











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
La enfermedad de Alzheimer familiar (FAD) es una enfermedad asociada a factores de riesgo genéticos, especialmente debido a mutaciones en los genes que se traducen a proteína precursora de amiloide (PPA), presenilina-1 (PSEN-1) y presenilina-2 (PSEN-2). Esta enfermedad puede manifestarse en menores de 65 años y las mutaciones asociadas a variantes familiares, pueden producir deterioro cognitivo de manera precoz (Alonso vilatela, López-López, and Yesca-Gómez 2012; Rovelet-Lecrux et al. 2006; Shao, Peng, and Wang 2017). En el entendimiento de los mecanismos bioquímicos de procesamiento y producción del péptido amiloide, la enzima y-secretasa realiza un procesamiento proteasa con su sitio activo la proteína presenilina-1 (PS1), ubicado en los dos ácidos aspárticos con localización en las TM6 y TM7 (Zhou, R; Yang, Guanghui; Guo Xuefei; Zhou, Qiang; Lei, jianlin and Shi 2019; Ballard et al. 2011; Martinelli 2022), esta enzima realiza los cortes de manera transmembranal y el punto inicial de procesamiento es el corte épsilon en los aminoácidos LV (49 aminoácido) o TL (48 aminoácido) para el sustrato PPA CTF99, dependiendo de ese corte inicial se hace una ruta de procesamiento que consiste en los cortes sucesivos de 49-46-43 hasta el corte y en el aminoácido 40 y los cortes sucesivos cuando la ruta es 48-45 hasta el corte y en el aminoácido 42 o 38 dependiendo de la difusión del sustrato y la interacción con su bolsillo activo (Szaruga et al. 2017; Hitzenberger and Zacharias 2019). La enzima y-secretasa es una aspartil proteasa muy importante en el metabolismo celular y se encuentra relacionada con el procesamiento de aproximadamente 93 sustratos, los cuales ayudan en funciones de señalización celular, adhesión, expresión, diferenciación celular, neuro-desarrollo, movilización o anclaje en membrana celular y regulación de factores de transcripción, todo esto dependiendo del tipo de sustrato (Kaether, Haass, and Steiner 2006; Medoro et al. 2018; Golriz Khatami et al. 2022).
Enfocados en el complejo enzimático de y-secretasa, en especial en su sitio activo con la sub unidad presenilina-1 (PS1), la cual tiene 7 isoformas, pero en el cerebro, la isoforma que más se expresa, es aquella que tiene 467 aminoácidos y que exhibe 9 pases transmembrana (Apweiler et al. 2014; Soto-Ospina, Araque Marín, Bedoya, Sepulveda-Falla, et al. 2021). Para ser activado el complejo enzimático, aún no se conoce muy bien el mecanismo, pero en la literatura se propone el mecanismo de endo-proteólisis, que supone un autocorte del loop que une las TM6 y TM7, lo que produce un N-terminal y C-terminal, los cuales sirven para el reconocimiento de los sustratos (Saura et al. 2000; Levitan et al. 2001; Khan, Ali, and Mirza 2020). Esta proteína participa en la activación de varios sustratos por reacciones de rompimiento de péptidos, pero también se caracteriza porque tiene algunos sitios de fosforilación que pueden ilustrarse, por ejemplo, en la región C-terminal, en el aminoácido Ser346 ubicado en el motivo de reconocimiento de caspasa donde la PKC quinasa se fosforila para el procesamiento proteolítico de PS1 y la activación de la apoptosis (Fluhrer et al. 2004; Zhang, Chen, and Lee 2020). De la misma manera, los aminoácidos Ser353, Ser357 y Ser397 son fosforilados por la proteína quinasa GSK-3B, que está involucrada en la interacción con los sustratos de B-catenina y N-cadherina, que tienen funciones en la expresión de la superficie celular y la señalización celular de PS1. La proteína quinasa GSK-3B también tiene un papel en la alteración del procesamiento del péptido B-amiloide y también está relacionada con la proporción 40/42 de fragmentos B-amiloide (Soto-Ospina, Araque Marín, Bedoya, Sepulveda-Falla, et al. 2021; Soto-Ospina, Araque Marín, Bedoya, and Villegas Lanau 2021; Okochi et al. 2013; Fernandez et al. 2014; L. Sun et al. 2016).
Esta importancia funcional puede verse afectada por mutaciones que pueden aparecer en su información genética, lo que produce cambios en el desempeño de su funcionalidad. En humanos esta proteína tiene reportadas aproximadamente 327 mutaciones (Alzforum 2022), las cuales incluyen variantes sin sentido, deleciones e inserciones, que producen diversas patologías, por lo que, en los estudios evolutivos sobre la proteína, es crucial observar los efectos y propuestas de las mutaciones en la especiación. La proteína presenilin-1 es paráloga de presenilin-2 (Khan, Ali, and Mirza 2020), son dos proteínas que se traducen de dos genes localizados en dos cromosomas diferentes como lo son el cromosoma 1 y 14 (Clark et al. 1995; Vidal et al. 1996), pero por motivos de este trabajo, se centrará en entender a nivel molecular presenilina-1, la cual cuenta con una plantilla estructural, obtenida por crio-microscopía electrónica, que proporciona la base para la propuesta de modelado de las proteínas del grupo de homínidos y permite el acercamiento, desde el punto de vista funcional, a nivel computacional, cuyo foco principal es la comprensión estructural a partir de la visualización de genomas, transcriptomas y proteomas (Haas et al. 2017; Y. V. Sun and Hu 2016; Bruggeman and Westerhoff 2007).
Materiales y métodos
Selección de homínidos y Macacos
La búsqueda de la secuencia primaria de aminoácidos, se hizo para una muestra de cinco homínidos, los cuales fueron: Orangután de Sumatra (Pongo abelii), Chimpancé (Pan troglodytes), gorila del este (Gorilla-gorilla), Bonobo (Pan paniscus) y Humano (Homo sapiens); los macacos se escogieron porque han sido ampliamente usados en bioensayos en neurodegeneración y se utilizaron las especies Macaco cangrejero (Macaca fascicularis) y Macaco Rhesus (Macaca mulatta), ya que se tenía la información estructural depurada para esas especies. Las bases de datos utilizadas fueron UniProt para recolectar la secuencia de los aminoácidos, la búsqueda de las estructuras proteicas disponibles se hizo con Protein Model Portal (PMP) y en el Protein Data Bank (PDB) (Berman et al. 2012; UniProt 2010; Consortium 2012) , la plantilla que se usó para el modelamiento molecular tiene ID: 6IYC, obtenida por crio-microscopía electrónica con una resolución de 2.60A (Zhou, R; Yang, Guanghui; Guo Xuefei; Zhou, Qiang; Lei, jianlin and Shi 2019).
Alineamiento de secuencias primarias
Al obtener las secuencias primarias en formato FASTA para los homínidos a partir de las bases de datos, se hizo un alineamiento de estas con el software Jalview (Waterhouse et al. 2009), seleccionando el histograma normalizado sin consenso, asimismo se hizo el cálculo de índice de similaridad ID a partir de las herramientas T-Coffee 2.0 (Notredame, Higgins, and Heringa 2000) y ClustalX (Larkin et al. 2007) para estudiar la conservación potencial de los fragmentos estructurales entre la muestra de homínidos seleccionados.
Modelos de predicción estructural para PS1
La secuencia primaria de aminoácidos para cada proteína de la especie, fue sometida en el software predictor de estructura de proteína y péptidos Phyre2 (Kelley et al. 2015), el cual produce construcción de modelos por homología con estructuras reportadas y comparación de regiones de aminoácidos por su secuencia con el reconocimiento de plegamientos estructurales en folding y ensamblaje por threading (hilamiento), se van uniendo los fragmentos encontrados por similitud, teniendo en cuenta el esqueleto atómico en los ensambles (Kelley et al. 2015).
Refinamiento de los modelos estructurales
El refinamiento de los modelos obtenidos por Phyre2 se hizo con el software Modeller (Webb and Sali 2016), con el cual se hizo el refinamiento de las regiones flexibles, con el fin de evitar solapamientos de los loops dentro de la misma estructura y localización en las hélices alfa de las estructuras localizadas en la membrana, esto se hace a partir del algoritmo ZDOPE (Energía de la proteína discreta optimizada) (Shen and Sali 2006), el cual presenta un estimado estadístico a partir de la distancia interatómica, entregando cinco modelos con calificación normalizada, de los cuales se seleccionó el mejor modelo calificado (Krieger et al. 2009; Webb and Sali 2016).
Visualizadores estructurales
Los visualizadores tridimensionales se usaron en el alineamiento y en la representación de los modelos ensamblados, los alineamientos se hacen con el software Chimera U.C.S.F. versión 1.11 (Pettersen et al. 2004), para ello se usa la comparación estructural con Matchmaker, para los parámetros de matriz de sustitución de bloques BLOSUM62 (Henikoff and Henikoff 1992) y el algoritmo de programación dinámica que produce un alineamiento global Needleman-Wunsch (Needleman and Wunsch 1970), para proteínas, ácidos nucleicos y canales de proteína transmembranal. La identificación y localización topológica se hace con el software Deepview/Swiss-PdbViewer versión 3.7 (N. Guex and M. C. Peitsch 1997).
Modificaciones postraduccionales
Las secuencias primarias de aminoácidos para cinco especies de homínidos y dos especies de macacos, se analizaron a partir del software suite ExPASy, utilizando las herramientas para calcular la glicosilación en los aminoácidos que tienen aminas en su cadena lateral N-glicosilación y O-glicosilación para los aminoácidos que tienen grupos funcionales oxigenados y disponibles para interactuar (Artimo et al. 2012). De igual forma, se evaluaron las secuencias primarias de la proteína PS1 de tipo silvestre en el servidor NetPhosK 3.1, con el fin de predecir sitios susceptibles de fosforilación debido a serina, treonina o tirosinas por diversas quinasas, por ejemplo, quinasas PKA, PKC, PKE, RSK, EGFR o MAPK38 (Blom et al. 2004; Blom, Gammeltoft, and Brunak 1999).
Resultados
Para los homínidos seleccionados en el análisis de secuencia primaria, se hizo la búsqueda en la base de datos UniProt para la proteína presenilina-1, la cual se considera en el análisis con la misma isoforma de un tamaño de 467 aminoácidos entre la muestra evaluada.
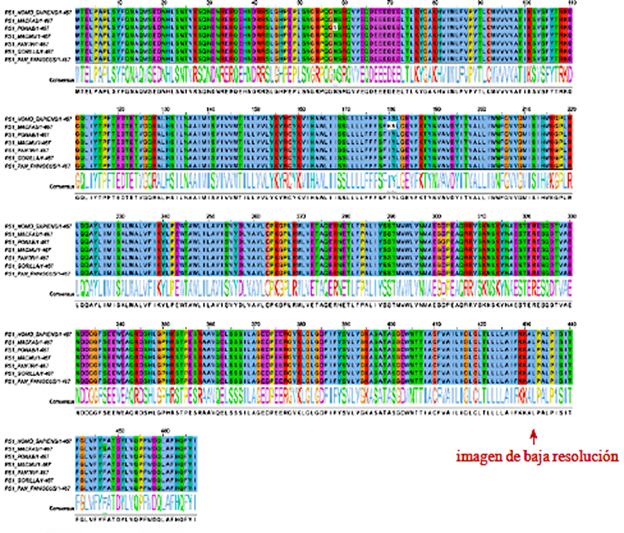
El alineamiento de las secuencias primarias de aminoácidos, se hizo con el software Jalview y el porcentaje de identidad del alineamiento se hizo con el software T-Coffee 2.0, presentando el alineamiento entre las especies Homo sapiens-Macaca fascicularis (Macfas), Pongo abelii (Ponab)- Homo sapiens, Ponab-Macfas, Macaca mulatta (Macmu)- Homo sapiens, Macmu-Macfa, Macmu-Ponab, Pan troglodytes (Pantro)-Macfas, Ponab-Pantro, Homo sapiens-Gorilla gorilla (Gor), Homo sapiens-Pan paniscus (Panis) y Pantro-Macmu, el cual tiene un valor de similitud de 99,69 % como promedio de los alineamientos evaluados. En la Figura 1, se presenta el alineamiento de la estructura para su secuencia primaria de aminoácidos, luego de realizar su respectiva traducción y obtener los aminoácidos constituyentes en formato FASTA, T-Coffee 2.0 consideró todos los aminoácidos, mostrando algunos cambios debido a mutaciones, las cuales, al no participar en el consenso, el software las trata sin relleno de color, evidenciando las regiones de mutación:
Fuente: Elaborada por los autores
Figura 1 Alineamiento de secuencia primaria de la proteína PS1 para los homínidos Homo sapiens, Macaca fascicularis, Pongo abelii, Macaca mulatta, Pan troglodytes, Gorilla gorilla y Pan paniscus.
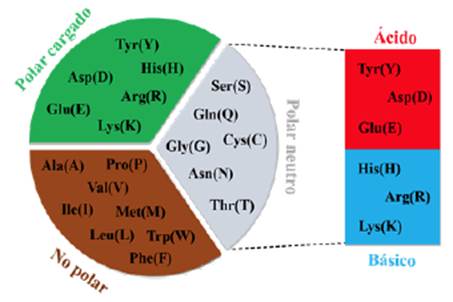
En el alineamiento con la herramienta T-Coffee 2.0 del software Jalview y código de colores ClustalX que considera la naturaleza química polar, no polar, aromática, neutra, ácido o básica para cada aminoácido perteneciente a la secuencia de composición proteica, en donde se pudo obtener un alineamiento de las secuencias de PS1467. El consenso muestra el logomat de la secuencia, en la que se observan regiones muy conservadas a lo largo de la secuencia proteica, pero para Pongo abelii hay una mutación en la región flexible del loop que corresponde al N-terminal y también una mutación en la región que da al C-terminal de PS1467. Otra de las secuencias que presenta mutaciones en la región que comprende la transmembrana 3 (TM3) en la especie Macaca fascicularis, se presentan dos cambios adyacentes, lo que cambia el ambiente químico en el que estaba la estructura proteica. Para analizar los cambios que se generaron entre especies, bajo una conceptualización bioquímica, se realiza un agrupamiento de los aminoácidos, teniendo en cuenta su naturaleza, como se muestra en la Figura 2:
Fuente: Elaborada por los autores
Figura 2 Diagrama de clasificación de la naturaleza química de los aminoácidos
Según las variantes en los aminoácidos encontrados para la especie Pongo Abelii que en la posición 58 tiene Ser(S) en vez de Asn(N), indica que hay un cambio en el aminoácido con un grupo funcional diferente en su cadena lateral. Asimismo, para la especie Macaca fascicularis se obtienen un par de mutaciones adyacentes en la posición 180Ile(I)-180Tyr(Y), por los aminoácidos 181Lys(K)-181Asn(N), los cuales están localizados en la transmembrana 3 extracelular, esta mutación presenta un cambio de no polar-polar cargado a un aumento de polaridad de polar cargado-polar neutro.
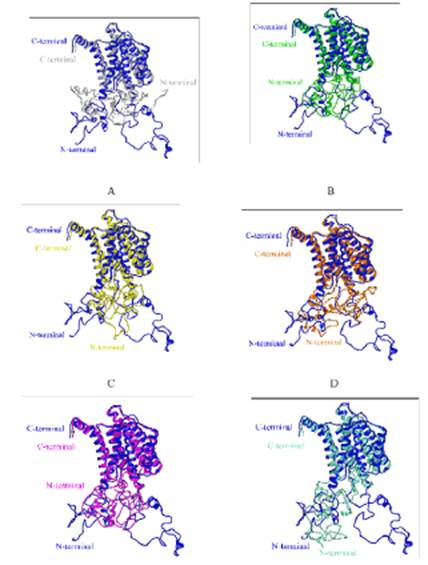
Considerando la secuencia primaria de las especies de homínidos para la proteína PS1467, se hizo la modelación molecular de la estructura de la proteína para cada una de las secuencias de las muestras de homínidos, con el fin de determinar si existen diferencias importantes a nivel estructural, para ello se hace el uso del predictor de estructura Phyre2, el cual permite predecir la estructura de aquellas proteínas que experimentalmente se les desconoce, dadas las difíciles condiciones experimentales de aislamiento para obtener la proteína o los problemas de flexibilidad en las regiones loops que producen zonas de baja electrodensidad, incluso la característica química de los aminoácidos que no permite la nucleación del cristal. Se hace el modelamiento molecular y luego a cada modelo hipotético se le hace un alineamiento estructural (ver Figura 3), en el que se tiene como referente en cada alineamiento 3D la proteína PS1467 Homo sapiens (color azul). Estos alineamientos permiten visualizar si en el sistema se producen cambios a nivel tridimensional, lo que afectaría la función de la proteína en su sitio activo y se tendría el efecto directo de las variantes según la especiación de los homínidos:
Fuente: Elaborada por los autores
Figura 3 Alineamiento estructural de PS1467Humana (azul) ID: 6IYC con PS1467de: A) Macfas (plateado), B) Macmu (verde), C) Pantro (amarillo), D) Ponab (naranja), E) Gor(magenta)y F) Panis (aguamarina). Visualización en cintas con el software UCSF Chimera 1.11.
En la visualización de los alineamientos tridimensionales por pares para la proteína PS1467 en las especies de homínidos, se puede apreciar que en el dominio transmembrana de hélices alfa hay una alta coincidencia estructural, es decir, se presenta superposición estructural entre las mismas (ver Figura 3. A, B, C, D, E y F). No obstante, en la región intracelular de la proteína es donde la coincidencia estructural difiere, ya que dicha región está constituida mayoritariamente por loops, los cuales pueden tomar diferentes conformaciones. Lo mencionado con anterioridad, se refuerza con los datos listados en la Tabla 1, donde se presenta la desviación cuadrática media (RMSD) estimada para cada uno de los alineamientos tridimensionales, obtenidas mediante el algoritmo de Needleman-Wunsch.
Tabla 1 Desviación cuadrática media (RMSD) estimada de cada uno de los alineamientos tridimensionales por pares PS1 homínidos
| Alineamiento 3D de PS1 | RMSD estimada [A] | Alineamiento 3D de PS1 | RMSD estimada [A] |
|---|---|---|---|
| Homo Sapiens-Macfas | 4,41 | Macfas-Panis | 2.56 |
| Homo Sapiens-Macmu | 3,39 | Macmu-Pantro | 1.55 |
| Homo Sapiens-Pantro | 4,21 | Macmu-Ponab | 1,14 |
| Homo Sapiens-Ponab | 2,56 | Macmu-Gor | 0,03 |
| Homo Sapiens-Gor | 3,26 | Macmu-Panis | 1,66 |
| Homo Sapiens-Panis | 2,96 | Pantro-Ponab | 1,39 |
| Macfas-Macmu | 1,43 | Pantro-Gor | 1,64 |
| Macfas-Pantro | 3,16 | Pantro-Panis | 1,81 |
| Macfas-Ponab | 4,11 | Gor-Panis | 1,13 |
| Macfas-Gor | 1,17 |
Fuente: Elaborada por los autores
En la Tabla 1, se observa que en el alineamiento estructural de PS1467 Homosapiens con los otros homínidos hay una menor desviación estándar con la especie Ponab (RMSD=2,56 Â) y la especie Panis (RMSD=2,96 A), mientras que hay mayor desviación estándar (RMSD=4,41 Â) con la especie Macfas que presentan varias mutaciones en PS1467. Asimismo, se tiene que los alineamientos 3D de PS1467 entre las especies de homínidos examinados, excluyendo la PS 1 467 Homo sapiens, presentan desviaciones estándar en el rango de 0,03-4,11 Â, siendo el alineamiento 3D de Macmu-Gor la de menor desviación estándar (RMSD=0,03 Â) y la Macfas-Ponab (especies con mutaciones) la de mayor desviación estándar (RMSD=4,11 Â). Sin embargo, las anteriores no superan una desviación estándar de 5Â, por tanto, se puede decir que no hay diferencias significativas en la distancia promedio entre los átomos alineados tridimensionalmente para PS 1 467 entre las especies evaluadas.
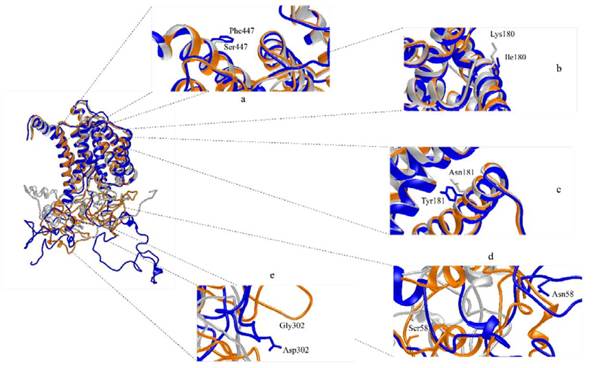
Cada una de las variantes encontradas en los alineamientos de Jalview se modelan para visualizar el efecto en Pongo abelii y Macaca fascicularis, las dos estructuras fueron comparadas para la región que comprende sus variaciones respecto a Homo sapiens, con el fin de entender el efecto a partir de la tridimensionalidad estructural como se muestra en la Figura 4.
Fuente: Elaborada por los autores
Figura 4 Alineamiento estructural de PS1 467 Humana (azul) ID: 6IYC con PS1 467 de Pongo abelii (Naranja)y Macaca fascicularis (Plateado), donde se observa variaciones en residuos: a) Phe447a Ser447, b) Ile180 a Lys180, c) Tyr181 a Asn181, d) Asn58 a Ser58y e) Asp302 a Gly302. Visualización en cintas con el software UCSF Chimera 1.11.
Discusión
En el alineamiento de las secuencias primarias de homínidos para la proteína PS1 presentó variantes para Pongo Abelii, ya que se cambió una amida por un grupo hidroxilo, pero la naturaleza del aminoácido no se altera, debido a que el aminoácido sigue siendo polar neutro, esto quiere decir, que esa posición puede mantener las interacciones en la formación de aductos o dipolo-dipolo de Keesom; por su localización en el loop flexible de la región N-terminal de PS1467, tiene un papel importante en la interacción con las otras subunidades en el ensamble de la enzima y-secretasa como APH1 y PEN-2 que son dos proteínas andamiaje del sitio activo. Además, la variación al residuo de Ser(S) puede posibilitar que en procesos postraduccionales o incluso en membrana, sea probable la O-Glicosilación y la fosforilación por quinasas y, con ello, contribuir con la funcionalidad de la proteína (ver Tabla 2). Por otro lado, en la posición 302, sí hay un cambio de polaridad: de polar cargado Asp(D) a polar neutro Gly(G), pero sigue localizado en las regiones loop y no afecta en su distribución tridimensional.
Para Macaca Fascicularis, ocurren dos cambios adyacentes, ya que se producen ligeros cambios en el ambiente químico de la TM3, como se puede evidenciar para el cálculo de pases transmembranales (ver Tabla 2). Sin embargo, la estructura tridimensional tiende a mantenerse, dado que la variante se localiza en la porción extracelular de TMD3. También se observa cambios en la posición 447, del aminoácido Phe(F) al aminoácido Ser(S), adicionando un nuevo sitio de fosforilación a la proteína, ya que es posible ver el efecto directo en las estructuras desde la discusión resumida en la Tabla 2:
Tabla 2 Sitios de: fosforilación, O-Glicosilación, N-Glicosilación y pasos transmembrana de PS1 Para las especies Homo sapiens, Macfas y Ponab
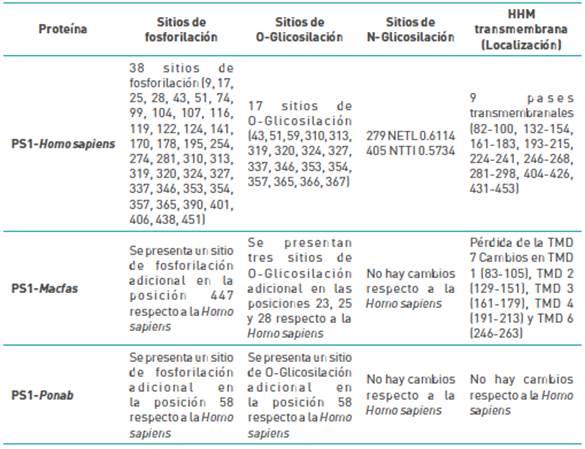
Fuente: Elaborada por los autores.
Las especies con variaciones que se revelaron en la secuencia primaria estudiada con Jalview, fueron alineadas de manera global con Matchmaker del software Chimera U.C.S.F, con el fin de comparar si hay un efecto en la funcionalidad, por lo que se relacionan Homo sapiens con Pongo abelii y con Macus fascicularis, mostrando que la proteína del Macaco a pesar de tener varios cambios en las zonas un poco más rígidas como las a-hélice en la membrana, se evidencia que no hay cambios estructurales marcados e incluso la hélice no se pierde por cambio de la variante en la modelación, aunque tenga un ligero desplazamiento en el alineamiento y la naturaleza del aminoácido en primera instancia, no parece afectar la estabilidad de la estructura secundaria. Las variantes del homínido Pongo abelii, las describimos con anterioridad a partir de su naturaleza química y a nivel estructural. Esta variante se encuentra localizada en las regiones flexibles y no parece inducir ningún efecto respecto al sitio activo de la proteína que queda entre la transmembrana 6 y 7, para los aminoácidos ácido aspártico 257 y 385, dejando como perspectiva un análisis considerando las otras subunidades del complejo enzimático, para evaluar su efecto a nivel de estructura cuaternaria.
Además, se puede observar en los alineamientos globales que las estructuras de presenilina-1, en términos generales, presentan una muy buena conservación de los fragmentos que no son tan flexibles, es decir, el sitio activo y las alfa hélices que presentan los nueve pases transmembranales que coinciden de igual forma con la isoforma 1 que es la más representativa en cerebro, con una constitución de 467 aminoácidos. Los cambios en las regiones flexibles, cuya funcionalidad se asocia en el reconocimiento de sustratos y en las interacciones con las subunidades laterales, las cuales reconocen y movilizan los sustratos cuando se tiene el complejo y-secretasa ensamblado, es fundamental para la estructura proteica y su funcionalidad, pero los cambios no difieren de la estructura de Homo sapiens de referencia, como se puede mostrar cuantitativa y cualitativamente.
Por otro lado, es importante señalar, que al revisar las mutaciones patogénicas para Homo sapiens reportadas en la base de datos de Alzforum, no se encuentran variantes en las posiciones 58 o 302, que corresponden a las variaciones en la secuencia para el Pongo abelii (ver Figura 4. d y e), aunque se encuentra reportada una variante aledaña al sitio 302, que corresponde al cambio Pro(P) por Leu(L) en el aminoácido 303 y que ha sido asociada a demencia frontotemporal (Koriath et al. 2020). Para el caso de las variaciones en Macaca fascicularis, no se han reportado mutaciones patogénicas para Homo sapiens en la posición 447 (ver Figura 4. a), tampoco se han reportado variaciones en la posición 181(ver Figura 4. c), sin embargo, si se encuentra reportada una variante en el codón 180, que lleva al cambio de Ile (I) por Asn (N), que se considera posiblemente responsable de causar FAD de inicio precoz (Hélène-Marie et al. 2017). El cambio de Ile (I) por Asn (N) (en el humano) o por Lys (K) (en el macaco) (ver Figura 4. b), aunque son diferentes aminoácidos, en ambos casos, se generan un cambio en la polaridad; sin embargo, se debe considerar que la PS1 es solo una porción del complejo y-secretasa y que puede haber variaciones en las subunidades que hagan que los complejos se ensamblen con cambios entre estas especies, haciendo que estas variaciones no tengan efecto sobre la función enzimática.
Es notoria la homología entre las secuencias de PSEN1 en los homínidos (100 % en 4 de los 5 estudiados), indicando una función importante conservada en todas estas especies. Probablemente está homología en las secuencias, tiene relación con los diferentes reportes de cambios involutivos en los cerebros de los simios, similares a los que se presentan en el Homo sapiens durante el envejecimiento. Aunque lo común son reportes de depósitos del péptido B-amiloide, también se describen con menor frecuencia ovillos neurofibrilares y pérdida de memoria o cambios comportamentales durante el envejecimiento, lo cual claramente está en relación con los cambios neuropatológicos para la enfermedad de Alzheimer (Gearing et al. 1997; Edler et al. 2017; King 2018).
Esta proteína y su función proteasa es importante en los mecanismos patológicos, en donde el péptido amiloide se le asocia funcionalidad a nivel de señalización celular, con el fragmento dominio intracelular AICD. A nivel tridimensional no se observan diferencias, lo que quiere decir que muy probablemente, bajo una visión computacional, los mecanismos se pueden desarrollar de igual manera para los homínidos evaluados y la muerte neuronal se daría con el envejecimiento en estas especies. Este estudio como perspectiva, podría complementarse con un análisis experimental, en donde se tengan todas las enzimas y-secretasa y se evalúen cinéticas enzimáticas de procesamiento del sustrato PPA, para Homo sapiens, Pongo abelii y Macaca Fascicularis.
Conclusión
Entender la enfermedad de Alzheimer y el papel del mecanismo patogénico en la actualidad, conlleva a realizar análisis desde una perspectiva molecular, del complejo multiproteico y-secretasa y en últimas de la subunidad comprometida con el sitio activo y la funcionalidad proteasa. PS1 es una proteína muy conservada entre las especies y para los homínidos, también presentó solo unas ligeras diferencias en los lazos flexibles, pues cabe recordar que estas regiones son dinámicas y se le asocia la función de reconocer moléculas y realizar la auto proteólisis como el corte de la proteína para poder activarse. Con los resultados actuales puede concluirse que para la proteína PS1, no hay cambios considerables en su estructura para la muestra de homínidos, como se demuestra con el cálculo de RMSD de los alineamientos por pares, en donde se mostraron valores por debajo de 5 A y las mutaciones que se tenían en algunas especies como Macaca Fascicularis y Pongo abelii, están localizadas en regiones muy flexibles y alejadas de los sitios activos, en donde podrían tener un efecto directo en la funcionalidad de PS1. Como aspecto relevante, queda realizar el modelado de las enzimas completas para una muestra mucho más grande de especies, que incluso no quede restringida sólo a organismos eucariotas, de igual forma determinar la función y ver qué organismos tienen la presencia de las subunidades.

