











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Los estuarios son ecosistemas de transición que se caracterizan por presentar gradientes ambientales producto, ante todo, de la mezcla entre los sistemas marinos y de agua dulce (Ferreira et al., 2019). Estas zonas son muy productivas, exhiben variaciones naturales en los parámetros bióticos y abióticos, y presentan cambios estacionales (Mcelligott & Chick, 1996) que, en su conjunto, influyen en la disponibilidad de recursos, así como en las estrategias de alimentación de la biota (Blasina et al., 2016)Sciaenidae. En consecuencia, la dinámica de los parámetros oceanográficos dentro de un área relativamente pequeña y semicerrada resulta en una diversidad de hábitats, así como en la complejidad de los factores naturales y las interferencias antrópicas, entre estas, la contaminación (Ferreira et al., 2019).
En este sentido, los peces que pasan mayor tiempo en los estuarios son vulnerables al consumo incidental de todo tipo de contaminantes, especialmente de microplásticos (Ferreira et al., 2019). Los microplásticos son micropartículas sintéticas con un diámetro inferior a los 5 mm, que se generan de fuentes primarias y secundarias. Los microplásticos primarios se fabrican intencionalmente para diversas aplicaciones, entre las que se destacan los productos de cuidado personal y cosméticos (por ejemplo, limpiadores faciales, detergentes, pasta de dientes, polvos faciales, cremas desvanecedoras, espumas de afeitar, entre muchos otros); los secundarios, por su parte, se originan a partir de procesos físicos, químicos y biológicos que resultan en la fragmentación del plástico (Ashrafy et al., 2023). Los microplásticos pueden llegar a los estuarios a través de la disposición directa de residuos sólidos, el desgaste de las redes de pesca y transportados en los efluentes domésticos (Barletta et al., 2020).
Asimismo, los microplásticos pueden ser fácilmente ingeridos por la fauna acuática debido a su alta biodisponibilidad y su tamaño (Lusher, 2015). La ingestión de microplásticos puede provocar diversas anomalías morfológicas en los peces, estrés oxidativo y fisiológico, reducción de las actividades enzimáticas, falsa satisfacción del hambre, baja tasa de crecimiento, complicaciones reproductivas (Sutton et al., 2016), así como obstrucciones internas y daños en el tracto digestivo (Baalkhuyur et al., 2018). Además, se ha registrado que la presencia de microplásticos en el tracto gastrointestinal puede generar efectos negativos asociados con los aditivos incorporados durante su fabricación o los contaminantes ambientales absorbidos por los plásticos en el transcurso del tiempo (Rochman et al., 2013). Por otro lado, estas partículas pueden biomagnificarse a lo largo de las redes tróficas debido a su naturaleza no degradable (Reisser et al., 2014). Por lo tanto, los depredadores superiores pueden estar más expuestos tanto a la ingestión directa como a la transferencia trófica de una presa previamente contaminada con microplásticos (Ferreira et al., 2019).
Por ello, cobra importancia el estudio de la ecología trófica de los peces, ya que permite examinar la superposición de nichos ecológicos y la competencia entre depredadores, e investigar las relaciones intraespecíficas e interespecíficas en la red trófica, siendo esto fundamental en los modelos de pesquerías de múltiples especies (Sá et al., 2006). Para comprender la ecología trófica de los peces deben estudiarse sus hábitos alimentarios. Sin embargo, dadas las dificultades inherentes a la observación in situ de dichos hábitos, se ha utilizado el análisis del contenido estomacal como el método más común para estudiar su dieta (Sá et al., 2006). También, los hábitos alimentarios y la distribución espaciotemporal son muy variados, lo cual reduce la competencia por recursos (Piet et al., 1998). Muchos de los patrones de comportamiento y las adaptaciones morfológicas de las especies han evolucionado en respuesta a la necesidad de capturar alimentos, los requisitos de reproducción y la evasión de depredadores (Labropoulou & Eleftheriou, 1997). Así, dichas adaptaciones les permiten aprovechar determinados tipos de presa (Russo et al., 2008).
En el caso de los peces de la familia Sciaenidae (conocidos comúnmente como corvinas), se han registrado especies demersales que habitan fondos lodosos o arenosos (Rueda et al., 2006), en su mayoría de hábitos costeros y dependencia estuarina durante etapas ontogénicas debido a su gran adaptabilidad a las condiciones fisicoquímicas fluctuantes (temperatura, salinidad, turbidez y oxígeno disuelto) (Blasina et al., 2016). En el Pacífico colombiano se han registrado 49 especies (Muriel-Hoyos & Carmona-Guerra, 2020), las cuales contribuyen de manera importante a la riqueza y ensamblaje de peces demersales en él (Sánchez-Gil & Yáñez-Arancibia, 1998). Son principalmente depredadores y su dieta incluye peces pequeños, invertebrados bentónicos y cefalópodos (Castro, 2019). Constituyen un componente importante de las pesquerías artesanales y de arrastre industrial, siendo el grupo más diverso y uno de los de mayor abundancia dentro de la captura incidental del camarón de aguas someras (Castro, 2019; Martínez-Muñoz, 2012; Zárrate, 2008) Además, constituyen una importante fuente de proteína y son apetecidos comercialmente en el mercado local y nacional (Rueda et al., 2006).
Los peces de la familia Sciaenidae en la bahía de Tumaco han sido importantes para el comercio y la soberanía alimentaria de los habitantes de la zona; sin embargo, se registran pocos estudios sobre sus hábitos tróficos en esta bahía. Por ello, en este estudio nos propusimos determinar la ecología trófica y el consumo incidental de microplásticos en peces de la familia Sciaenidae, y su variación con respecto a las dinámicas ambientales en la bahía de Tumaco. Por un lado, se esperaba que las variaciones espaciotemporales en la bahía influyeran en la abundancia y la disponibilidad de diversas presas apetecibles para los peces, por lo que se presentarían diferentes nichos entre especies. Por otro lado, teniendo en cuenta la inadecuada gestión de residuos sólidos en la bahía de Tumaco y la gran cantidad de residuos plásticos que terminan en las zonas costeras (Villarreal & Bastidas, 2016), se esperaba encontrar en los peces de la familia Sciaenidae la ingesta incidental de microplásticos debido a dicha contaminación.
Materiales y métodos
Area de estudio
La bahía de Tumaco se ubica en el extremo sur de la costa pacífica colombiana, en el departamento de Nariño, entre la latitud 1° 45' y 2° 00' N y la longitud 78° 30' y 78° 45' W, con un área aproximada de 350 km2 y profundidades que varían entre 0 y 50 m (Figura 1) (Tejada, 2003); recibe el aporte de ocho ríos, entre los cuales se destacan el Rosario, el Mira y el Patía. Este hecho, y la gran cantidad de sedimentos que recibe, hacen de la bahía un sistema hidrográfico complejo (Tejada et al., 2003). La bahía presenta, además, las temperaturas más bajas del Pacífico colombiano debido a factores como la radiación solar, la distribución de vientos, las precipitaciones, la intensidad de los aportes fluviales en las desembocaduras de los ríos y las variaciones de la corriente de Humboldt y la contracorriente ecuatorial (IDEAM-METEO, 2008).También registra una alta humedad, con un promedio anual superior al 86 %, una elevada nubosidad y pocas horas de brillo solar durante la mayor parte del año (Tejada et al., 2003).
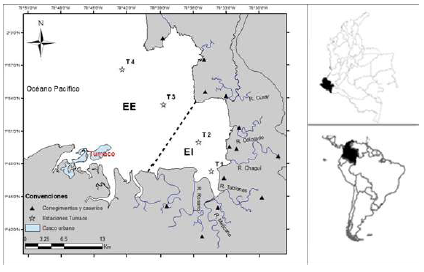
Cabe destacar que el sitio de muestreo se dividió en dos sectores: estuario interno (EI) y estuario externo (EE) (Figura 1). El estuario interno se caracteriza por una mayor influencia de las aguas dulces y sedimentos provenientes de la descarga de los ríos, en tanto que el externo presenta agua más salada y una mayor influencia marina (Piccolo & Perillo, 1997).
Fase de campo
Se hicieron cuatro muestreos en el periodo entre la temporada lluviosa (febrero-marzo) y la seca (octubre-diciembre) de 2020 y 2021 en cuatro estaciones de la bahía de Tumaco (Figura 1). En cada estación se midieron por triplicado los parámetros fisicoquímicos del agua: salinidad (ups), temperatura (°C) y oxígeno disuelto (mg/L), con una sonda multiparamétrica (Thermo Scientific Orion Five Stars), así como la transparencia (cm), con un disco Secchi.
Se obtuvieron muestras biológicas (peces) en cada estación mediante tres artes de pesca artesanales: línea de anzuelo o palangre de fondo, red de arrastre o changa y trasmallo. La línea de anzuelo de 10 mm de calibre, con una distancia entre anzuelos de 1,5 m y de fondo a boya de 1,2 m, situada a 7 m de profundidad, con un tiempo efectivo de pesca de 45 a 50 min. La red de arrastre o changa tenía un ojo de malla de 25,4 mm, un largo de relinga de 6 m y se situó a una profundidad de 12 m en T1 y T2 y 18 m en T3 y T4, con un tiempo efectivo de pesca de 10 a 11 min. El trasmallo tenía un ojo de malla de 50,8 mm, un largo de red de 360 m, y se situó a una profundidad de 3 m, con un tiempo efectivo de pesca de 30 a 60 min. Los peces capturados se separaron en bolsas plásticas rotuladas con el número de la estación y el arte de pesca y posteriormente se almacenaron en neveras con hielo para llevarlos al laboratorio, con el fin de identificarlos y procesarlos.
Fase de laboratorio
Los peces recolectados fueron identificados taxonómicamente siguiendo las claves de identificación de la FAO (Fischer et al., 1995), las bases de datos de Fishbase (Froese & Pauly, 2022) y del Instituto Smithsonian de Investigaciones Tropicales (Robertson & Allen, 2015), y a partir de la revisión bibliográfica. Después se midieron las longitudes total y estándar y el peso total de cada individuo.
Se extrajo el estómago y se conservó en formol al 4 %. Los ítems alimentarios se observaron en el estereoscopio y se clasificaron en las siguientes categorías: crustáceos (Cru), peces (Pe), moluscos (Mol), anélidos (An), detritos (De), microplásticos (MP) y material digerido. Estos ítems se secaron en el horno a 70 °C durante 24 h y posteriormente se contaron y pesaron en una balanza analítica (precisión de 0,0001 g) (Majdi et al., 2018). Para la correcta identificación de microplásticos se realizó una prueba de resistencia térmica.
Tratamiento de datos
Con el fin de determinar la ecología trófica, se calcularon los siguientes índices: composición numérica (CN) (Ec.1), composición gravimétrica (CG) (Ec.2), y frecuencia de presencia de presas (FO) (Ec.3). Además, la importancia de cada presa se determinó mediante el índice de importancia relativa (IIR) (Ec.4) y se determinó el coeficiente de vacuidad (CV) (Ec.5) (Cogua et al., 2013; Duque & Acero, 2003; Tafurt et al., 2021). También se hizo un análisis de varianza multivariante permutacional (permANOVAs) utilizando 9999 permutaciones, con p<0,05 como diferencia significativa, así como un análisis de escalamiento multidimensional no paramétrico (nMDS) transformando los datos (raíz cuadrada) y utilizando las matrices de similitud de Bray-Curtis para establecer diferencias y asociaciones entre la dieta de las especies.
Composición numérica (CN)
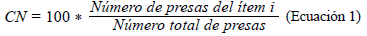
Composición gravimétrica (CG)
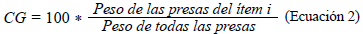
Frecuencia de ocurrencia de presas (FO)
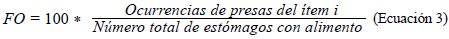
Clasificación de las presas en:
Constantes (presente en más del 50 % de las muestras)
Accesoria (entre 25 y 50 %)
Accidental (menos de 25 %)
Índice de importancia relativa (IIR)

Clasificación de la importancia de las presas en:
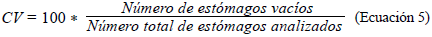
Para evaluar el efecto de la madurez sobre la ecología trófica, los individuos se dividieron según su longitud total en juveniles y adultos, como se observa en las ecuaciones 6 y 7 (Tirado-Ibarra et al., 2018). Para detallar la variación espaciotemporal, se utilizaron el nMDS y el permANOVAs para identificar las asociaciones y los cambios en la dieta de las especies ocurridos entre épocas (lluvia-seca) y áreas (interna-externa).
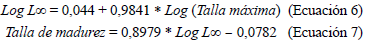
Con el fin de relacionar la ecología trófica con la dinámica ambiental, inicialmente se hicieron nMDS para determinar la agrupación entre la dieta de las especies y los parámetros ambientales del agua. Además, se establecieron modelos aditivos generalizados (GAM) relacionando la respuesta de consumo de cada ítem alimentario con los parámetros fisicoquímicos del agua. Estos modelos son sensibles a asociaciones no lineales entre variables en las que los grados de libertad efectivos (edf) son un indicador del grado de no linealidad; los valores de edf equivalentes a 1 indican una relación lineal y los valores superiores indican relaciones no lineales. La desviación explicada representa la medida generalizada de bondad de ajuste (Hunsicker et al., 2016).
Por último, se detectaron los peces con ingestión de microplásticos, se detalló el estado de madurez, la época y la zona, y se hicieron GAM para relacionar su consumo con los parámetros fisicoquímicos del agua. Asimismo, los microplásticos encontrados se examinaron visualmente, se contabilizaron y se describieron (tipo y color).
Resultados
Ecología trófica
Se evaluaron 467 individuos pertenecientes a 26 especies, de los cuales 93 no presentaron contenido estomacal (CV=19,91 %). Las especies con mayor vacuidad fueron Cynoscion analis (CV=66,67 %), Cynoscion squamipinnis (CV=33,33 %), Stellifer typicus (CV=31,09 %), Stellifer fuerthii (CV=21,74 %) y Stellifer melanocheir (CV=21,05 %) (Tabla 1).
En cuanto a los 374 ejemplares restantes, se encontró que para S. typicus, S. pizarroensis, Larimus argenteus, S. melanocheir y S. oscitans los crustáceos representaron un ítem alimentario muy importante y de consumo constante (IIR>40 % y FO>50 %), siendo su presa principal. Asimismo, los crustáceos fueron la presa principal de S. imiceps, S. illecebrosus, C. analis, S. fuerthii y S. ericymba, sin embargo, también se detectó el consumo de detritos de manera accesoria. Por otro lado, Paralonchurus dumerilii, C. squamipinnis y C. stolzmanni no consumían una única presa principal. En el caso de P. dumerilii, se encontró que consume de manera constante crustáceos (FO=58,33 %), anélidos (FO= 50 %) y detritos (FO=66,67 %), siendo estos ítems de importancia secundaria (IIRcrustáceos= 22,64 %, IIRanélidos=15,31 %, IIRdetritos=14,08 %). Por otro lado, para C. squamipinnis, los peces (FO=62,5 %) y crustáceos (FO=62,5 %) fueron los ítems alimentarios constantes, pero de importancia secundaria (IIRcrustáceos=36,64 %, IIRpeces=25,11 %), en tanto que para C. stolzmanni los peces (FO=40 %) y crustáceos (FO=40 %) fueron de consumo accesorio e importancia secundaria (IIRcrustáceos=16 %, IIRpeces=15,99 %).
Tabla 1 Índices tróficos por especie
| Frecuencia | Importancia | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Constante | Accesoria | Accidental | Alta | Secundaria | Baja | |||||||||
| Especie | na | CV (%) | Cru | Pe | Mol | An | De | MP | ||||||
| FO (%) | IIR (%) | FO(%) | IIR | FO (%) | IIR | FO(%) | IIR(%) | FO(%) | IIR | FO(%) | IIR(%) | |||
| Stellifer typicus | 119 | 31,09 | 93,90 | 84,27 | 20,73 | 1,62 | 1,22 | 17,07 | 0,36 | 9,76 | 0,01 | |||
| Stellifer pizarroensis | 73 | 13,70 | 93,65 | 83,45 | 23,81 | 1,28 | 3,17 | 0,03 | 1,59 | 0,01 | 4,76 | 0,10 | 6,35 | 0,01 |
| Stellifer imiceps | 64 | 7,81 | 96,61 | 86,06 | 15,25 | 0,35 | 1,69 | 0,01 | 5,08 | 0,13 | 27,12 | 1,39 | 1,69 | |
| Larimus argenteus | 48 | 14,58 | 97,56 | 89,06 | 17,07 | 0,42 | 4,88 | 0,02 | 2,44 | 0,01 | 17,07 | 0,47 | 17,07 | 0,06 |
| Stellifer melanocheir | 38 | 21,05 | 96,67 | 89,72 | 6,67 | 0,06 | 3,33 | 0,02 | 3,33 | 0,03 | 10 | 0,10 | 3,33 | 0,02 |
| Stellifer fuerthii | 23 | 21,74 | 100 | 84,76 | 5,56 | 0,08 | 33,33 | 4,62 | ||||||
| Stellifer illecebrosus | 16 | 6,25 | 100 | 86,28 | 6,67 | 6,67 | 0,06 | 26,67 | 2,96 | 20 | 0,34 | |||
| Paralonchurus dumerilii | 13 | 7,69 | 58,33 | 22,64 | 25 | 0,07 | 16,67 | 1,53 | 50 | 15,31 | 66,67 | 14,08 | ||
| Cynoscion analis | 12 | 66,67 | 100 | 99,83 | 25 | 0,04 | ||||||||
| Cynoscion squamipinnis | 12 | 33,33 | 62,50 | 36,64 | 62,50 | 25,11 | 12,5 | 0,01 | 37,50 | 0,40 | ||||
| Stellifer oscitans | 12 | 25,00 | 100 | 98,69 | 22,22 | 0,29 | ||||||||
| Cynoscion stolzmanni | 6 | 16,67 | 40 | 16 | 40 | 15,99 | 20 | 4,00 | 20 | 20 | 0,01 | |||
| Stellifer ericymba | 5 | 100 | 92,31 | 40 | 3,08 | |||||||||
| Stellifer mancorensis | 4 | 25,00 | 100 | 92,70 | 66,67 | 2,65 | 33,33 | 1,11 | ||||||
| Menticirrhus nasus | 3 | 33,33 | 100 | 97,19 | 50 | 1,40 | ||||||||
| Menticirrhus panamensis | 3 | 33,33 | 100 | 54,56 | 50 | 0,10 | 50 | 17,91 | 50 | 4,71 | ||||
| Paralonchurus petersi | 3 | 100 | 69,14 | 33,33 | 6,58 | 33,33 | 3,70 | |||||||
| Cynoscion phoxocephalus | 2 | 100 | 36,82 | 100 | 41,15 | 50 | 11,01 | |||||||
| Cynoscion reticulatus | 2 | 100 | 100 | |||||||||||
| Stellifer chrysoleuca | 2 | 100 | 99,94 | 50 | 0,03 | |||||||||
| Stellifer strabo | 2 | 100 | 56,82 | 50 | 21,59 | |||||||||
| Isopisthus remifer | 100 | 67,50 | 100 | 32,41 | 100 | 0,09 | ||||||||
| Larimus effulgens | 100 | 100 | ||||||||||||
| Menticirrhus elongatus | 100 | 47,54 | 100 | 52,46 | ||||||||||
| Nebris occidentalis | 100 | 99,51 | 100 | 0,49 | ||||||||||
| Paralonchurus peruanus | 100 | 100 | ||||||||||||
a n = número de individuos; IIR = Indice de importancia relativa; FO = Frecuencia de ocurrencia de presas
La mayoría de los crustáceos encontrados en los contenidos estomacales correspondieron a camarones, entre estos se identificó al camarón blanco (Penaeus occidentalis) y al camarón tigre (Riampenaeus byrdi), así como camarones con tenazas. Además, en el contenido estomacal de un individuo de C. squamipinnis se identificó un bagre (Ariopsis simonsi), así como una corvina plateada o cajero (Larimus sp.) en el estómago de un S. imiceps.
En cuanto a los hábitos tróficos, se encontró que la dieta de las especies es diferente (p(PERM)<0,05). No obstante, se observó que los crustáceos y los peces fueron las presas que explicaron en mayor medida las agrupaciones de algunas especies según su nicho alimentario (Figura 2). Con un 80 % de similitud, L. argenteus, S. imiceps y S. typicus compartieron una dieta compuesta principalmente por un elevado consumo de crustáceos. Asimismo, S. fuerthii, S. ericymba y S. pizarroensis compartieron una dieta conformada por crustáceos y un aumento en el consumo de detritos. Además, S. oscitans y S. melanocheir compartieron una dieta constituida por crustáceos, peces y moluscos. Por otro lado, con 60 % de similitud, compartieron nicho C. stolzmanni y C. squamipinnis, con una dieta principalmente piscívora.
Variación espaciotemporal
En la temporada lluviosa se observó una menor diversidad de especies (10), entre las cuales L. argenteus (n=9), S. typicus (n=7), C. analis (n=4), C. stolzmanni (n=3) y C. squamipinnis (n=2) representaron el 80 % de la abundancia. Además, en esta época los crustáceos y los peces fueron las presas con mayor contribución a la dieta de los peces (Figura 3). L. argenteus estuvo presente en ambos sectores, alimentándose principalmente de crustáceos y, en menor medida, detritos. De forma similar, S. typicus consumió crustáceos en ambos sectores, sin embargo, en el externo también exhibió un ligero aprovechamiento de detritos. En contraste, C. squamipinnis y C. stolzmanni compartieron nicho, compuesto por el consumo de peces exclusivamente en la zona interna, aunque C. stolzmanni también presentó un ligero aprovechamiento de moluscos.
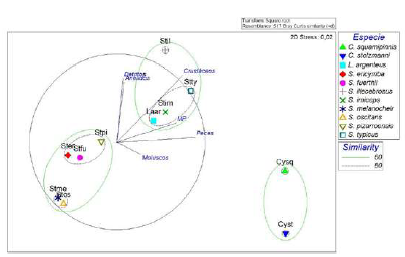
Figura 2 Análisis nMDS de la dieta de algunas especies de la familia Sciaenidae en la bahía de Tumaco. En el gráfico, las especies se denominan con las dos primeras letras del género y la especie.
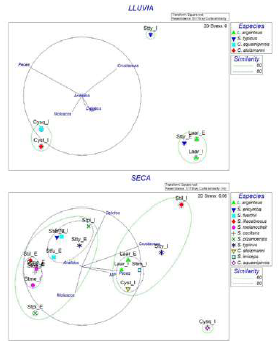
Figura 3 Análisis nMDS de la dieta de algunas especies de la familia Sciaenidae presentes en las temporadas de lluvia y seca. En el gráfico las especies se denominan con las dos primeras letras del género y la especie, así como del sector del estuario (I=interno; E=externo).
En la temporada seca se observó una mayor diversidad de especies (23), de las cuales S. typicus (n=75), S. pizarroensis (n=63), S. imiceps (n=59), L. argenteus (n=32), S. melanocheir (n=30), S. illecebrosus (n=15) y P. dumerilii (n=12) representaron el 80 % de la abundancia. En esta época los crustáceos y los detritos fueron las presas que explicaron en mayor medida la distribución y agrupación de las especies según su nicho alimentario (Figura 3). Por un lado, L. argenteus estuvo presente en ambas zonas y, con un 80 % de similitud, compartió nicho alimentario con S. imiceps y C. stolzmanni en el área interna, caracterizada por un elevado consumo de crustáceos y, en menor medida, de peces. Por otro lado, S. fuerthii, presente en ambas zonas, con un 80 % de similitud, compartió nicho con S. ericymba y S. typicus en el área externa y con S. pizarroensis en la interna, caracterizándose por una dieta de crustáceos y detritos. Además, S. melanocheir y S. oscitans, presentes en ambas zonas, compartieron con una similitud de 80 % nicho alimentario con S. illecebrosus en el área externa; su dieta se compuso de crustáceos y un bajo contenido de detritos.
En el caso de aquellas especies que no compartieron nicho alimentario con otras, o presentaron diferencias en su nicho entre zonas, se encontró que S. typicus se alimentó mayormente de crustáceos en el área interna, en tanto que en S. illecebrosus hubo un mayor aprovechamiento de crustáceos y detritos en la zona interna. En contraste, S. pizarroensis en el área externa estuvo más influenciada por el consumo de moluscos y en la interna, por el de detritos. Por último, C. squamipinnis se diferenció de las demás especies porque presentó un nicho de elevada ingesta de peces.
Como puede observarse en la tabla 1 y la figura 3, el recurso alimentario más importante para la mayoría de las especies de la familia Sciaenidae fueron los crustáceos y, en menor medida, los peces. En cuanto a las variaciones espaciales, se presentaron mayores o menores aprovechamientos de los recursos alimentarios, pero no se encontraron cambios en la presa principal entre sectores. En lo que respecta a las especies presentes tanto en época seca como de lluvia, se halló que la dieta de L. argenteus estuvo compuesta principalmente por crustáceos en ambas temporadas. Por otro lado, S. typicus consumió más crustáceos en temporada seca (23,64 ± 3,57 mg) que en la de lluvias (22,32 ± 10,65 mg) (Tabla 2). En el caso del género Cynoscion, se encontró que C. stolzmanni presentó cambios en la dieta: en temporada lluviosa esta se compuso principalmente de peces (58,55 ± 26,94 mg), en tanto que en la temporada seca primaron los crustáceos (18,90 ± 10,32 mg). Por su parte, C. squamipinnis consumió en ambas épocas principalmente peces, siendo mayor el consumo en la temporada lluviosa (101,20 ± 34,37 mg) que en la seca (58,38 ± 51,72 mg).
En cuanto a la variación temporal en la dieta, se encontró que los adultos de S. typicus presentaron diferencias (p(PERM)<0,05) en el consumo de crustáceos entre épocas, con un mayor aprovechamiento del recurso alimentario en la estación seca (39,78 ± 5,24 mg) que en la de lluvia (30,63 ± 17,37 mg) (Tabla 3).
Relación entre ecología trófica y dinámica ambiental
En época lluviosa se observó que L. argenteus presentó elevados consumos de crustáceos en ambos sectores del estuario, lo que se asoció con aguas de alta salinidad, concentración de oxígeno disuelto y transparencia. Algo similar ocurrió con S. typicus en el sector externo (Figura 4). Por otro lado, C. squamipinnis y C. stolzmanni presentaron un mayor consumo de peces exclusivamente en el área interna, asociado con aguas cálidas y de mayor transparencia, pero de baja salinidad y oxígeno disuelto, características éstas de las aguas más dulces del estuario con influencia de los ríos. En época seca C. squamipinnis también exhibió una tendencia piscívora relacionada con las áreas más internas de aguas cálidas del estuario.
Tabla 2 Variación en el consumo promedio de ítems alimentarios de especies comparables por temporadas.
| Especie | n | Crustáceos (mg) | Peces (mg) | Detritos (mg) | MP (CN) |
|---|---|---|---|---|---|
| Temporada de lluvia | |||||
| Cynoscion squamipinnis | 2 | 101,20 ± 34,37 | 0,15 ± 0,11 | 29,17 ± 4,17 | |
| Cynoscion stolzmanni | 3 | 58,55 ± 26,94 | 0,03 ± 0,03 | 33,33 ± 33,33 | |
| Larimus argenteus | 9 | 21,99 ± 7,30 | 0,04 ± 0,04 | 0,09 ± 0,05 | 6,48 ± 4,34 |
| Stellifer typicus | 7 | 22,32 ± 10,65 | 0,96 ± 0,54 | 4,76 ± 4,76 | |
| Temporada seca | |||||
| Cynoscion squamipinnis | 6 | 16,65 ± 7,45 | 58,38 ± 51,72 | 4,17 ± 4,17 | |
| Cynoscion stolzmanni | 2 | 18,90 ± 10,32 | |||
| Larimus argenteus | 32 | 16,09 ± 2,91 | 0,20 ± 0,13 | 0,23 ± 0,13 | 3,68 ± 1,81 |
| Stellifer typicus | 75 | 23,64 ± 3,57 | 2,81 ± 2,63 | 0,43 ± 0,28 | 3,56 ± 1,34 |
Los valores se presentan como promedio ± error estándar. CN: composición numérica; MP: Microplásticos
Tabla 3 Variación espaciotemporal en la dieta de Stellifer typicus según su estado de madurez.
Los valores se presentan como promedio ± error estándar. Las diferencias significativas se denominaron con letras ordenadas alfabéticamente, siendo la mayor la b. CN: composición numérica; MP: Microplásticos
Por otra parte, en la temporada seca L. argenteus, consumidor de crustáceos, tuvo presencia en aguas de alta salinidad y oxígeno disuelto, condiciones más marinas, en tanto que en especies con dietas mixtas se presentaron las siguientes asociaciones: C. stolzmanni y S. imiceps con una dieta de crustáceos y peces asociada a aguas de baja transparencia, salinidad y oxígeno disuelto. En contraste, S. fuerthii, S. ericymba, S. melanocheir y S. oscitans tuvieron dietas compuestas por crustáceos y detritos, asociados con altos niveles de salinidad, oxígeno disuelto y transparencia.
Con respecto a aquellas especies que presentaron cambios en su nicho alimentario entre áreas, se observó que, en la externa, S. typicus consumió principalmente crustáceos, lo cual se asoció con mayores salinidades y oxígeno disuelto, condiciones propiamente marinas, en tanto que en el área interna su nicho alimentario se caracterizó por un mayor consumo de crustáceos y, en menor medida, de peces, lo que se asoció con menores salinidades y oxígeno disuelto, así como aguas más cálidas. En cuanto a S. illecebrosus, en la zona interna tuvo mayores consumos de crustáceos y detritos asociados a aguas un poco más cálidas y menos transparentes. Además, en el área interna se observó que S. pizarroensis presentó mayor consumo de detritos asociado a una menor transparencia, salinidad y oxígeno disuelto.
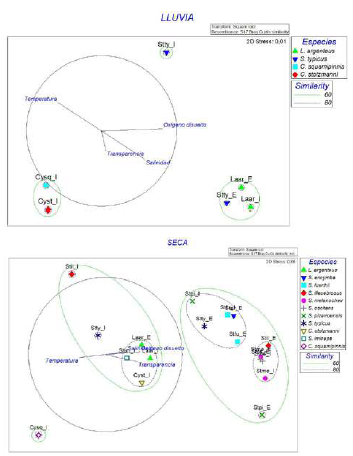
Figura 4 Asociación de la dieta de algunas especies de Sciaenidae con la dinámica ambiental en las temporadas lluviosa y seca. En el gráfico las especies se denominan con las dos primeras letras del género y la especie, así como del sector del estuario (I=interno; E=externo).
En general, cada especie de la familia Sciaenidae presentó una asociación particular entre el mayor aprovechamiento de los recursos alimentarios y las variables ambientales, lo cual indicaría que los peces aprovecharon de manera diferente el recurso, según sus especificaciones fisiológicas y adaptaciones a los microhábitats, haciendo más eficiente la transferencia de biomasa y la energía en el estuario.
En el análisis específico, se determinó que el consumo de crustáceos de S. typicus presentó relaciones significativas no lineales con la temperatura, el oxígeno disuelto y la transparencia (Tabla 4). El mayor consumo de crustáceos se asoció con un aumento de la temperatura de entre 27 y 28 °C (Figura 5a). Con respecto al oxígeno disuelto, el mayor consumo de crustáceos ocurrió en valores mínimos, de 5,5 mg/L, y máximos, de 8,5 mg/L (Figura 5b), en tanto que con valores intermedios de transparencia (70 cm), se presentó el mayor aprovechamiento (Figura 5c). Además, en S. typicus se determinó una relación no lineal entre el consumo de peces y la temperatura, en la cual el mayor consumo de peces sucedió en aguas por encima de 28 °C (Figura 5d). Estos resultados mostraron un cambio en la alimentación de peces y crustáceos direccionado por los cambios en la temperatura, pues en aguas más frías S. typicus se alimentó principalmente de crustáceos (Figura 5a), pero en aguas más cálidas se observó un aumento en el consumo de peces (Figura 5d).
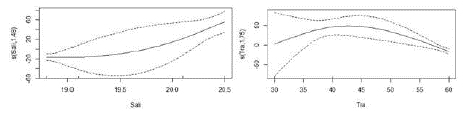
Con respecto a S. imiceps, el consumo de crustáceos se relacionó de manera no lineal con la salinidad y transparencia (Tabla 4) y fue significativamente mayor en aguas más salinas (Figura 6), con transparencias de alrededor de 43 cm. (Figura 6).
En L. argenteus se determinó que el consumo de crustáceos presentó una relación no lineal con la salinidad, la temperatura y el oxígeno disuelto (Tabla 4). El mayor consumo de crustáceos se produjo en las aguas más marinas del estuario, entre los 26 y 28 ups (Figura 7a). Con respecto a la temperatura, el aprovechamiento de crustáceos mostró dos picos, uno en aguas relativamente frías, de 26 °C, y otro en aguas más cálidas, alrededor de 28,5 °C (Figura 7b), lo cual puede estar asociado a las aguas más salinas en diferentes temporadas climáticas. Asimismo, el mayor consumo de crustáceos ocurrió en las aguas con mayor oxígeno disuelto (Figura 7c). Por otro lado, el consumo de peces de S. melanocheir se relacionó significativamente y de forma no lineal con la temperatura y el oxígeno disuelto (Tabla 4). Se encontró una mayor ingesta de peces en áreas con valores intermedios de temperatura (27 °C) y oxígeno disuelto (5,8 mg/L) (Figura 8).
Tabla 4 Resultados del análisis GAM relacionado el consumo de los ítems alimentarios con los parámetros fisicoquímicos del agua.
| Stellifer typicus (n=82) | Stellifer imiceps (n=59) | Larimus argenteus (n=41) | Stellifer melanocheir (n=30) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cru | Pe | MP | Cru | MPb | Cru | MP | Pe | MP | ||
| Salinidad | Edf | 1,08 | 1,877 | (+1.000) | 1,481 | (-1.000) | 4,906 | 2,248 | (-1.000) | (-1.000) |
| F | 2,214 | 1,013 | 0,914 | 13,030 | 0,345 | 7,605 | 1,177 | 0,034 | 0,107 | |
| p | 0,152 | 0,407 | 0,342 | *** | 0,560 | *** | 0,294 | 0,854 | 0,746 | |
| D.e.(%) | 3,03 | 3,8 | 1,31 | 31,8 | 0,598 | 57,9 | 18,8 | 0,123 | 0,38 | |
| Transparencia | Edf | 1,878 | 1,468 | (+1.000) | 1,751 | (+1.000) | 1,994 | 2,781 | 1,763 | (+1.000) |
| F | 3,724 | 0,438 | 0,263 | 11,890 | 0,364 | 0,811 | 3,045 | 2,819 | 0,011 | |
| p | * | 0,499 | 0,609 | *** | 0,549 | 0,399 | * | 0,055 | 0,917 | |
| D.e.(%) | 9,4 | 2,36 | 0,882 | 27,6 | 0,63 | 9 | 31,4 | 21,6 | 0,0376 | |
| Temperatura | Edf | 3,449 | 4,137 | (-1.000) | 1,820 | (+1.000) | 5,288 | (+1.000) | 1,907 | (+1.000) |
| F | 2,488 | 3,192 | 0,223 | 13,930 | 0,215 | 4,600 | 1,538 | 5,308 | 0,057 | |
| p | * | * | 0,638 | *** | 0,645 | ** | 0,222 | *** | 0,813 | |
| D.e.(%) | 14,3 | 20,4 | 0,416 | 31,6 | 0,477 | 49,9 | 0,471 | 30,8 | 0,211 | |
| Oxígeno disuelto | Edf | 2,488 | 2,694 | 1,001 | (+1.000) | (-1.000) | 2,403 | 1,430 | 1,904 | (-1.000) |
| F | 4,758 | 1,750 | 0,014 | 26,160 | 0,261 | 13,200 | 1,457 | 5,461 | 0,091 | |
| p | ** | 0,124 | 0,910 | *** | 0,611 | *** | 0,171 | * | 0,765 | |
| D.e.(%) | 17,1 | 9,97 | 0,266 | 31,5 | 0,462 | 52 | 13 | 29,7 | 0,328 | |
Edf: grados de libertad efectivos (1 significa relación lineal; >1 relación no lineal. Cuando la relación fue lineal se registró entre paréntesis y se puso el signo (+ o -) para representar el sentido de la relación). Los asteriscos representan relaciones significativas de la siguiente forma: p<0,001: ***; p<0,01: **; p<0,05: *; F-value (Estadístico de Fisher); D.e.(%): desviación explicada (%).
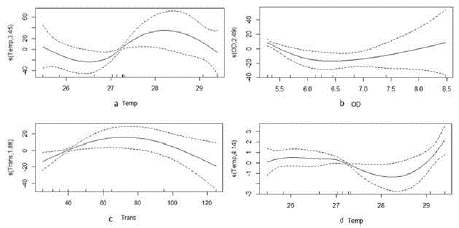
Figura 5 Análisis GAM de la dieta de Stellifer typicus relacionando la respuesta del consumo de crustáceos con las variables predictoras: a. Temperatura (Temp). b. Oxígeno disuelto (OD). c. Transparencia (Trans) y la respuesta del consumo de peces con la variable predictora d. Temperatura
Ingesta de microplásticos
Se detectó consumo de microplásticos en 30 individuos pertenecientes a 10 especies, lo que corresponde al 7 % de la muestra evaluada (Figura 9a). El orden descendente por ingesta de microplásticos fue el siguiente: S. typicus (27 %), L. argenteus (23 %), S. pizarroensis (13 %), C. squamipinnis (10 %) y S. illecebrosus (10 %), en tanto que S. imiceps, S. melanocheir, C. stolzmanni, S. chrysoleuca e I. remifer únicamente registraron un individuo con ingesta de microplásticos (Figura 9a). Entre los microplásticos se encontraron fibras de diversos colores (transparentes, azules, negras, rojas y verdes), gránulos y fragmentos, siendo las fibras transparentes (39 %) y azules (36 %) las más abundantes (Figura 9b). La frecuencia de consumo de microplásticos en todas las especies fue accidental, excepto en C. squamipinnis, en la que fue accesorio (Tabla 1); cabe destacar, que no se detectó su presencia en peces con el estómago vacío.
Por otro lado, se encontró que en S. typicus y L. argenteus el mayor consumo se presentó en individuos juveniles, mientras que en C. squamipinnis, S. illecebrosus y S. pizarroensis se dio en adultos. Las mayores ingestas de microplásticos se presentaron en el área interna y en temporada seca, excepto en C. squamipinnis en que se dieron en época lluviosa. En el caso contrario, en aquellas especies que únicamente presentaban un individuo con ingesta de microplásticos, se encontró que C. stolzmanni, S. chrysoleuca, S. melanocheir eran adultos, mientras que S. imiceps, era juvenil. En todas las especies el consumo se dio en la temporada seca y en el sector interno, excepto en C. stolzmanni, con un consumo preferente en la época de lluvias, y en I. remifer, en la parte externa.
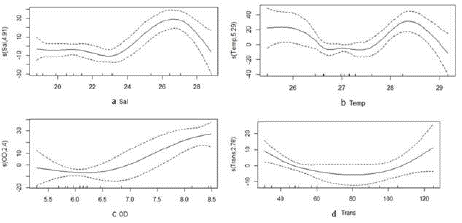
Figura 7 Análisis GAM relacionando la respuesta del consumo de crustáceos de Larimus argenteus con las variables predictoras: a. Salinidad (Sal). b. Temperatura (Temp). c. Oxígeno disuelto (OD) y la respuesta de la ingesta de microplásticos con la variable predictora d. Transparencia (Trans)
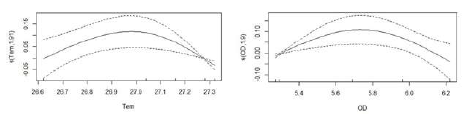
Figura 8 Análisis GAM relacionando la respuesta del consumo de peces de Stellifer melanocheir con las variables predictoras de temperatura (Tem) y oxígeno disuelto (OD)
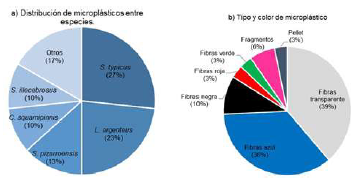
Figura 9 Distribución y tipo de microplásticos hallados en el estómago de individuos de la familia Sciaenidae en la bahía de Tumaco
En cuanto al análisis de las relaciones entre las dinámicas ambientales y la ingesta de microplásticos, se observó que L. argenteus presentó una relación no lineal entre su consumo y la transparencia (Tabla 4), con los mayores consumos en aguas de transparencia extrema, es decir, tanto en aguas de transparencia baja (30 cm), asociadas con las áreas internas de mayor influencia de los ríos, como en las de transparencia alta, asociadas con las externas y más marinas (Figura 7d).
Discusión
Ecología trófica
La familia Sciaenidae es una de las más representativas del Pacífico en cuanto a riqueza de especies y abundancia (Muriel-Hoyos & Carmona-Guerra, 2020). Estos peces, en su mayoría, son visitantes ocasionales, transitorios o dependientes de los estuarios (Feutry et al., 2010), por lo que su entrada y estadía en el estuario está ligada con la reproducción, el refugio o el suministro de alimentos en alguna etapa de su ciclo de vida (Molina et al., 2020). Siendo así, es posible que los hábitos tróficos de estos peces varíen en función de la disponibilidad de recursos alimenticios en el estuario.
En este estudio se encontró que los mayores porcentajes de estómagos vacíos se obtuvieron en C. analis (CV=66,67 %) y C. squamipinnis (CV=33,33 %), lo que coincide con un estudio en el Pacifico colombiano realizado en su congénere Cynoscion phoxocephalus, en el cual se encontró que la mayoría de individuos tenía el estómago vacío (CV=81 %), lo que se explicaría por una alta tasa de digestión o por la escasez de alimento (Rosero et al., 2016). Asimismo, se encontraron altas tasas de estómagos vacíos en S. typicus (CV=31,09 %), S. fuerthii (CV=21,74 %) y S. melanocheir (CV=21,05 %), lo cual sugiere que estas especies pueden estarse alimentando principalmente durante la noche o en las primeras horas de la madrugada (Muriel-Hoyos & Carmona-Guerra, 2020).
En cuanto a aquellos ejemplares que presentaron contenido estomacal, se encontró que el 92 % de las especies evaluadas consumió crustáceos de manera constante. En el 80 % de ellas los crustáceos fueron un alimento de gran importancia. Esto permite afirmar que estos fueron la presa principal de la mayoría de los miembros de la familia Sciaenidae. Estos resultados coinciden con estudios realizados en especímenes del género Stellifer, Larimus y Parolonchurus, como S. melanocheir, S. ericymba, S. fuerthii, S. mancorensis y S. oscitans (Muriel-Hoyos & Carmona-Guerra, 2020), L. breviceps y L. argenteus (Costa et al., 2020; Polo-Silva et al., 2007; Santos et al., 2022; Santos et al., 2016), y Paralonchurus brasiliensis (Santos et al., 2016).
Variación espaciotemporal y su relación con la dinámica ambiental
Con relación a la variación espaciotemporal, se encontró que en la mayoría de las especies hubo mayor o menor aprovechamiento de los recursos alimentarios según el área del estuario o la época; sin embargo, no se presentaron cambios en cuanto a la presa principal. En general, se encontró un mayor consumo de crustáceos en el sector externo del estuario, lo que se asoció a una mayor salinidad, oxígeno disuelto y transparencia.
En el análisis por especie se encontró que L. argenteus no presentó cambios en sus hábitos alimentarios, por lo que puede afirmarse que es una especie selectiva que consume principalmente crustáceos del grupo de los Penaeidae. Además, el consumo de crustáceos presentó una relación no lineal con la salinidad, es decir, a mayor salinidad, mayor consumo, lo que concuerda con lo reportado por Gamboa-García et al. (2018) y Salcedo et al. (2022) en las bahías de Buenaventura y Tumaco, en donde la presencia y abundancia de macroinvertebrados apetecibles para los peces, como el camarón blanco (P. occidentalis) y el camarón tigre (R. byrdi), se asociaron de manera positiva con la salinidad del agua, presentando sus mayores abundancias en el área externa del estuario.
Asimismo, se encontraron algunas relaciones no lineales entre el consumo de crustáceos, el oxígeno disuelto y la transparencia en L. argenteus, S. typicus y S. imiceps, en las cuales, cuanto mayores eran los valores de estos parámetros, mayor era el consumo de crustáceos. Esto reafirma la idea anterior, pues los valores altos de oxígeno disuelto, transparencia y salinidad corresponden a condiciones ambientales de influencia principalmente marina, como las presentes en el área externa, en las cuales la riqueza de macroinverte-brados es mayor debido a la interacción y oferta de microhábitats (Gamboa-García et al., 2018; Salcedo et al., 2022).
Además, fenómenos como la corriente de Humboldt pueden conducir a un aumento en la abundancia de macroinvertebrados, dado que los vientos que viajan hacia el ecuador a lo largo de las costas inducen el afloramiento de masas de agua fría subsuperficiales ricas en nutrientes que alimentan el crecimiento de fitoplancton y, por ende, suponen un aumento en la productividad primaria, así como en toda la cadena trófica. Con base en esto es posible plantear que un mayor consumo de crustáceos en la zona externa del estuario podría asociarse a una mayor abundancia de crustáceos provocada por la corriente de Humboldt (Weidberg et al., 2020). En el caso de S. typicus, no se presentaron cambios en la presa principal, sin embargo, se encontraron diferencias significativas en el consumo de crustáceos en los adultos entre temporadas, siendo mayor el aprovechamiento en la época seca, en tanto que en la estación lluviosa se presentó un ligero aumento en el consumo de peces. Esto puede explicarse por el aumento de las precipitaciones, las cuales generan mayores entradas de agua dulce, lo que proporciona mayores nutrientes para la productividad primaria y secundaria y, por lo tanto, mayores recursos alimentarios para peces pequeños y jóvenes (Pichler et al., 2015).
Por lo tanto, el aumento en el consumo de peces en época lluviosa está ligado a un aumento en la disponibilidad de esas presas. Además, en S. typicus y S. melanocheir se encontró una relación no lineal entre el consumo de peces y la temperatura, es decir, a mayor temperatura, mayor consumo de peces, lo que se explicaría porque la época lluviosa coincide con la influencia del afloramiento de masas de agua fría subsuperficiales ricas en nutrientes (Weidberg et al., 2020), que estarían asociados con las épocas de desove y crecimiento larval, lo que favorecería la abundancia de peces juveniles que pueden ser depredados por estas especies (Pichler et al., 2015).
Por otro lado, en C. stolzmanni se encontró que consumió exclusivamente crustáceos en la temporada seca y peces en la época lluviosa, en lo que coincide con C. squamipinnis, en la cual se observó un aumento del consumo de crustáceos en época seca. Este comportamiento se debe a cambios ontogénicos, dado a que en la época seca los individuos capturados fueron juveniles, mientras que en la lluviosa fueron adultos, por lo que, como se ha constatado en otros estudios en especies del género Cynoscion (Deary, 2020; Rosero et al., 2016), los juveniles tienden a consumir crustáceos mientras que los adultos optan por dietas más piscívoras.
Ingesta de microplásticos
La presencia de microplásticos en el Pacífico colombiano ha sido previamente documentada en agua (Arboleda et al., 2022; Vidal et al., 2021) y en sedimentos (Vásquez-Molano et al., 2021); asimismo, se ha evidenciado la ingesta incidental de microplásticos en peces (Becerra et al., 2022; Herrera et al., 2022; Tafurt et al., 2021). En este estudio se determinó que las mayores ingestas de microplásticos ocurrieron en el área interna del estuario. Este sector es más cercano a la población y, por ende, presentó una mayor intervención antrópica, siendo más propenso a ser contaminado con microplásticos provenientes de las descargas de aguas residuales domésticas vertidas directamente al estuario (Justino et al., 2021) o a la disposición directa de residuos sólidos en los cuerpos de agua costeros de la bahía de Tumaco (Tejada et al., 2003).
En general, el tipo de microplásticos más ingerido fueron las fibras transparentes (39 %) y azules (36 %), lo cual concuerda con otros estudios realizados en peces, en los que las fibras de estos colores fueron el tipo de microplásticos más común en los estómagos analizados (Dantas et al., 2020; Justino et al., 2021; McGregor & Strydom, 2020; Neto et al., 2020; Tafurt et al., 2021; Wright et al., 2013). Estas fibras provienen principalmente de la industria textil, así como del uso, mantenimiento, desgaste y pérdida de artes de pesca (Dantas et al., 2020).
También se observó que en la mayoría de las especies el consumo de microplásticos se presentó en los adultos, excepto en L. argenteus, S. typicus y S. imiceps. Aunque en diversos estudios se ha encontrado que no hay una relación clara entre el tamaño del pez y el consumo de microplásticos (Neto et al., 2020; Wright et al., 2013), es importante considerar que las especies depredadoras son más vulnerables a la contaminación por microplásticos debido a la transferencia trófica que ocurre cuando ingieren una presa contaminada (Justino et al., 2021).
Asimismo, se observó que la transparencia influenciaba el consumo de microplásticos. Por ejemplo, en L. argenteus se encontró una relación no lineal entre la transparencia y el consumo de microplásticos debido a que la profundidad y la turbidez pueden enmascarar los microplásticos, lo que representa un problema para los peces demerso-pelágicos y demersales, ya que puede haber ingestión accidental al buscar alimento, ya que su visión es secundaria a otros sentidos como el olfato (Neto et al., 2020).
Por otra parte, el 70 % de las especies con microplásticos en su contenido estomacal es importante para la subsistencia de la población y del sector comercial, por lo que esta contaminación puede comprometer la sostenibilidad de los recursos pesqueros en el futuro (Dantas et al., 2020) debido a los daños morfológicos, anatómicos, fisiológicos e intestinales que provoca en los peces (Sutton et al., 2016). Además, es un riesgo potencial para la salud humana debido a la biomagnificación de estos microplásticos al absorber contaminantes orgánicos persistentes y transferirlos a través de la cadena trófica (Dantas et al., 2020).
Conclusiones
El estudio permitió determinar que la mayoría de las especies de la familia Sciaenidae en la bahía de Tumaco consumió principalmente crustáceos, lo que los convierte en un recurso alimentario importante que conecta las redes bentónicas y pelágicas. En cuanto a las variaciones espaciotemporales, no se encontraron cambios en la presa principal de los ejemplares entre áreas, por lo que es posible plantear que los hábitos tróficos de los peces de esta familia están influenciados en gran medida por el aprovechamiento de un recurso alimentario común y abundante en la bahía, es decir, los crustáceos. Sin embargo, eventualmente puede presentarse el aprovechamiento de otro tipo de presas abundantes en el estuario según las variaciones estacionales y ambientales, como los peces o detritos. Además, los mayores consumos de crustáceos se presentaron en la zona externa del estuario, asociada a condiciones ambientales mayormente marinas con mayor abundancia y riqueza de macroinvertebrados.
Por otro lado, se detectó la ingesta incidental de microplásticos en el 7 % de los ejemplares evaluados, pertenecientes a 10 especies, 70 % de las cuales es de interés comercial o importante para la subsistencia de la población de Tumaco. Además, el tipo de micro-plástico más ingerido fueron las fibras y los mayores consumos se presentaron en el área interna del estuario, que tiene una mayor influencia antropogénica. Por último, los resultados plantean la creciente preocupación por el futuro de la pesca artesanal en la bahía de Tumaco, dado que la presencia de contaminantes puede comprometer la sostenibilidad de la pesca y generar afectaciones en la salud de peces y humanos.

