











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introduction
The Uncaria (Schreb) genus belongs to the Rubiaceae family and comprises about 60 species with a pantropical distribution (Gomes et al., 2016). Only Uncaria tomentosa (Willd.) DC. and Uncaria guianensis (Aublet) J.F. Gmel. are known in the Amazon region as cat's claw. Both species are widely distributed in humid tropical forests, especially in the Amazon region (Alvarenga, 2010), and they are used in traditional medicine to treat degenerative, infectious, and respiratory diseases and tumors (Garzón, 2019). Uncaria species are known as medicinal herbs commercialized worldwide (Gomes et al., 2016). There are cultivation initiatives, but most of the commercialized raw material comes from the natural environment, which has decreased populations, especially in some areas of the Peruvian Amazon (Domínguez & Castillo, 2007).
The species differ in the leaf, spine and bark morphology: U. guianensis has ovate leaves, strongly twisted hook-like woody spines, and the main stem has a brown outer shell (Zevallos & Tomazello, 2010), while U. tomentosa is characterized by lanceolate leaves, adult spines sparsely curved downward, and yellowish inner bark (Zevallos & Flores, 2003; Cruz et aL, 2011). They generally grow in secondary forests but can also be found in primary forests (Gomes et al., 2016). Although cat's claw species have been reported in various soil types (dystric Cambisols, Fluvisols, and ortic Acrisols), U. tomentosa is more nutritionally demanding than U. guianensis (Domínguez, 1997).
The most studied and commercialized species is U. tomentosa, presumably because it is more abundant than U. guianensis (Gomes et al., 2016). Some reports on the distribution and abundance of U. tomentosa come from Perú (Quinteros, 2001; Domínguez & Castillo, 2007; Canales-Springett et aL, 2013), and both species have been studied in the State of Acre in Brazil (Miranda et al., 2003). There is no additional information from other areas in the Amazon region. The study by Zavala & Zevallos (1996) in Perú is the only one reporting on the soil conditions for their growth and development, and, therefore, edaphological information on U. guianensis is limited. In this context, an interdisciplinary study was conducted with Tikuna indigenous communities to understand both species' distribution and abundance in the southern Colombian Amazon and the influence of soil conditions in natural and disturbed areas. This will help to determine whether U. tomentosds abundance is higher than that of U. guianensis as it happens in other areas of the Amazon region.
Materials and methods
Study area
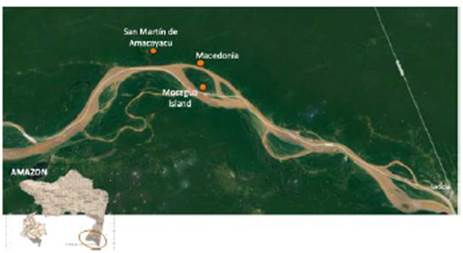
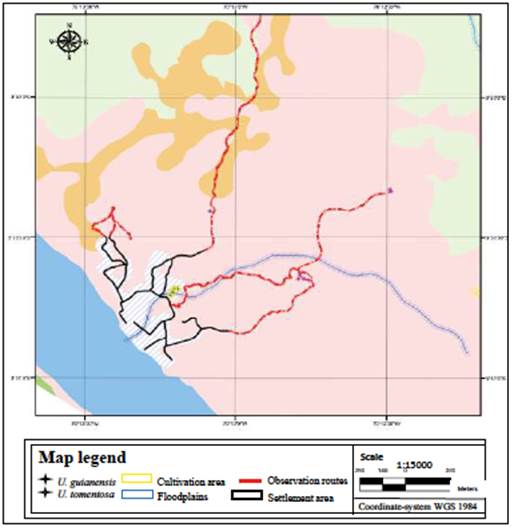
Three areas were selected: one in the indigenous community of San Martín de Amacayacu and the other two in Macedonia. The first area (at 3°46'38"S, 70°18'12"W, on the riverside of the Amacayacu River) has denudation soils, as well as the second one (at 03°53'00"S, 70°11'17"W, on the Amazon River). The third area (at 3°51'21"S, 70°13'19"W), known as Mocagua Island, has alluvial soils (Figure 1).
Methodology
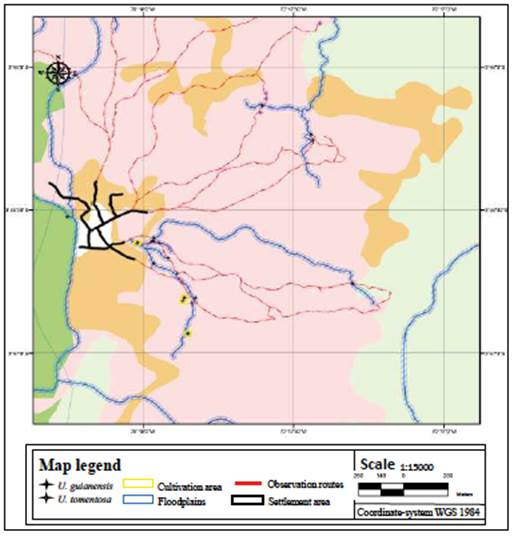
The indigenous people with the best knowledge of cat's claw species were identified through a non-probabilistic chain or snowball sampling (Otzen & Manterola, 2017). Semi-structured interviews were conducted with them to gather information on variables of interest: common names and species identity, areas in their territory where they grow, and associations with other plants. Tikuna translators helped when the interviewee's first language was not Spanish, and the plant's common names were mentioned also in the Tikuna language. Social mapping (Vavilinaa & Skalaban, 2015) helped determine the cat's claw-growing areas in maps made by the participants, including the types of landscapes and soils. With this information, Uncaria species roads and observation routes were defined. Given the terrain conditions and the time required in the communities, an area of influence was established following those roads with lengths between 1 and 7 km; these did not cover every locality but allowed us to estimate the distribution pattern of both species.
Based on this cartography, field trips were made with the community experts to search for plants locally recognized as cat's claws. During the trips, they described in their terms the color of the bark, the shape and color of the leaf, the size of the spines, and the changes in landscape types and soil characteristics. Forty-one samples were georeferenced, 38 corresponding to cat's claw individuals (11 of U. tomentosa and 27 of U. guianensis). There were three samples from other species that some indigenous people indicated as cat's claws: one of Macfadyena unguis-cati (L.) A.H. Gentry (Bognoniaceae) and two of Machaerium sp (Fabaceae). For confirmation, herbarium samples were collected according to the Arnelas et al. (2012) methodology and subsequently deposited and identified at the Colombian Amazon Herbarium - COAH, SINCHI Institute.
For the physicochemical analysis, 500 g of rhizospheric soil was collected at 0-10 cm depth in the sampling points with plants. The description of soil types followed the Peña-Venegas et al. (2015) methodology: indigenous participants mention the characteristics according to the colors and textures observed. Codes were assigned using the Munsell chart of hue, clarity, and purity parameters (Domínguez et al., 2012).
Soil samples were analyzed at the Instituto Agustín Codazzi - IGAC soil laboratory in Bogotá, including granulometry, texture class, pH, exchangeable acidity, aluminum saturation, base saturation, total carbon, available phosphorus, cation exchange content (CEC), total bases, and the presence of calcium, potassium, magnesium, and sodium.
Data analysis
The ATLAS.ti 7.0 software served for coding and analyzing the qualitative information from the interviews, and ArcGIS 10.6.1 for the geographic data. First, the sampling points and the observation routes were digitized, and then the area of influence to determine the abundance of the cat's claw species was calculated. This area totaled 25.34 hectares, including the buffer established on the observation routes with a margin of 5 m on each side of the road.
The level of local knowledge of U. tomentosa and U. guianensis botanical aspects was estimated by the percentage frequency. Species distribution and frequency on the different types of landscapes were also determined. A Kruskal-Wallis non-parametric variance test was performed to establish significant differences with a 5% probability for each soil type, for soils in the study areas and soils associated with the species.
Results
Local identification of Uncaria species
Cat's claw commonly names both Uncaria species in these communities, but "garabato" is also used, albeit less commonly. Indigenous populations in the Peruvian Amazon use it also (López, 2006). However, these communities have specific names to identify each species: U. tomentosa, for example, is known as white cat's claw (u«a de gato blanca in
Spanish and michipatü choü in Tikuna language). The local experts described it as a plant with a green vine and a white inner bark, green, ovate leaves, and a slightly curved spine. U. guianensis is locally called red cat's claw (uña de gato roja, michipatü dauü) and it is identified by its reddish inner bark, reddish leaves on the underside, and fully curled spines (Figure 2). Another classification of these species claims that U. guianensis is the "male" and U. tomentosa is the "female".

Figure 2 Morphological identification of leaf, bark, and spine of Uncaria species. A) Uncaria tomentosa; B) Uncaria guianensis
Three samples from Macedonia identified as a cat's claw belong to another species. Two uña de gato roja samples from Mocagua Island belong to the Machaerium sp. genus (Fabaceae), and an uña de gato blanca belonged to Macfadyena unguis-cati (Bignoniaceae). Although some species reported as cat's claws did not belong to the Uncaria genus, it was evident that the experts are very knowledgeable of the morphological characteristics of the species as 90% of the samples were correctly identified as U. tomentosa and 89% as U. guianensis, which demonstrate locals' expertise in establishing the species location and distribution.
Uncaria species distribution by landscapes
Indigenous communities identified six landscape types where cat's claw species could occur (Table 1). This categorization is based on spatial location, temporal aspects of land use, flooding periods, soil types, vegetation cover, and predominant fauna. According to the degree of intervention and the type of vegetation, cultivated land (chagra - naane in Tikuna language), stubble (rastrojo - íchikü), and primary forest (naimacatüxü) were identified.
Table 1 Local classification of landscapes in the study area
| Common name | Tikuna name | Local description |
|---|---|---|
| Stubble on floodplains | íchikü nibaiü | Young secondary forest area, which floods seasonally or during heavy rains |
| Stubble on highlands | íchikü dauchitacüãã | Highland or upland area with natural secondary forest |
| Cultivation area on floodplains | Naane nibaiü | Planting plots in floodable areas |
| Cultivation area on highlands | Naane dauchitacüãã | Cultivation area on dry land or high altitude |
| Primary forest | Naimacatüxü | Natural forest area on the highland |
| Várzea | Tatúmüànè | Annual flooded area (Mocagua Island) |
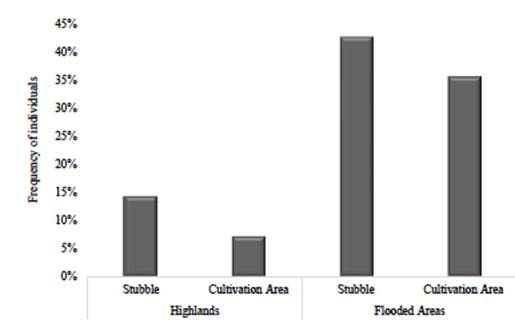
The Tikuna also distinguish two divisions in the physical space: the highland areas (dauchitáààné) and the annual flooded areas (tatüànè). Depending on the type of flooding, this second space can be divided into floodplains during the rainy months (várzea, tatúmüànè) and flooded areas during days of prolonged rains (bajial, nibaiü) (Montes, 2002). Field observations showed that U. guianensis distribution and abundance are higher than U. tomentosa, especially in flooded areas. There were 1.62 cat's claws individuals per hectare and 1.06 U. guianensis individuals in high altitudes and floodplains (Figure 3). In contrast, U. tomentosa abundance was 0.43 individuals per hectare, restricted to secondary forests in highlands (Figure 4). This was the only one observed in mature forests with low intervention.
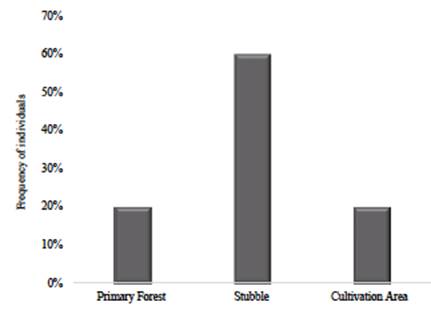
The highest number of U. guianensis plants was found in San Martín de Amacayacu, i.e., 82% of the individuals of this species (Figure 5). U. tomentosa was more frequent in Macedonia (60%), all in the stubble on highlands (Figure 6). None of these species were found on the Mocagua Island.
Regarding the most frequent species in Uncaria habitats, in flooded areas, Scleria secans (Cyperaceae) and Calathea lutea (Maranthaceae) were found with U. guianensis, and in the highlands, Vismia sp. (Hypericaceae) and Cecropia sp. (Urticaceae). In non-flooded areas habitats, Apeiba membranacea (Malvaceae) and Ormosia sp. (Fabaceae) were frequent next to U. tomentosa (Figure 7).

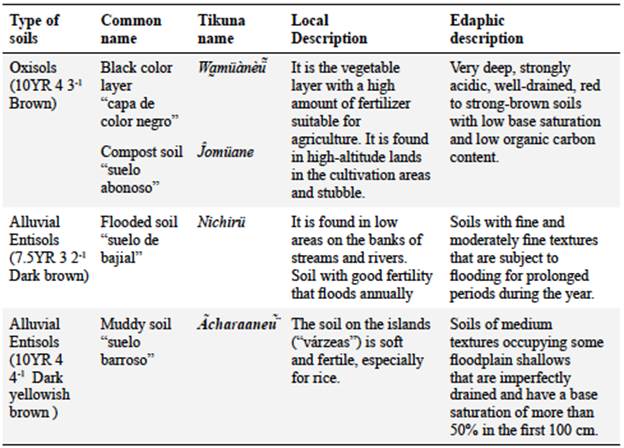
Local identification of soils
Local experts identified four common names for three soil classes in which they believe cat's claw species grow (Table 2): Oxisols and Entisols (according to the USDA nomenclature system). "Black color layer" (capa de color negro - wamüànèü) and "compost soil" (suelo abonoso - jomüane) refer to the same kind of soil found in cultivated areas and in dryland stubble. Commonly, soil types receive different names depending on the characteristics they are associated with. The first name is related to the color and the second to the texture. Their properties are described according to their fertility and location in high and flooded areas during the rainy months (várzeas) or those flooded on days of prolonged rain (bajiales).
Uncaria species distribution by soils
Uncaria guianensis grows in the two soil types evaluated (Table 3): 68% of the U. guianensis individuals were found in alluvial Entisols and 32% in Oxisols on highlands. U. tomentosa was found only in Oxisols on the highlands. According to the soil physicochemical composition in the study areas, the Uncaria species grow in soils with diverse textural classes (Table 4) categorized as highly (pH less than 4.5) or strongly acidic (maximum pH of 5.5), with low to medium available phosphorus ranging from 5.08 to 15.63 mg/kg and medium to high cation exchange capacity (CEC) from 21.65 to 29.71 cmol(+)/kg.
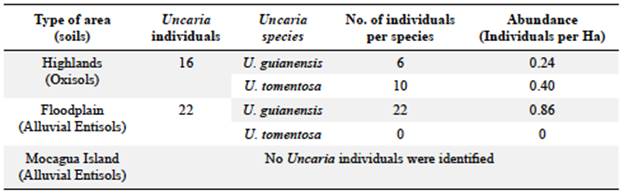
Table 4 Physicochemical composition of soils identified in Tikuna communities in the study area
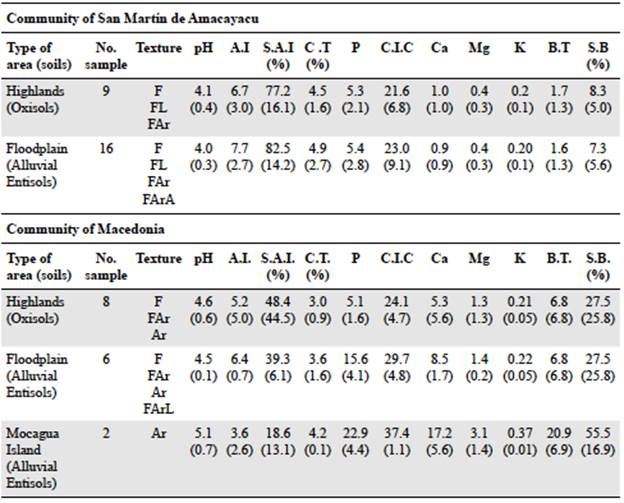
Variables are expressed in mean and standard deviation (SD) in parentheses according to the number of soils collected for each soil type. Texture: F = loam; Ar = clay; FL = silt loam; FAr = clay loam; FArA = sandy clayloam; FArL = silty clay loam; FArL = silty clay loam. pH (1:1 in water); A.I = Exchangeable acidity with KCl in cmol(+) kg-1; S.A.I% = Percentage saturation exchangeable acidity with KCl; C.T% = Percentage total carbón with complete oxidation; P = Phosphorus available by Bray II in mg/kg; C. I.C = Cation exchange capacity with neutral ammonium acetate in cmol(+) kg-1; Ca = Calcium by DTPA in cmol(+) kg-1; Mg = Magnesium by DTPA in cmol(+) kg-1; K = Potassium by DTPA in cmol(+) kg-1; B.T. = Total bases; S.B.% = Percent saturation of bases with neutral ammonium acetate
The statistical results showed a homogeneous physicochemical composition in the corresponding soil types (Table 5). Differences were found in the physicochemical composition of Oxisols and Entisols. The concentration of phosphorus (22.94 mg/kg) and sodium (0.14 cmol(+)/kg) were higher in Entisols. On average, the pH levels in soils where U. guianensis grows were lower than in U. tomentosa.
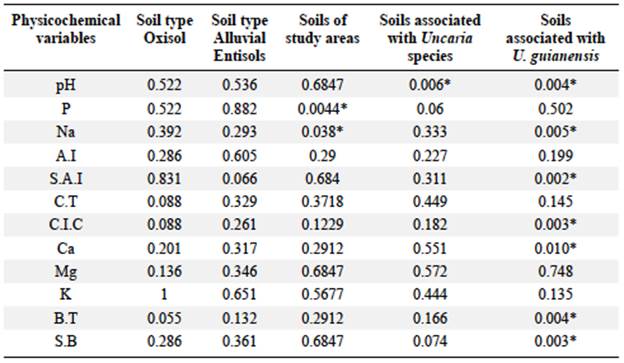
The variability in soil chemical properties is reflected in the significant differences in pH, sodium and calcium levels, the percentage of aluminum saturation, the base saturation, and the cation exchange concentration. U. tomentosa was found only in Oxisols on drylands with a pH between 3.78 and 5.53, exchangeable aluminum variations ranging from 0.15 to 12.62 cmol(+)/kg, and aluminum saturation percentages from 3.04 to 85.42%. U. guianensis grows especially in acidic soils with a pH ranging from 3.43 to 4.62, exchangeable aluminum levels from 4.66 to 12.36 cmol(+)/kg, and aluminum saturation percentages from 29.44 - 95.18%.
Discussion
Local landscape identification
The common names for the Uncaria species are associated with the external characteristics of stems, leaves, and spines. For indigenous experts, the most outstanding feature is the shape of the spine, which resembles the anatomical structure of the claw (patü) of some animals: michipatü [michi - cat (Felis catus)], bauepatü [baue - charapa turtle (Podocnemis expansa)] or daupatü [dau - sparrowhawk (Buteo sp.)]. Although there is no clear explanation for the gender differentiation between these species, some local experts make this distinction based on their healing. U. tomentosa is the "female" and U. guianensis is the "male". They consider that "males" have more medicinal uses and effectiveness in healing. The non- Uncaria species identified here have been reported before in other studies in the Amazon region (Obregón, 1995; Domínguez, 1997). The Machaerium genus, especially Machaerium nyctitans, is more commonly associated with the name "cockspur" (espuela de gallo (Bisby et al., 1994)). Macfadyena unguis-cati had previously been reported in Colombia with the name cat's claw and in Perú with bat's claw (Gallegos, 2017).
In the study communities, Uncaria species are well distinguished, but many species are associated with the name cat's claw in the Amazon region. Domínguez (1997) reported 22 species called cat's claw in Perú belonging to the Bignoniaceae, Caesalpinaceae, Liliaceae, Mimosaseae, Rosaseae, Rutaceae, Sterculeaceae, and Fabaceae families. Beutelspacher & Farrera-Sarmiento (2015) reported 31 species in México where other families (Cactaceae, Cannabaceae, Loginiaceae, Martyniaceae, Nyctaginaceae, Solanaceae, and Verbenaceae) have been found. The number of botanical families related to this common name is evidence of the confusion in identifying their botanical characteristics. The name is used by local peoples in the Amazon region for plants with similar anatomical structures, especially the presence of the spine (Beutelspacher & Farrera-Sarmiento, 2015). The correct determination of cat's claws is crucial for their medicinal use. Obregón (1995) points out that misidentification can lead to social and health problems, as some species may be toxic or may not have the pharmacological properties against specific diseases.
Uncaria species distribution by landscapes
San Martín de Amacayacu has more streams than Macedonia, so there may be more floodable areas that favor U. guianensis development and growth in this territory. Although Uncaria species are not cultivated directly in the chagras, usually remnant plants are left as they are considered medicinal. People sometimes preserve the seedlings that have grown spontaneously in the stubble when prepared for new chagras. Likewise, when these cultivation areas are abandoned to let the soil rest, the plant can grow and continue its propagation, as these species have no cultural relevance for the Tikuna people and are considered invasive.
Uncaria species distribution and abundance patterns are similar to Miranda et al. (2003) reports in the State of Acre, in the Brazilian Amazon, with a higher number of U. guianensis individuals per hectare than U. tomentosa in areas of 200 m.a.s.l. on average. However, U. guianensis abundance in the study area was higher (24.02 individuals per hectare), especially in flooded areas, and its distribution in the southern Colombian Amazon is related to its ability to grow in low areas between 100 and 800 m.a.s.l., near big rivers or second and third order streams. Due to crop rotation in indigenous communities, U. guianensis is commonly found in stubble fields because it usually colonizes the area after planting ceases. Its presence in chagras and stubble fields responds to its ephemeral heliophyte life form. U. guianensis is more susceptible to luminosity levels; as sun rays in open areas reach the soil with medium effectiveness, they promote its development (Zevallos-Pollito et al., 2000).
As for U. tomentosa, some nearby species support for its entanglement and growth in disturbed areas of primary and secondary forests, and its heliophyte life form requires less light consumption to regenerate (Zevallos-Pollito et al., 2000). The abundance of U. tomentosa in the study areas was low compared to those registered by Quinteros (2001) and Canales-Springett et al. (2003) Ucayali and Huánuco forest reserve areas in the Peruvian Amazon. They found 51 individuals per hectare in undisturbed forests and 900 individuals per hectare in mature disturbed forests.
The Macedonia and San Martín de Amacayacu indigenous communities in the Colombian Amazon have high population densities, which puts additional pressure on the abundance and distribution of the natural populations of Uncaria species. Self-consumption agricultural production plots may have implied high levels of intervention in the territory, especially in secondary forests where many wild species are removed, including cat's claws. This anthropic intervention would explain the differences between this and Quinteros' study (2001), which was conducted in areas of undisturbed forest.
Local identification of soils
The four soils identified in the communities are Oxisols and Entisols (USDA classification). Although local experts mentioned that muddy soil (alluvial Entisols) favors plant growth, none of the Uncaria species were found in this soil. These soils correspond to the first layer, i.e., the A horizon, at approximately 15 cm. The Tikuna relate these soils directly to the layer that provides the necessary nutrition for plants to grow because the microbial activity is restricted to the first 20 cm of these soils (Peña-Venegas et al., 2007), which is essential for the species to absorb phosphorus and other nutrients for their growth and development.
Uncaria species distribution by soils
Uncaria tomentosa was identified in loam-textured and clay loam soils. According to Quinteros (2001), the texture does not have a significant effect on the abundance of this species, while Mechán et al. (2007) have reported that U. tomentosa grows better in clay loam soils in the Aguaytía river basin (Ucayali, Perú), as those soils have greater aeration capacity and water availability, which facilitates the absorption of nutrients. Here, this species was the only one observed in primary and secondary forests, but no relevant differences were observed in the composition of the soil samples.
Peña-Venegas & Cardona (2010) reported a close relationship between highlands successional forests biogeochemical cycles in the southern Colombian Amazon. They found that they tend to be more acidic than those of the Ucayali region studied by Domínguez & Castillo (2007). In the acidity measurements, aluminum saturation percentages were above 60% in the Colombian Amazon, which can affect the species' growth by decreasing its capacity to absorb water and nutrients.
The abundance of U. tomentosa (0.40 individuals per hectare) is low compared to the values reported by Quinteros (2001) in the Ucayali region (51 individuals per hectare in undisturbed forests). The author points out that the available phosphorus content (estimated at 3.62 mg/kg) is directly related to the abundance of the species. However, in our study, there was no significant difference in this edaphic variable (5.34 mg kg-1), so other factors must be responsible. U. guianensis is the cat's claw species that grows in the most acidic soils, with higher exchangeable aluminum levels and saturation percentages ranging from 29.44 to 95.18%. These results support the data reported by Zavala & Zevallos (1996) in the Peruvian Amazon, where they concluded that U. guianensis grows in more acid soils than U. tomentosa. This species has a greater capacity to adapt to the variability of physicochemical conditions in soils of secondary forests and is less demanding in nutritional terms than U. tomentosa (Domínguez, 1997).
Although the alluvial Entisols in the study areas tend to be less acidic, there were no significant differences concerning the Oxisols. It cannot be said that pH is conditioning Uncaria growth, and other edaphological characteristics should be considered to explain the changes in these species growth, for example, drainage conditions; when these are good, aeration and organic matter mineralization processes are favored, which facilitates the nutrition of various plant species (Monsalve et al., 2017). Oxisols are highly evolved, deep, and well drained soils, while alluvial Entisols are not so well-evolved and tend to have low depth and poor drainage (IGAC, 2015). The distribution of Uncaria species in these soil types coincides with the report of Zevallos-Pollito et al. (2000) that found U. tomentosa individuals only in well-drained soils, which commonly favor its growth, while U. guianensis was more abundant in poorly drained soils (Quevedo, 1995). More detailed research on the physical, chemical, and biological characteristics of soil types is required to clarify the behavior of Uncaria population distribution.
On the other hand, the physicochemical composition of the soil on Mocagua Island is not a limiting factor for the cat's claw growth. Quevedo (1995) has documented that the dispersal range of these plants is wide as the wind disperses their filiform seeds. The indigenous communities' establishment of plots for agricultural production reinforces the idea that anthropogenic intervention has influenced the population density of Uncaria species in the study areas.
Conclusions
The local Tikuna experts showed good botanical knowledge of the cat's claw species. Theirs is a crucial contribution that facilitates the location and estimation of their abundance and enhances the reliability of their medicinal products. U. guianensis was the most abundant species both in upland and floodplain areas. It is well-adapted to the predominant soil types, and the edaphological differences do not directly affect its distribution. U. tomentosa was found only in mature highlands. The variations in edaphoclimatic conditions throughout the Amazonian territory make it impossible to consider a homogeneous pattern of distribution and abundance for Uncaria species in this region. The abundance and natural regeneration of Uncaria species seem to be adversely affected by the water table and the intervention of the communities in the forests to extract them for medicinal uses. Therefore, it is necessary to implement planting strategies for these species in situ according to habitat and soil types.

