











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Suramérica alberga un tercio de la diversidad de peces de aguas continentales descritas en el mundo (Reis et al., 2016), las cuales, sin embargo, no se distribuyen homogéneamente en el espacio ni entre grupos taxonómicos (Cassemiro et al., 2023). Las cuencas de la periferia continental suramericana presentan una menor riqueza de especies, pero poseen un mayor grado de endemismo (Albert & Reis, 2011). La costa peruana, que se extiende por más de 2000 km y se caracteriza principalmente por su entorno desértico, alberga un total de 62 cuencas hidrográficas (Autoridad Nacional del Agua - ANA, 2013) y presenta una gran variedad de ecosistemas de aguas continentales entre estuarios, humedales y ríos, que son de corto recorrido, torrentosos y, en su mayoría, estacionales, y desembocan en el océano Pacífico (Ortega & Hidalgo, 2008; Ministerio del Ambiente - MINAM, 2019). La biodiversidad de estos ecosistemas se ve amenazada por el incremento de la contaminación y el deterioro de los hábitats debido a las actividades humanas en esta región habitada por el 50 % del total de la población peruana (Instituto Nacional de Estadística e Informática - INEI, 2021).
En los últimos años las investigaciones han enriquecido el conocimiento sobre la ictiofauna de las cuencas costeras peruanas, especialmente de la costa norte del Perú (Meza-Vargas et aL, 2022; Siccha-Ramírez et al, 2023; Valenzuela et al., 2023). La información sobre la mayoría de las cuencas sigue siendo limitada, principalmente de aquellas ubicadas en el centro y el sur. En esta área se destaca la cuenca del río Pisco, uno de los principales del departamento de Ica, que abastece a la población a lo largo de su recorrido. La parte baja de esta cuenca se utiliza para el consumo humano y la ganadería y la agricultura de exportación, las cuales constituyen la principal actividad socioeconómica de este valle (Instituto Nacional de Recursos Naturales - Inrena, 2003). Además, los humedales aledaños tienen importancia turística y son áreas de refugio y alimentación de una gran variedad de especies (ANA, 2018). Como la gran mayoría de ambientes acuáticos en la costa peruana, infortunadamente el área se ve amenazada por las descargas de residuos sólidos, aguas servidas urbanas y la presencia de especies exóticas (Ortega et al., 2012; ANA, 2016).
Se han publicado registros de la flora y la fauna de esta cuenca (ANA, 2018), pero la información sobre la fauna íctica actual aún es muy limitada. Fowler (1945) registró a la especie Mugil cephalus en el cauce principal del río Pisco. Posteriormente, ProNaturaleza (2010) realizó el registro de especies en el humedal Pisco Playa (bajo río Pisco), usando sus nombres comunes: "tilapia" (Coptodon rendalli y Oreochromis sp.), "guppy" (Poecilia sp.) y "monengue" (Dormitator latifrons).Faustino-Fuster et al. (2017) evaluaron la relación entre longitud y peso de peces en los ríos Pisco y San Juan (provincia de Pisco), entre ellos, Trichomycterus punctulatus, Basilichthys archaeus, Andinoacara stalsbergi y Poecilia reticulata. En los humedales costeros de Pisco ANA (2018) registró las especies M. cephalus, Cyprinus carpio, A, stalsbergi, Poecilia sp. y Oreochromis sp. Actualmente no existe una lista taxonómica de la fauna íctica de esta importante cuenca costera.
En este contexto, el conocimiento de la composición de especies de peces es fundamental para la elaboración de planes de gestión y conservación de la biodiversidad del área evaluada. Nuestro objetivo en este estudio fue determinar la composición de la fauna íctica del bajo río Pisco y proporcionar información sobre su presencia en los ambientes acuáticos evaluados y su estado de conservación frente a las principales amenazas.
Area de estudio
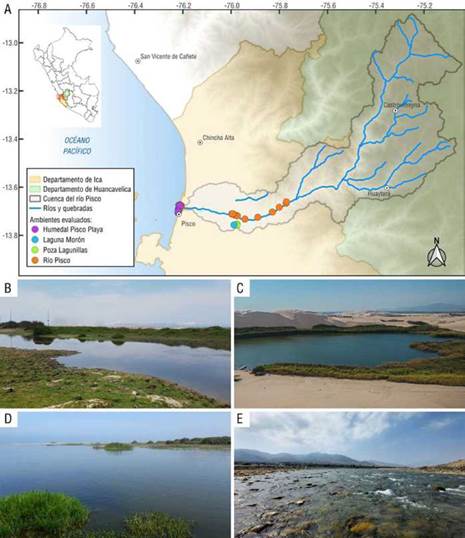
El río Pisco se origina en la laguna Pultoc altoandina (departamento de Huancavelica) y desemboca en la vertiente del océano Pacífico cubriendo una área de 4.122,4 km2 de nororiente a suroccidente y altitudinalmente desde el nivel del mar hasta los 5.218 m (Gamarra et al., 2017). La parte baja del río pertenece a la zona costera, en la provincia de Pisco (departamento de Ica) entre 0 y 250 m s.n.m. (ANA, 2018). Presenta un clima subtropical árido con escasas precipitaciones anuales (0,60 a 1,40 mm). El río Pisco posee un cauce de características meándricas que recorre los distritos de Bernales, Santa Isabel, Independencia, Costa Rica, La Villa, Tambo, Hualla Grande, Pisco y La Alborada, representando el 21,5% del área total de la cuenca (INRENA, 2003)
En este estudio se recogió la información de evaluaciones hidrobiológicas en 34 puntos de muestreo localizados en el bajo río Pisco, que incluye el cauce principal del río y los humedales aledaños (Figura 1). Los humedales de la parte baja son considerados oasis por los valiosos recursos hídricos en esta región desértica. El humedal Pisco Playa se extiende a lo largo del margen costero de la ciudad de Pisco, con una superficie de 8,45 km2 al norte y 15,29 km2 al sur (Instituto Geofísico del Perú - IGP, 2022). La laguna Morón comprende 0,18 km y se encuentra a una altitud de 294 m s.n.m; debido a su contenido salino, se la clasifica como una laguna salobre, con pH neutro; la laguna se originó a partir de la filtración de los cultivos agrícolas y de los acuíferos (ANA, 2018). No hay información detallada sobre la poza Lagunillas, pero forma parte de una red de humedales de pequeña extensión en el margen izquierdo del río Pisco que se originaron por filtraciones de agua subterránea (IGP, 2022).
Metodología
Para la actualización taxonómica se examinaron especímenes depositados en la Colección Ictiológica del Museo de Historia Natural de la Universidad Nacional de San Marcos (MUSM) en Lima, recolectados durante expediciones realizadas entre 1952 y 2020 (Tabla S1,https://www.raccefyn.co/index.php/raccefyn/article/view/2563/4025). La identidad taxonómica de los especímenes fue corroborada en la literatura especializada (Eigenmann, 1922; Eigenmann, 1928; Trewavas, 1983; Arredo, 1989; Schmitter-Soto, 2006; Nelson, 2016), en descripciones originales (Musilova et al., 2009), así como información de la literatura gris (ProNaturaleza, 2010; ANA, 2018). La lista taxonómica se clasificó en orden filogenético según Betancur-R et al. (2017) y la nomenclatura taxonómica se validó siguiendo a Fricke et al. (2023).
Se analizó la riqueza de especies agrupadas en familias y mediante un diagrama de Venn-Euler se visualizó la distribución de dicha riqueza en cada ambiente. Se comparó la composición taxonómica de cada ambiente considerando el porcentaje del total de especies en un gráfico de barras. El recambio de especies en los ambientes se analizó mediante la disimilitud de Jaccard representada en un dendrograma. Las especies se clasificaron de acuerdo con su tolerancia a la salinidad en primarias (dulceacuícolas que son intolerantes al agua salada), secundarias (dulceacuícolas que presentan tolerancia a la salinidad) y periféricas (peces marinos que pueden estar asociados a ambientes estuarinos y ríos) (Myers, 1949; Jiménez-Prado et al., 2015). En cuanto a su origen biogeográfico, se clasificaron en endémicas, nativas y exóticas. La categoría de amenaza de las especies se evaluó según la IUCN (2022). Por último, se analizó el registro acumulado de especies a lo largo del tiempo según la fecha de muestreo de los lotes del MUSM o su aparición en la bibliografía. Los datos se analizaron y representaron gráficamente utilizando el programa R (R Core Team, 2023) y los paquetes ggplot2, ggvenn y microeco.
Resultados
Desde el primer registro de M. cephalus hecho por Fowler en 1945, pasaron más de 40 años hasta el segundo registro de otra especie en la ictiofauna del río Pisco. A partir de 2001, los nuevos registros han aumentado, incluyendo los de especies exóticas (Figura 2A) (Suplementario 2,https://www.raccefyn.co/index.php/raccefyn/article/view/2563/4025).
Riqueza por familias
Se registraron 13 especies de peces distribuidas en diez géneros, ocho familias y ocho órdenes (Tabla 1,figura 2B yfigura 3). La familia con mayor riqueza fue Cichlidae, con cuatro especies, seguida por Poeciliidae, con tres especies. Las familias con menor riqueza específica fueron Atherinopsidae, Lebiasinidae, Trichomycteridae, Mugilidae, Cyprinidae y Eleotridae, representadas cada una por una especie (Figura 2B).
Composición de especies según el ambiente evaluado
El humedal Pisco Playa, río Pisco y la laguna Morón destacaron como los ambientes con mayor riqueza, con nueve, ocho y siete especies, respectivamente (Tabla 2,figura 2C y D). Por otro lado, en la poza Lagunillas se encontraron tres especies (Figura 2C y D). Además, B. semotilus solo se registró en el río Pisco (Tabla 2,figura 2C). Es relevante señalar que A. stalsbergi, P. reticulata y Oreochromis sp. se encontraron en todas las estaciones (Tabla 2,figura 2C y D). Asimismo, es notable la presencia de M. cephalus en tres de los cuatro ambientes evaluados. Las especies restantes estaban presentes en uno o dos ambientes.
Recambio de especies
Los ambientes se dividieron en dos grupos según la disimilitud en el índice de Jaccard (Figura 2E). El río Pisco y el humedal Pisco Playa integraron el primer grupo al compartir seis especies, en tanto que en el otro grupo la laguna Morón y la poza Lagunillas compartieron tres especies.
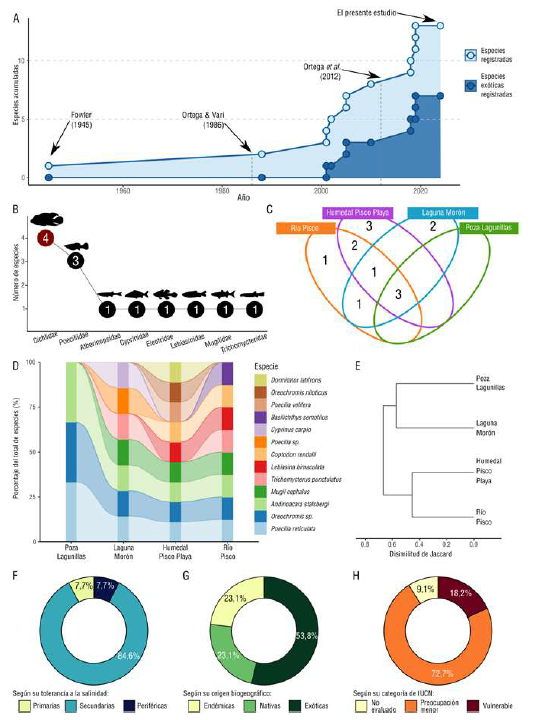
Figura 2 (A) Riqueza de especies por familia en la zona baja del río Pisco. (B) Diagrama de Venn-Euler que representa el número de especies por ambiente evaluado. (C) Gráfica de barras a nivel de especie para cada ambiente evaluado. (D) Dendrograma basado en la disimilitud de Jaccard de los ambientes evaluados. (E) Distribución de las especies según su tolerancia a la salinidad. (F) Distribución de las especies según su origen biogeográfico. (G) Distribución de las especies según su estado de conservación. (H) Acumulación de especies registradas en la cuenca del bajo río Pisco
Especies según su tolerancia a la salinidad
Por su tolerancia a la salinidad, se observó que B. semotilus fue la única catalogada como dulceacuícola intolerante al agua salada (primaria), con el 7,7 % de la riqueza total. Por otro lado, 11 especies (84,6 % de la riqueza) exhibieron cierta tolerancia a la salinidad (secundarias) y únicamente M. cephalus (7,7 % de la riqueza) se catalogó como periférica, es decir, conformada por peces marinos con capacidad para habitar en entornos estuarinos y de agua dulce (Tabla 1yfigura 2F).
Tabla 1 Lista de especies de peces registradas para la zona baja del río Pisco. LC: preocupación menor; NT: casi amenazado, VU: vulnerable, NE: no evaluado.
| Taxón | Nombre común | Tolerancia | IUCN | Referencia/ Voucher |
|---|---|---|---|---|
| Cypriniformes (1) | ||||
| Cyprinidae (1) | ||||
| Cyprinus carpio Linnaeus 1758 | Carpa común | S | VU | ANA, 2018 |
| Characiformes (1) | ||||
| Lebiasinidae (1) | ||||
| Lebiasina bimaculata Valenciennes 1847* | Charcoca, guavina | S | LC | MUSM 70422, MUSM 70434 |
| Siluriformes (1) | ||||
| Trichomycteridae (1) | ||||
| Trichomycterus punctulatus Valenciennes 1846 | Life | S | NE | MUSM 1995, MUSM 18792, MUSM 18793, MUSM 18794, MUSM 20297, MUSM 20302, MUSM 24960, MUSM 24977, MUSM 24979, MUSM 35214 |
| Cichliformes (4) | ||||
| Cichlidae (4) | ||||
| Andinoacara stalsbergiMusilová, Schindler & Staeck 2009 | Mojarra | S | LC | MUSM 18796, MUSM 20298; MUSM 24969, MUSM 24972, MUSM 24976, MUSM 24980, MUSM 24981, MUSM 25535, MUSM 70426, MUSM 70438. |
| Coptodon rendalli (Boulenger 1897) | Tilapia de pecho rojo | S | LC | MUSM 70439, MUSM 70443, MUSM 71298 |
| Oreochromis niloticus (Linnaeus 1758) | Tilapia del Nilo | S | LC | MUSM 70428, 70440, MUSM 70444 |
| Oreochromis sp. | Tilapia | S | MUSM 24967, MUSM 70430, MUSM 70436, MUSM 70441 | |
| Atheriniformes (1) | ||||
| Atherinopsidae (1) | ||||
| Basilichthys semotilus (Cope 1874) | Pejerrey de río | P | LC | MUSM 22416 |
| Cyprinodontiformes (3) | ||||
| Poeciliidae (3) | ||||
| Poecilia reticulata Peters 1859 | Gupi | S | LC | MUSM 18795, MUSM 24968, MUSM 24973 MUSM 24974, MUSM 25537, MUSM 25538, MUSM 25541, MUSM 25542, MUSM 70424, MUSM 70431 |
| Poecilia velifera (Regan 1914) | Velifera | S | VU | MUSM 70427, MUSM 70429, MUSM 70432 |
| Poecilia sp. | Gupi | S | ANA, 2018 | |
| MugUiformes (1) | ||||
| Mugilidae (1) | ||||
| Mugil cephalus Linnaeus 1758 | Liza | Pe | LC | MUSM 1174, MUSM 1956, MUSM 70421, MUSM 70425 MUSM 70433, MUSM 70435, MUSM 70437, MUSM 70442 |
| Perciformes (1) | ||||
| Eleotridae (1) | ||||
| Dormitator latifrons (Richardson 1844) | Monengue, chalaco | S | LC | ProNaturaleza, 2010 |
P: primario, Pe: periférico, S: secundario. (*) = Nuevo registro
Especies según su origen biogeográfico
Se registraron tres especies endémicas, tres nativas y siete exóticas (Tabla 1,figura 2G). Las especies exóticas pertenecen a los géneros Cyprinus, Coptodon, Oreochromis y Poecilia y representaron el 53,8 % de la riqueza total. Además, es relevante destacar que L. bimaculata constituye un nuevo registro para la ictiofauna del río Pisco.
Estado de conservación
Diez especies (90,9 % de la riqueza reportada) se categorizaron según los criterios de la IUCN (Tabla 1,figura 2H); de ellas, cinco eran especies exóticas, tres nativas (L. bimaculata, D. latifrons y M. cephalus) y dos endémicas (A. stalsbergi y B. semotilus), estas últimas catalogadas como de preocupación menor (LC) (Tabla 1,tabla 2).
Discusión
En 1945, Fowler publicó el catálogo sistemático "Peces del Perú", documentando 724 especies de peces en ambientes marinos, costeros, andinos y amazónicos. Nuestra área de estudio se encuentra en el ambiente costero del bajo río Pisco, donde hemos identificado 13 especies. Sin embargo, Fowler solo documentó M. cephalus en el río Pisco.
Es importante tener en cuenta que, en esos años, los especímenes más estudiados eran los procedentes de ambientes marinos. Por ejemplo, en 1907 y 1908 Coker clasificó 500 variedades de peces marinos en el litoral peruano. Fue solo en años posteriores cuando se dio mayor énfasis al estudio de peces continentales, con el registro de 736 especies de peces nativas y exóticas en Perú (Ortega & Vari, 1986), una lista que siguió actualizándose en los años siguientes.

Figura 3 Especies registradas en la zona baja del río Pisco. (1) Especies endémicas: A, B y E. (2) Especies nativas: C y D. (3) Especies exóticas: F-J. Escala = 1 cm
Tabla 2 Ictiofauna según su origen y el ambiente muestreado en la zona baja del río Pisco
| N° | Origen/Especie | Río Pisco | Humedal Pisco Playa | Laguna Morón | Poza Lagunillas |
|---|---|---|---|---|---|
| Endémica | |||||
| 1 | Andinoacara stalsbergi | X | X | X | X |
| 2 | Basilichthys semotilus | X | |||
| 3 | Trichomycterus punctulatus | X | X | ||
| Nativa | |||||
| 4 | Dormitator latifrons | X | |||
| 5 | Lebiasina bimaculata | X | X | ||
| 6 | Mugil cephalus | X | X | X | |
| Exótica | |||||
| 7 | Coptodon rendalli | X | X | ||
| 8 | Cyprinus carpio | X | |||
| 9 | Oreochromis niloticus | X | |||
| 10 | Oreochromis sp. | X | X | X | X |
| 11 | Poecilia reticulata | X | X | X | X |
| 12 | Poecilia velifera | X | |||
| 13 | Poecilia sp. | X | |||
Aunque la introducción de especies exóticas en Perú comenzó en 1921 con P. reticulata (Cossíos, 2010), en la Colección Científica MUSM la muestra más antigua de una especie exótica registrada en el río Pisco es Oreochromis sp., depositada recién en 2005. En cuanto a las especies nativas, la recolección de ejemplares distintos al documentado por Fowler no ocurrió sino hasta 1988.
En el bajo río Pisco, ubicado en la costa central del Pacífico peruano, hay dos tipos de ecosistemas: 1) el desierto costero, compuesto principalmente por suelos arenosos y caracterizado por su aridez y la presencia de extensas áreas desprovistas de vegetación, y 2) el humedal costero, que abarca superficies de agua permanentes o no, de origen natural o artificial, que albergan comunidades biológicas distintivas (MINAM, 2019) y que se clasifican en manglares, lagunas, estuarios, albuferas, deltas, oasis y pantanos (ANA, 2018). La diversidad de la ictiofauna en las cuencas costeras exhibe una limitada riqueza de peces en aguas continentales, influenciada también por las invasiones de especies marinas (Winemiller et al., 2008).
El principal resultado del estudio fue la actualización de la riqueza íctica en el bajo río Pisco, con el registro de 13 especies, de las cuales solo Oreochromis sp. se identificó a nivel de género. Las especies del género Oreochromis tienden a la hibridación (Genner & Turner, 2012; Ford et al., 2019), lo que produce especímenes con características morfológicas intermedias difíciles de identificar correctamente.
Como una nueva incorporación de especie no exótica en esta parte de la cuenca se registró L. bimaculata. Esta especie presenta una distribución que, según Fowler (1945) y Jiménez et al. (2015), abarca desde la costa occidental del Perú hasta Ecuador. Además, dentro del territorio peruano Fowler (1945) cita su presencia desde el río Rímac (región Lima, localidad tipo) hasta el río Zarumilla (región Piura); posteriormente, Valenzuela et al. (2023) la registraron en el río Tumbes. Así, la distribución previamente conocida de L. bimaculata en la costa peruana abarca desde la región de Tumbes (costa norte) hasta la región de Lima (costa central). Este nuevo registro implicaría la expansión de su rango de presencia hasta la costa sur peruana (río Pisco), lo que no se habría detectado previamente debido a la dificultad de realizar estudios en ríos torrentosos y a los pocos monitoreos en la zona, tal como lo demuestra la información de los lotes de la Colección Ictiológica MUSM revisados en este estudio (Suplementario 2,https://www.raccefyn.co/index.php/raccefyn/article/view/2563/4025), lo que resalta la importancia de incrementar los esfuerzos para estudiar la fauna íctica en otros ríos costeros, en especial los de la región sur. Otra posibilidad es que la presencia de L. bimaculata en el río Pisco sea producto de acciones intencionales de los pobladores, pues la especie podría usarse como controladora natural de mosquitos a causa de su dieta insectívora, lo cual tiene antecedentes en el alto Marañón (región Cajamarca), donde la especie pudo haber sido introducida más allá de su rango de distribución natural por pobladores preincas, y logró adaptarse a las condiciones de elevada altitud por su capacidad de sobrevivir en condiciones de estrés, como el bajo nivel de oxígeno en el agua (Reis et al., 2003). Esta segunda hipótesis es menos probable, ya que no hay evidencia del traslado de la especie entre los ríos costeros donde se distribuye naturalmente.
El presente estudio registra una riqueza superior a la reportada por Fowler (1945), Musilová et al. (2009), ProNaturaleza (2010), Vera et al. (2013), Faustino-Fuster et al. (2017) y ANA (2018), que registraron en conjunto ocho especies (M. cephalus, A. stalsbergi, T. punctulatus, B. semotilus, C. carpio, D. latifrons, Poecilia sp. y Oreochromis sp.), así como a la de los ríos de la costa central peruana, donde Ortega et al. (2012) registraron siete especies. Sin embargo, la diversidad registrada en nuestro estudio es menor que la de los ríos costeros del norte peruano, donde se han documentado 27 especies en el río Chira (Meza-Vargas et al., 2022) y 47 en el río Tumbes (Siccha et al., 2023; Valenzuela et al., 2023). La disminución de la riqueza en la franja costera central sería atribuible principalmente a factores geológicos (formación de los Andes y paleohidrografía de Amazonas - Guayas de 20-40 millones de años), según Albert & Reis (2011) y, en segundo lugar, a factores ambientales. Elementos como el paisaje regional, las características ecológicas, las precipitaciones y la cubierta forestal también son variables que influyen en la diversidad local (Reis et al., 2016).
Con respecto a la riqueza según el ambiente evaluado, a pesar de que los peces han logrado colonizar prácticamente todos los ecosistemas acuáticos que muestran cierto grado de permanencia (Helfman et al., 2009), actualmente hay poca información sobre qué especies ícticas están presentes en los diversos hábitats de la costa peruana. Este vacío de información dificulta una comprensión completa de la diversidad de la ictiofauna y sus interacciones en los ecosistemas acuáticos de la región. El humedal costero Pisco Playa posee la mayor riqueza (nueve especies), cifra superior a la registrada en el humedal de Puerto Viejo (Lima), que cuenta con siete especies (García et al., 2021), pero inferior a la cantidad reportada por Castro et al. (1998) en el Refugio de Vida Silvestre Pantanos de Villa (Lima), donde se identificaron 13 especies. En el río Pisco se han registrado ocho especies de peces, y se ha determinado que B. semotilus utiliza este hábitat de manera exclusiva, registros previos en otros ríos de la costa como Vitor y Chira (Fowler, 1945; Meza-Vargas et al., 2022) y la ausencia bibliográfica que indique la presencia de B. semotilus en humedales o lagos, evidencia su especializacion a la vida en ambientes lóticos. En la poza Lagunillas se identificaron tres especies, entre ellas, A. stalsbergi, siendo esta la única endémica descrita en la región.
En cuanto a la composición de especies en los ambientes evaluados, A. stalsbergi, P. reticulata y Oreochromis sp. se registraron en los cuatro ambientes acuáticos evaluados y se las podría considerar como especies núcleo en la cuenca baja del río Pisco, ya que se caracterizan por estar bien distribuidas en sus respectivos hábitats (alta prevalencia) y por ser abundantes y comunes (Pandit et al., 2009; Magurran et al., 2011).
Con respecto al recambio de la composición de especies, tanto el río Pisco como el humedal Pisco Playa presentaron un porcentaje de disimilitud menor a 45 %. Estos ambientes tienen una mayor variación espacial, lo que les permite albergar una mayor diversidad; además, su proximidad facilita la presencia de especies compartidas. El humedal Pisco Playa es un sistema complejo que por su cercanía al ambiente marino puede albergar especies con características ecológicas afines, en tanto que el río Pisco tiene una heterogeneidad de hábitats (orillas amplias y vegetación ribereña) y, por ende, un mayor número de especies. En el caso de la poza Lagunillas y la laguna Morón, que se agruparon con un porcentaje de disimilitud menor al 60 %, sus ambientes están separados y no hay conectividad entre ellos casi en ningún periodo del año; esta condición de sistemas cerrados limita el ingreso y egreso de peces y por ello es menor su posibilidad de compartir especies que, además, toleren las condiciones limnológicas de dichos ambientes.
En cuanto a la tolerancia a la salinidad, en el bajo río Pisco B. semotilus, con hábitos dulceacuícolas exclusivos, fue la única especie primaria que se encontró. Por otra parte, se registraron 11 especies secundarias (84,6 % de la riqueza total) pertenecientes a las familias Cyprinidae, Poeciliidae, Cichlidae, Lebiasinidae, Trichomycteridae y Eleotridae. A pesar de que Meza-Vargas et al. (2022) y Valenzuela et al. (2023) clasifican a L. bimaculata y A. stalsbergi como especies primarias, en este estudio las catalogamos como secundarias porque fueron registradas en el humedal Pisco Playa, el cual ha sido clasificado como salobre (ANA, 2018). Por su parte T. punctulatus se registró en la laguna Morón, también clasificada como salobre por la ANA (2018). Estas tres especies también se han reportado en el refugio de vida silvestre Los Pantanos de Villa (Castro et al., 1998), descrito, asimismo, como un entorno salobre (León et al., 1995). En consecuencia, se puede afirmar que estas tres especies son tolerantes a ambientes con salinidad variable. En cuanto a los aspectos ecológicos que podrían favorecer la adaptación a estos ambientes, se destaca que A. stalsbergi muestra una notable adaptación a aguas alcalinas y duras, ricas en minerales disueltos, e incluso tolera agua salobre (Lüling, 1973). Por su parte L. bimaculata puede encontrarse en aguas estancadas, remansos, quebradas, humedales y pozas, tolerando altos niveles de contaminación, con poco oxígeno e, incluso, puede salir de los esteros a tierra firme (Jiménez-Prado et al., 2015). La familia Trichomycteridae se ha reportado en una gran diversidad de hábitats y ha demostrado tener un gran potencial para colonizar arroyos de altitud (4,000 m. s.n.m), islas de alta mar, aguas freáticas (subterráneas) y aguas termales cálidas o frías (Van der Sleen & Albert, 2018). Otra especie secundaria registrada fue D. latifrons, que puede habitar aguas salobres y corrientes turbias cercanas al mar (Miller, 1966). Presenta resistencia fisiológica a bajos niveles de oxígeno, así como a las variaciones de salinidad y temperatura (Ancieta & Landa, 1977). Además, D. latifrons es una especie anfídroma, lo que implica una migración de individuos en estadio larval hacia el mar poco después de la eclosión de los huevos para alimentarse y crecer allí en su estadio temprano y migrar después como pequeñas poslarvas o juveniles desde el mar hacia aguas dulces (Blanco-Libreros et al., 2015). La única especie periférica identificada fue M. cephalus, la cual exhibe una marcada preferencia eurihalina (Ibáñez et al., 2012; Siccha-Ramírez et al., 2023). La mayor parte de su ciclo de vida transcurre en aguas de salinidad variable, adentrándose brevemente en aguas marinas durante su migración desde la costa hacia el mar para llevar a cabo el desove (Ibáñez et al., 2012).
En este estudio se identificaron tres especies endémicas del Perú: A. stalsbergi, cuya localidad tipo es el río Pisco, región de Ica (Musilová et al., 2009); T. punctulatus, con localidad tipo en el río Rímac, región de Lima (Fowler, 1945), y B. semotilus, que procede de los Andes peruanos a 4.000 m s.n.m., pero cuya localidad tipo se desconoce (Cope, 1874).
Si bien no existen compilaciones extensas sobre la fauna exótica presente en el Perú, se sabe que la mayor parte de los peces introducidos en Suramérica que han sido estudiados pertenece a las familias de los salmónidos y los cíclidos (Seebens et al., 2023). El Convenio sobre la Diversidad Biológica (CDB) define las especies exóticas invasoras como aquellas que se introducen y propagan fuera de su área natural y representan una amenaza para la diversidad biológica (MINAM, 2022). Aunque las invasiones biológicas son naturales, su impacto se agrava en hábitats vulnerables, causando graves daños a la biodiversidad local, como es el caso en Perú. Actualmente, no se conoce con certeza el número de especies no nativas establecidas en el país. Muchas de ellas ingresaron intencionalmente y se emplean como forraje, alimentación u ornamentación (MINAM, 2022). La naturalización de especies acuáticas exóticas en Perú se ha llevado a cabo por diversos motivos, entre ellos, el acuarismo (P. velifera), el control biológico de insectos (P. reticulata), y la acuicultura (O. niloticus, C. rendalli) (Cossíos, 2010). Por otra parte, en la década de los setenta, C. carpio fue importada desde Israel y Panamá con el propósito de llevar a cabo investigaciones en la Universidad Nacional Agraria La Molina (Ortega et al., 2007). Aquí registramos la presencia de siete especies exóticas, de las cuales dos (P. reticulata y Oreochromis sp.) se encontraron en todos los hábitats evaluados. Ortega et al. (2007) señalan que en los cuerpos de agua lénticos la adaptación de especies invasoras puede haberse visto favorecida por la presencia de fondos arenosos y aguas tranquilas, la ausencia de depredadores y la baja diversidad de especies nativas. Incluso en ciertos contextos han reducido, posiblemente de manera significativa, la distribución y abundancia de las especies nativas, principalmente mediante el desplazamiento y la depredación (Ortega et al., 2007).
Hoy las zonas costeras continentales son foco de invasiones de especies exóticas que afectan la riqueza y la abundancia de las especies nativas, aumentan el riesgo de extinción, alteran la composición genética, modifican el comportamiento y perturban las redes tróficas (Pysek et al., 2020). Además, muchas de estas especies alteran el funcionamiento de los ecosistemas y sus servicios. Estos impactos sobre la biodiversidad y los ecosistemas se han intensificado y se espera que aumenten en el futuro (Pysek et al., 2020). Ortega et al. (2007) señalaron que, entre todos los peces introducidos, las tilapias, con tres especies registradas en la cuenca baja del río Pisco, deberían ser motivo de especial preocupación. Además, O. niloticus y P. reticulata, mencionadas en el presente estudio, están catalogadas entre las 10 especies de peces exóticos invasores más ampliamente distribuidas en el mundo (Seebens et al., 2023). Ortega et al. (2007) señalaron que, como consecuencia del impacto causado por la ictiofauna invasora en la región costera de la cuenca río Grande (Ica), los peces nativos fueron completamente desplazados por O. niloticus y P. reticulata, especialmente cerca de la desembocadura del río durante la estación seca. Actualmente el MINAM (2022) ha presentado el "Plan de Acción Nacional sobre las Especies Exóticas Invasoras en el Perú 2022-2026" para el territorio nacional. El plan busca fortalecer el control de estas especies mediante la recopilación de información técnica que facilite su manejo, controle su dispersión y mitigue los impactos negativos en la diversidad biológica. Asimismo, se busca informar a la población peruana sobre los riesgos e impactos en la biodiversidad, la salud y la economía asociados a estas especies. En este sentido, Pysek et al. (2020) señalan que, aunque en muchos países hay estrategias para reducir futuras invasiones, su implementación suele ser ineficaz. A diferencia de otras dimensiones del cambio global, las invasiones biológicas pueden gestionarse y mitigarse con previsión y recursos adecuados. Sin embargo, a menudo reciben una atención insuficiente.
La reciente actualización de la IUCN (2023) destaca la difícil situación de los peces de agua dulce a nivel global, señalando que al menos el 17 % de las especies amenazadas de peces de agua dulce se ven afectadas por el cambio climático. La contaminación afecta al 57 % de los peces de agua dulce en peligro de extinción, mientras que el 33 % se ve perjudicado por especies invasoras y enfermedades. En el contexto peruano, la información sobre las categorías de amenazas es insuficiente y aún no se dispone de una Lista Roja oficial para peces continentales. Algunos peces endémicos como T. punctulatus y A. stalsbergi también se ven afectados por el cambio climático y la introducción de especies exóticas, lo que ha generado competencia por el hábitat y los recursos alimentarios (Ortega et al, 2012).
El impacto humano en lagos, ríos, humedales y aguas subterráneas está reduciendo de manera significativa la biodiversidad (Albert et al., 2021), así como el efecto de los desechos municipales y otros contaminantes industriales vertidos directamente en el río Pisco (MINAM, 2019). Además, el humedal Pisco Playa enfrenta impactos derivados de la urbanización creciente, las actividades agrícolas, la ganadería, la acumulación de residuos sólidos y la contaminación con aguas residuales (Martínez & Mendoza, 2022). La laguna Morón, catalogada como en proceso de extinción por el Inrena (2003), ha sufrido el impacto derivado de la visita anual de turistas. Asimismo, se observan áreas afectadas por la presencia de residuos sólidos y heces de ganado (ANA, 2018). Se tiene conocimiento de que la poza Lagunillas se ha secado, posiblemente debido a varios de los factores mencionadas.
Por último, nuestro estudio presenta una lista actualizada de las especies de peces del bajo río Pisco, con lo que se amplía el conocimiento sobre la ictiofauna de las cuencas del océano Pacífico en la costa peruana. Esta lista es una herramienta útil para el manejo y conservación de la biota acuática de esta región. A pesar de la poca diversidad registrada, se destaca la presencia de ictiofauna endémica y nativa que se ve amenazada por el creciente aumento de especies exóticas y la preocupante degradación de los ambientes acuáticos. Sería recomendable monitorear la ictiofauna en las cuencas media y alta, todavía no estudiadas, y evaluar periódicamente aspectos ecológicos de las especies mencionadas en este estudio.

