tereticornis has been planted on the Colombian Atlantic Coast due to its resistance to drought with an MAI of 32.8 m3-ha-1-year-1 (Hernández, 1993), in our study the species did not show a good adaptation.
Given the limited experience with C. citriodora, E. brassiana, E. robusta, and E. exserta commercial plantations, there is little information on their silvicultural requirements and field behavior; however, previous studies agree with ours regarding the low growth rates of these species. Poersch, et al. (2017) reported a survival rate of 52.7 %, a height of 6.0 m, and collar diameter of 8.5 cm for C. citriodora 19 months after planting. In Madagascar, E. robusta grows well in low and medium altitude areas and acid and infertile soils, with an MAI between 11.3 and 15.8 m3-ha-1-year-1 two years after planting (Verhaegen, et al., 2014). In a species test, E. robusta reached a volume of 247.8 rrf-ha1 eight years after planting, one of the lowest in the study due to its high mortality in the field (Bôas, et al., 2009). Drumond, et al. (1998) tested three provenances of E. brassiana at eight years with a survival of 98 % and an MAI of 13.5 m3-ha-1-year-1 as their best result. In the south of China, E. exserta plantations were established to stop soil erosion given their ability to tolerate sandy soils with low nutrient content, however, their growth rates were very low, between 4 and 8 m3-ha-1-year-1 (Turnbull, 2007).
According to Gonçalves, et al. (2017), the MAI in eucalypt plantations in regions with nutritional deficiencies and water stress ranges between 20 and 45 m3-ha-1-year-1 depending on the level of environmental stress. Taking as reference the lower limit reported by these authors, half of the evaluated species achieved favorable growth rates while the remaining showed lower growth rates, possibly due to inadequate site conditions for appropriate performance. The Brazilian provenances of E. camaldulensis, E. urophylla, E. grandis, E. pellita, and the Colombian provenance of E. pellita showed high survival rates and the highest growth rates. Poersch, et al. (2017) had similar results with E. urophylla, E. camaldulensis, E. grandis, and E. saligna Sm. showing the best adaptation in a study in the state of Mato Grosso do Sul, with a mortality rate lower than 10 % during the early stages of development.
In our study, the interaction between planting spacing and species was not significant for all the variables under evaluation, which indicates that a species that performs adequately in a specific planting spacing keeps the same behavior in the other evaluated spacings.
Planting spacing had a significant effect only on stand basal area and volume. In our study, narrow spaces showed the greatest increases in basal area and volume, and a contrary trend was presented in wider spaces. The average MAI ranged from 16.9, 23.1 to 24.6 m3-ha-1-year-1 for 3x3, 3x2.5, and 3x2 m, respectively. Previous studies on Eucalyptus have shown that larger planting spacings promoted the development of individual trees while narrow spacings increased production at the stand level (Nagar, et al, 2015; Hakamada, et al., 2020). Nagar, et al. (2015) obtained the maximum total utilizable biomass (72.72 kg-tree-1) from the wider spacing of 3x3 m and the maximum total biomass (113.59 ton-ha-1) from the narrowest spacing of 3x1 m for E. camaldulensis in India. Similar findings were reported by Forrester, et al. (2013) in their study of the basal area growth of E. globulus Labill. planted in five sites with different qualities and six stand densities. Moreover, they reported no interaction between site quality and spacing; consequently, the effects of planting spacing obtained in a specific site quality may be generalized to other sites with different qualities.
Likewise, Filho, et al. (2018) found that eucalypt stands planted with low densities (667 stems-ha-1) showed a lower mortality rate compared to those with higher densities (1111 stems-ha-1), a result attributed to lower competition between trees in the stands with lower densities. In our study, there were no significant differences in survival between planting spacings, however, the highest survival rates were obtained with narrow planting spaces possibly because the specimens were young and the crown had not closed yet, after which the competition is expected to increase within the stand.
The non-significant effect of planting spacing on the total height is supported by the hypothesis that this variable is not influenced by the competition and the stand density (Vospernik & Sterba, 2015). Although there were no significant effects of planting spacing on dbh and crown diameter, other studies have reported a strong effect of this factor on these variables and a strong relationship between them (França, et al., 2017; Schuler, et al., 2013). Nutto, et al. (2006), Pinkard & Neilsen (2003), and Xue, et al. (2011) found for E. urophylla, E. grandis, and E. nitens (H. Deane & Maiden) Maiden stands that both the dbh and crown diameter decreased when the stand density increased. These authors agree that increasing stand density generates more intense competition for resources, which makes the above-ground space more limited affecting the photosynthetic specific area and, consequently, reducing the dbh growth.
In our study, we evaluated the global phytosanitary status of each tree to identify the damage present in the trees and the possible causal agents. All the species showed some type of phytosanitary affectation, which was the main cause of mortality and growth decrease in the trial. Our results showed that the Brazilian E. camaldulensis provenance and the Colombian E. pellita provenance registered the fewest pests and diseases.
The main damages identified in the trial were defoliation by leafcutter ants and termites in the bark and semi-occluded dead branches. The study made no attempt to taxonomically identify the causative agents, nevertheless, those we identified coincided with previous studies by Ospina, et al. (2006) and Nieto & Gasca (2010) who identified the genus Atta spp. (Hymenoptera order) and Heterotermes spp. (Isoptera order) as the main causal agents of defoliation and damages on dead branches and bark. Wilcken, et al. (2002) reported four groups of termites that caused some damage in eucalypt plantations in Brazil: a) seedling/ sapling termites, b) heartwood termites, c) bark termites, and d) wood termites.
The bark splitting we report here has been previously related to affectations by eucalypt canker (Chrysoporthe spp.) in some species like E. grandis (Ospina, et al., 2006; Juárez, et al., 2013), E. urophylla (Juárez, et al., 2013), and E. pellita (Chocontá, et al., 2017). Pest and disease outbreaks become more common with increasing water stress, especially in tropical regions, posing a serious threat to the productivity of eucalypt plantations (Gonçalves, et al., 2017).
The variations found in this study regarding the behavior of eucalypt species highlight the importance of studies on species selection and the need to deepen research in genetic and silvicultural strategies necessary for forest management in eucalypt plantations under water stress conditions.
Conclusions
The eucalypt species we evaluated presented an adequate adaptation to the study site with an average survival of 82 % 24 months after planting. The Brazilian provenances of E. camaldulensis, E. urophylla, E. grandis, and E. pellita, and the Colombian provenance of E. pellita were identified as promising for commercial reforestation programs in areas with water limitations in Colombian tropical dry forests. Our study showed that the development of each species is independent of planting spacings and that those of 3x2 m and 3x2.5 m maximized the basal area and the volume production of the species evaluated. Finally, the study showed that species with different origins vary in productivity at the same site while pests, diseases, and the quality of genetic material can modify growth and yield.

